T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
EKĠNEZYA TÜRLERĠNĠN (Echinacea pallida (Nutt) Nutt., Echinacea purpurea (L.) Moench)
DOKU KÜLTÜRÜ TEKNĠKLERĠ ĠLE ÇOĞALTILMASI ve IN VITRO KOġULLARDA
SEKONDER METABOLĠT ĠÇERĠĞĠNĠN ARAġTIRILMASI
Münüre TANUR ERKOYUNCU
DOKTORA TEZĠ
Tarla Bitkileri Anabilim Dalını
Aralık-2019 KONYA Her Hakkı Saklıdır
TEZ KABUL VE ONAYI
Münüre TANUR ERKOYUNCU tarafından hazırlanan ―EKĠNEZYA
TÜRLERĠNĠN (Echinacea pallida (Nutt) Nutt., Echinacea purpurea (L.) Moench) DOKU KÜLTÜRÜ TEKNĠKLERĠ ĠLE ÇOĞALTILMASI ve IN VITRO KOġULLARDA SEKONDER METABOLĠT ĠÇERĠĞĠNĠN ARAġTIRILMASI‖
adlı tez çalıĢması 23/12/2019 tarihinde aĢağıdaki jüri tarafından oy birliği ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü Tarla Bitkileri Anabilim Dalı‘nda DOKTORA TEZĠ olarak kabul edilmiĢtir.
Jüri Üyeleri Ġmza
BaĢkan
Prof. Dr. Yüksel KAN ………..
DanıĢman
Doç. Dr. Mustafa YORGANCILAR ………..
Üye
Prof. Dr. Semiha ERĠġEN ………..
Üye
Prof. Dr. Özden ÖZTÜRK ………..
Üye
Doç. Dr. Derya ARSLAN DANACIOĞLU ………..
Yukarıdaki sonucu onaylarım.
Prof. Dr. Mustafa YILMAZ FBE Müdürü
Bu tez çalıĢması Bilimsel AraĢtırma Koordinatörlüğü tarafından 18101018 nolu proje ile desteklenmiĢtir.
TEZ BĠLDĠRĠMĠ
Bu tezdeki bütün bilgilerin etik davranıĢ ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalıĢmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that. as required by these rules and conduct. I have fully cited and referenced all material and results that are not original to this work.
Ġmza
Münüre TANUR ERKOYUNCU 23.12.2019
ÖZET
DOKTORA TEZĠ
EKĠNEZYA TÜRLERĠNĠN (Echinacea pallida (Nutt) Nutt. Echinacea purpurea (L.) Moench) DOKU KÜLTÜRÜ TEKNĠKLERĠ ĠLE ÇOĞALTILMASI ve IN
VITRO KOġULLARDA SEKONDER METABOLĠT ĠÇERĠĞĠNĠN
ARAġTIRILMASI
Münüre TANUR ERKOYUNCU
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Tarla Bitkileri Anabilim Dalı
DanıĢman: Doç. Dr. Mustafa YORGANCILAR
2019, 144 Sayfa Jüri
Doç. Dr. Mustafa YORGANCILAR Prof. Dr. Yüksel KAN
Prof. Dr. Semiha ERĠġEN Prof. Dr. Özden ÖZTÜRK
Doç. Dr. Derya ARSLAN DANACIOĞLU
Bu çalıĢmada, Echinacea pallida ve Echinacea purpurea L. türlerinde kallus ve sürgün kültürlerinin oluĢturulması ile hem etkili bir rejenerasyon ve mikroçoğaltım sisteminin kurulması hem de
in vitro kültür koĢullarının optimizasyonu ile sekonder metabolit içeriğinin artırılması amaçlanmıĢtır.
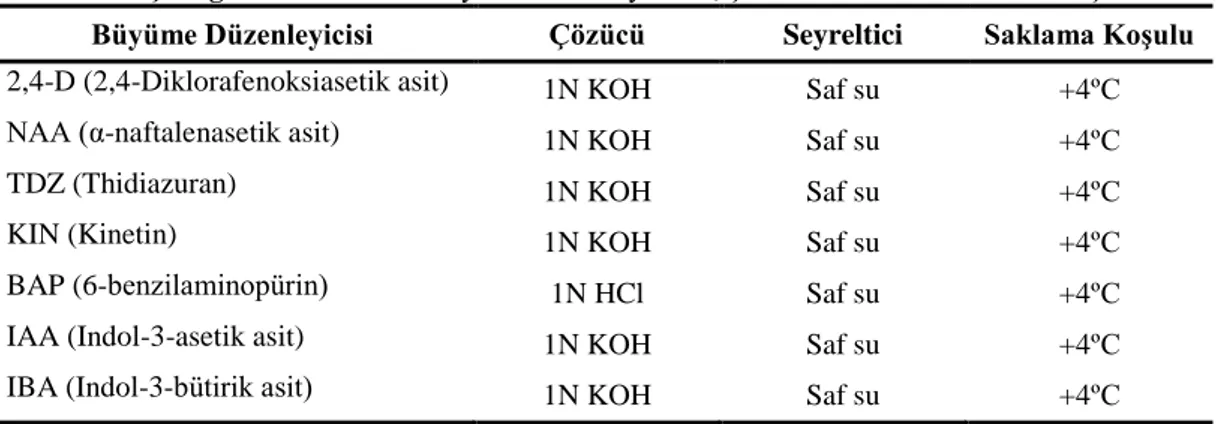
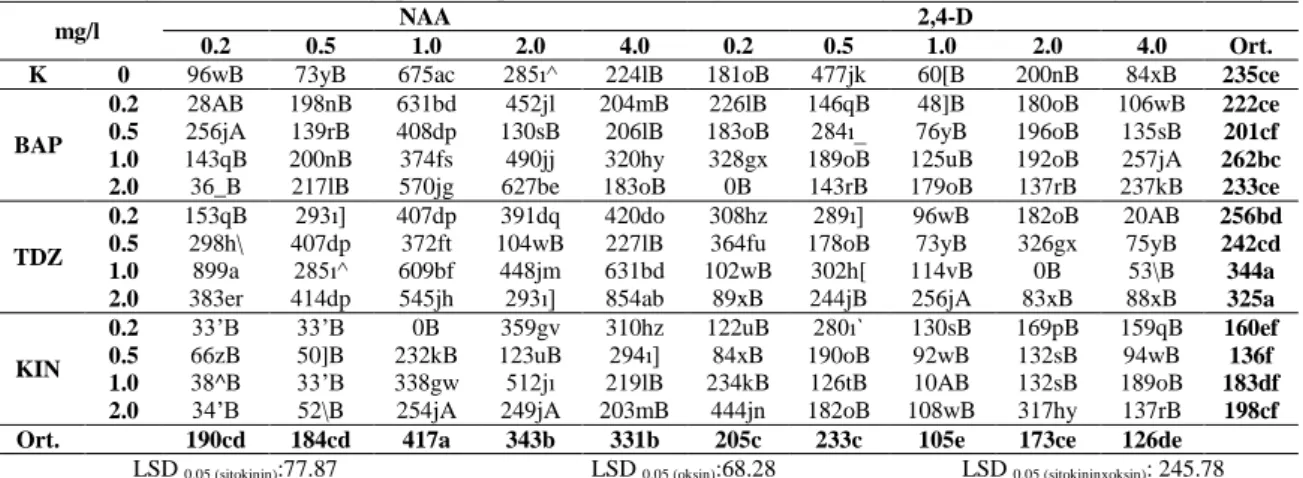
AraĢtırmada her iki türün yaprak, yaprak sapı, kotiledon ve kök eksplantlarından farklı tip ve konsantrasyonlarda büyüme düzenleyicileri içeren besin ortamlarında baĢarılı bir Ģekilde kallus uyarımı sağlanmıĢtır. Her eksplant tipi için en iyi kallus geliĢimi; E. purpurea türünde 1.0 mg/l 2,4-D+2.0 mg/l BAP (yaprak), 1.0 mg/l NAA+0.5 mg/l TDZ (yaprak sapı), 2.0 mg/l NAA+1.0 mg/l TDZ (kotiledon), 0.5 mg/l NAA+0.5 mg/l BAP (kök) içeren ortamlarda, E. pallida türünde ise 0.2 mg/l NAA+1.0 mg/l TDZ (yaprak), 4.0 mg/l NAA+0.5 mg/l TDZ (yaprak sapı), 4.0 mg/l NAA+0.2 mg/l TDZ (kotiledon), 0.5 mg/l NAA+0.2 mg/l TDZ (kök) içeren ortamlarda sağlanmıĢtır. Her iki bitki türünün, dört farklı eksplant tipi, belirlenen büyüme düzenleyicilerinde 4, 6, 8 ve 10 hafta boyunca kültüre alınmıĢ ve elde edilen kallus dokularında kaftarik asit, klorojenik asit, kafeik asit ve kikorik asit miktarları belirlenmiĢtir. En yüksek kaftarik asit (4.11 mg/g) ve kikorik asit (57.89 mg/g) miktarları E. purpurea L. türünün yaprak sapı eksplantlarından, klorojenik asit (8.83 mg/g) ise kök eksplantlarından 10 haftalık kültür süresi sonunda elde edilen kalluslarda tespit edilmiĢtir. Büyüme düzenleyicisi ve kültür süresi optimizasyonu yapılan kallus kültürlerinde, farklı azot oranlarının, farklı karbon kaynakları ve miktarlarının, farklı polietilen glikol (PEG) miktarlarının sekonder metabolit üretimi üzerine etkileri incelenmiĢtir. Bu uygulamalar arasında, en yüksek kaftarik asit (9.38 mg/g), klorojenik asit (0.71 mg/g), kafeik asit (0.29 mg/g) ve kikorik asit (34.77 mg/g) miktarları, 0:35 mM amonyum/nitrat oranı içeren besin ortamında 10 haftalık
kültür süresi sonunda E. purpurea L. türünün kök eksplantından elde edilen kallus dokularında tespit edilmiĢtir.
Mikroçoğaltım amacıyla, her iki türün nodal segmentlerinden farklı tip ve konsantrasyonlarda büyüme düzenleyicisi içeren ortamlarda çoklu sürgün uyarımı baĢarılı bir Ģekilde gerçekleĢtirilmiĢtir. E.
purpurea L. türünde en yüksek sürgün oluĢumu farklı konsantrasyonlarda BAP ve KIN içeren ortamlarda,
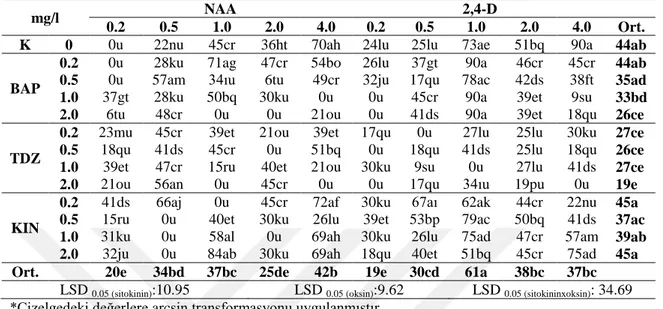
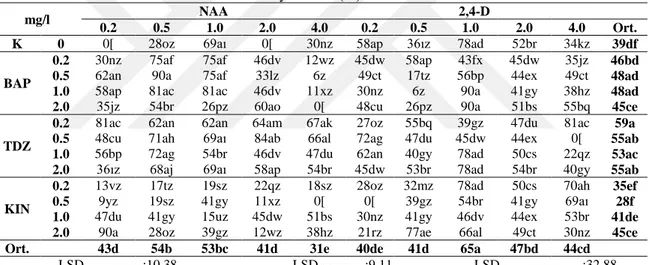
eksplant baĢına sürgün sayısı değeri ise (3.47 adet) 0.50 mg/l BAP + 0.10 mg/l NAA içeren ortamda elde edilmiĢtir. E. pallida türünde ise en yüksek sürgün oluĢumu BAP ve BAP‘ın NAA ile kombinasyonlarını içeren ortamlarda, eksplant baĢına sürgün sayısı ise (2.45 adet) 0.50 mg/l BAP + 0.50 mg/l NAA içeren ortamda elde edilmiĢtir. Her iki türde elde edilen sürgünler, kök uyarımı için farklı konsantrasyonlarda (0.5, 1.0 veya 1.5 mg/l) ve tiplerde oksin (NAA, IBA, IAA) içeren ve içermeyen besin ortamlarında kültüre alınmıĢtır. En yüksek köklenme oranları, E. purpurea L.‘da %29, E. pallida‘da ise %6 olmak üzere büyüme düzenleyicisi içermeyen ortamlarda elde edilmiĢtir.
Elde edilen sürgünlerde farklı tip ve konsantrasyonlardaki büyüme düzenleyicilerinin sekonder metabolit miktarlarına etkileri belirlenmiĢtir. E. purpurea L. türünde, en yüksek kaftarik asit (7.41 mg/g) 4.0 mg/l BAP, klorojenik asit (5.04 mg/g) 2.0 mg/l KIN, kikorik asit miktarları ise (46.27 ve 45.22 mg/g) sırasıyla 4.0 mg/l KIN+0.5 mg/l NAA ve 2.0 mg/l BAP + 0.5 mg/l NAA içeren ortamlardan elde edilen sürgünlerde tespit edilmiĢtir. E. pallida türünde ise en yüksek kaftarik asit (6.43 mg/g) 4.0 mg/l BAP+0.1 mg/l NAA, klorojenik asit (3.47 mg/g) 0.50 mg/l BAP+0.10 mg/l NAA, kikorik asit miktarı ise (24.83 mg/g) 1.0 mg/l BAP içeren ortamlarda elde edilen sürgünlerde tespit edilmiĢtir.
Bu tez çalıĢmasının sonucunda, önemli iki ekinezya türünde kallus ve sürgün kültürleri aracılığıyla etkili ve tekrarlanabilen rejenerasyon ve mikroçoğaltım protokolleri belirlenmiĢ olup, aynı zamanda in vitro koĢullarda kafeik asit türevlerinin içeriğinin artırılmasına yönelik kültür koĢullarının optimizasyonu ve farklı uygulamaların etkileri belirlenmiĢtir.
Anahtar Kelimeler:Ekinezya, Kafeik asit türevleri, Kallus kültürü, In vitro çoğaltım, Sekonder metabolit, Sürgün kültürü
ABSTRACT
Ph.D THESIS
PROPAGATION OF ECHINACEA SPECIES (Echinacea pallida (Nutt) Nutt.,
Echinacea purpurea (L.) Moench) USING TISSUE CULTURE TECHNIQUES
AND INVESTIGATION OF SECONDARY METABOLITE CONTENT UNDER
IN VITRO CONDITIONS
Münüre TANUR ERKOYUNCU
The Graduate School of Natural and Applied Science of Selçuk University Departman of Field Crops
Advisor: Assoc. Prof. Dr. Mustafa YORGANCILAR
2019, 144 Pages
Jury
Assoc. Prof. Dr. Mustafa YORGANCILAR Prof. Dr. Yüksel KAN
Prof. Dr. Semiha ERĠġEN Prof. Dr. Özden ÖZTÜRK
Assoc. Prof. Dr. Derya ARSLAN DANACIOĞLU
In this study, it was aimed to increase the secondary metabolite content by the formation of callus and shoot cultures in Echinacea pallida and Echinacea purpurea L. species, both by establishing an effective regeneration and micropropagation system and by optimization of in vitro culture conditions.
Callus induction was achieved in media containing growth regulators of different types and concentrations from leaf, petiole, cotyledon and root explants of both species. Best callus development for each explant type; In E. purpurea species, 1.0 mg/l 2,4-D+2.0 mg/l BAP (leaf), 1.0 mg/l NAA+0.5 mg/l TDZ (petiole), 2.0 mg/l NAA+1.0 mg/l TDZ (cotyledon), 0.5 mg/l NAA + 0.5 mg/l BAP (root) was provided in the media, while E. pallida species 0.2 mg/l NAA+1.0 mg/l TDZ (leaf), 4.0 mg/l NAA+0.5 mg/l TDZ (petiole), 4.0 mg/l NAA+0.2 mg/l TDZ (cotyledon), 0.5 mg/l NAA+0.2 mg/l TDZ (root) was provided in the media. Both plant species were cultured for 4, 6, 8 and 10 weeks in growth regulators with four different explant types and the amounts of caftaric acid, chlorogenic acid, caffeic acid and chicoric acid were determined in the callus tissues obtained. The highest amounts of caftaric acid (4.11 mg/g) and chicoric acid (57.89 mg/g) were obtained from petiole explants of E. purpurea L., the amount of chlorogenic acid (8.83 mg/g) was determined in callus obtained from root explants after 10 weeks of culture. The effects of different nitrogen ratios, different carbon sources and amounts, different amounts of polyethylene glycol (PEG) on secondary metabolite production were investigated in callus cultures with growth regulator and culture time optimized. Among these applications, the highest amounts of caftaric acid (9.38 mg/g), chlorogenic acid (0.71 mg/g), caffeic acid (0.29 mg/g) and chicoric acid (34.77
mg/g), 0:35 mM ammonium/nitrate rate of nutrient medium containing 10 weeks of culture at the end of
E. purpurea L. species obtained from the root explants were determined in callus tissues.
For the purpose of micropropagation, multiple shoot induction was successfully performed in media containing growth regulators of different types and concentrations from nodal segments of both species. The highest shoot formation in E. purpurea L. species was obtained in media containing different concentrations of BAP and KIN and the number of shoots per explant (3.47) was obtained in medium containing 0.50 mg/l BAP+ 0.10 mg/l NAA. In E. pallida species, the highest shoot formation was obtained in the medium containing BAP and BAP with NAA, and the number of shoots per explant (2.45) was obtained in the medium containing 0.50 mg/l BAP + 0.50 m /l NAA. The shoots obtained from both species were cultured in media with and without auxin (NAA, IBA, IAA) in different concentrations (0.5, 1.0 or 1.5 mg/l) and types for root induction. The highest rooting rates were obtained in 29% of E.
purpurea L. and 6% of E. pallida without growth regulators.
The effects of growth regulators of different types and concentrations on secondary metabolite amounts were determined in the shoots obtained. In E. purpurea L., the highest caftaric acid (7.41 mg/g) 4.0 mg/l BAP, chlorogenic acid (5.04 mg/g) 2.0 mg/l KIN, and the chicoric acid amounts (46.27 and 45.22 mg / g) were respectively, 4.0 mg/l KIN + 0.5 mg/l NAA and 2.0 mg/l BAP + 0.5 mg/l NAA. In E.
pallida species, the highest amount of caftaric acid (6.43 mg/g) 4.0 mg/l BAP + 0.1 mg/l NAA,
chlorogenic acid (3.47 mg/g) 0.50 mg/l BAP+0.10 mg/l NAA and chicoric acid (24.83 mg/g) 1.0 mg/l BAP.
As a result of this thesis, effective and reproducible regeneration and micropropagation protocols were determined by using callus and shoot cultures of two important echinacea species, and the effects of different conditions and optimization of culture conditions for increasing the content of caffeic acid derivatives in vitro were determined.
Keywords: Caffeic acid derivatives, Callus culture, Echinecea, In vitro propagation, Secondary metabolites, Shoot culture
ÖNSÖZ
ÇalıĢma konusunun belirlenmesi baĢta olmak üzere, araĢtırmanın planlanmasından yürütülmesine ve sonuçların değerlendirilmesine kadar her aĢamasında bilgi ve tecrübelerinden faydalandığım, her türlü desteği sağlayan, önerileri ile bana farklı bakıĢ açıları kazandıran danıĢman hocam, Sayın Doç. Dr. Mustafa YORGANCILAR‘a teĢekkürlerimi sunarım.
Tez çalıĢmamın önemli bir kısmını oluĢturan HPLC analizlerini yapabilmeme olanak sağlayan, S.Ü. Ziraat Fakültesi Tıbbi Bitkiler Laboratuvarlarının tüm imkânlarını bana sunan, destek ve yardımlarını esirgemeyen tez izleme jüri üyesi hocam, Sayın Prof. Dr. Yüksel KAN‘a, Dr. S. AyĢe ÇELĠK‘e ve ArĢ. Gör. Ġrem AYRAN‘a, doku kültürü çalıĢmalarımda katkı ve önerilerde bulunan, yardım ve teĢvikleri ile bana yol gösteren tez izleme jüri üyesi hocam, Sayın Prof. Dr. Semiha ERĠġEN‘e en içten teĢekkürlerimi sunarım.
Tez çalıĢmalarım boyunca, bilgi ve tecrübelerinden her daim faydalandığım çok değerli hocam, Sayın Dr. Öğretim Üyesi Emine ATALAY‘a, istatistiki analizlerde bilgi ve görüĢlerini paylaĢarak katkıda bulunan, desteği ile her zaman yanımda olan çok değerli çalıĢma arkadaĢım, Sayın Dr. Kevser YAZAR‘a, özveri gerektiren uzun ve yoğun laboratuvar çalıĢmaları sırasında yardımlarını esirgemeyen Zir. Müh. Havva ÇAĞLAR‘a, Tarla Bitkileri Bölümü lisans öğrencilerinden Fatmanur UÇAR ve Ġrem BÖLÜKBAġI‘na sonsuz teĢekkür ederim.
Bugünlere gelmemde büyük emeği olan, hayatımın her aĢamasında maddi ve manevi desteğini esirgemeyen aileme; bu çalıĢma için beni sürekli teĢvik eden, her daim yanımda olan, desteğini esirgemeyen sevgili eĢime ve bu süreçte beni sabırla bekleyen oğlum Ġsmail Tuna‘ya, göstermiĢ oldukları fedakârlıklardan dolayı sonsuz teĢekkürlerimi sunarım.
Münüre TANUR ERKOYUNCU KONYA-2019
ĠÇĠNDEKĠLER
ÖZET ... i
ABSTRACT ... iii
ÖNSÖZ ... v
ĠÇĠNDEKĠLER ... vi
SĠMGELER VE KISALTMALAR ... viii
1. GĠRĠġ ... 1
2. KAYNAK ARAġTIRMASI ... 5
2.1. Ekinezyanın Taksonomisi, Morfolojisi ve YetiĢtiriciliği ... 5
2.2. Ekinezyanın Tarihsel ve Popüler Kullanımı ... 6
2.3. Ekinezyanın Kimyasal BileĢimi ... 8
2.4. Ekinezya Türlerinde Doku Kültürü ÇalıĢmaları ... 11
2.5. Ekinezya Türlerinde Doku Kültürü ile Sekonder Metabolit Üretiminin AraĢtırılması ile Ġlgili ÇalıĢmalar ... 15
3. MATERYAL VE YÖNTEM ... 24
3.1. Materyal ... 24
3.2. Yöntem ... 24
3.2.1. In vitro çalıĢmalar için yapılan ön hazırlıklar ... 24
3.2.2. Tohumların yüzey sterilizasyonu ve çimlendirilmesi ... 26
3.2.3. Kallus kültürleri ... 27
3.2.4. Sürgün kültürleri ... 28
3.2.5. Sürgünlerin köklendirilmesi ve dıĢ ortama aktarılması ... 29
3.2.6. Kallus kültürlerinde sekonder metabolit üretimini artırmaya yönelik uygulamalar ... 30
3.2.7. Kallus ve sürgünlerde sekonder metabolitlerin analiz edilmesi ... 31
3.2.8. Verilerin elde edilmesi ... 35
3.2.9. Verilerin istatistiki değerlendirilmesi ... 36
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 37
4.1. Kallus Kültürleri ... 37
4.1.1. Echinecea purpurea L. türünde kallus kültürlerinin araĢtırılması ... 37
4.1.2. Echinecea pallida türünde kallus kültürlerinin araĢtırılması ... 53
4.2. Sürgün Kültürleri ... 68
4.2.1. Echinecea purpurea L. türünde sürgün kültürlerinin araĢtırılması ... 68
4.2.2. Echinecea pallida türünde sürgün kültürlerinin araĢtırılması ... 72
4.3. Sürgünlerin Köklendirilmesi ve DıĢ Ortama Aktarma ... 81
4.3.1. Echinecea purpurea L. türünde köklendirme çalıĢmaları ... 81
4.3.2. Echinecea pallida türünde köklendirme çalıĢmaları ... 85
4.4.1. Echinecea purpurea L. kallus ve sürgün kültürlerinde sekonder
metabolitlerin analizi ... 86
4.4.2. Echinecea pallida kallus ve sürgün kültürlerinde sekonder metabolitlerin analizi ... 97
4.5. Kallus kültürlerinde sekonder metabolit üretimini artırmaya yönelik uygulamalar ... 103
4.5.1. Farklı azot miktarlarının etkisi ... 103
4.5.2. Farklı karbon kaynaklarının ve konsantrasyonlarının etkisi ... 104
4.5.3. Farklı polietilen glikol (PEG) konsantrasyonlarının etkisi ... 107
4.6. Eksplant kaynağı olarak kullanılan fidelerin (E. pallida ve E. purpurea L.) sekonder metabolit içeriği ... 108
5. SONUÇLAR VE ÖNERĠLER ... 110 5.1. Sonuçlar ... 110 5.2. Öneriler ... 113 KAYNAKLAR ... 115 EKLER ... 128 ÖZGEÇMĠġ ... 143
SĠMGELER VE KISALTMALAR Simgeler °C: Santigrat Derece mg/l: Miligram/Litre g/l: Gram/ Litre ml: Mililitre a/h: Ağırlık/Hacim
Lux: Birim Alana DüĢen IĢık Akısı Ölçü Birimi M: Molar
mM: Milimolar µM: Mikromolar
Kısaltmalar
UV: Ultraviyole ıĢın HCI: Hidroklorik asit KOH: Potasyum hidroksit EtOH: Etil alkol
atm: Atmosfer basıncı dk: Dakika
2,4-D: 2,4-Diklorafenoksiasetik asit NAA: α-naftalenasetik asit
TDZ: Thidiazuran KIN: Kinetin
BAP: 6-benzilaminopürin IAA: Indol-3-asetik asit IBA: Indol-3-bütirik asit 2iP: 2-izo-penteniladenin WPM: Woody plant medium
1. GĠRĠġ
Ekinezya türleri, anavatanı Kuzey Amerika olan, Asteraceae familyasına ait, fitoterapide kullanılan çok yıllık, otsu bitkilerdir (Barrett, 2003; Toselli ve ark., 2009). Dünyada Black Sampson, Purple/Red Coneflower gibi farklı isimler verilen ekinezya türleri, ülkemizde ise yaygın olarak ekinezya, pembe/mor koni çiçeği veya kirpiotu gibi isimler ile bilinmektedir (Wichtl, 2004; Çelik ve Kan, 2019).
Ekinezya cinsi, 9 tür altında toplanmıĢ olup (McGregor, 1968), bunlardan özellikle üçü Echinacea purpurea (L.) Moench, Echinacea angustifolia DC. var. angustifolia ve Echinacea pallida (Nutt.) Nutt. tıbbi bitki olarak bilinen ve ekonomik öneme sahip olan türleridir (Barnes ve ark., 2005).
Ekinezyanın geleneksel tıpta kullanımı çok uzun bir geçmiĢe sahip olup, ilk olarak Amerika yerlileri tarafından boğaz ağrısı, soğuk algınlığı, diĢ eti ve bademcik iltihaplanması, yanık ve yaraların iyileĢmesi, kuduz, böcek ve yılan zehirlenmelerinin tedavisinde kullanılmıĢtır (Bauer ve Wagner, 1990). Günümüzde ise ekinezyanın tıbbi kullanımı, çoğunlukla bağıĢıklık sistemini uyarıcı/güçlendirici etkisine dayanmakta olup, bakteriyel ve viral hastalıkları tedavi etmek ve önlemek için kullanılmaktadır (Toselli ve ark., 2009). Ayrıca, iltihap önleyici ve antitoksin olarak tanımlanan ekinezyanın, hafif septisemi (kan zehirlenmesi), fronküloz (deride sürekli tekrarlayan ağrılı ve kalıcı nodüller), çıban, karbonkül (deri Ģarbonu) gibi hastalıkların tedavisinde kullanılmasının (Kligler, 2003; Barnes ve ark., 2005) yanı sıra, tümör (Currier ve Miller, 2002; Lee ve ark., 2010) ve kanser hücrelerinin (Chicca ve ark., 2007; Tsai ve ark., 2012) büyümesini durdurduğu da belirlenmiĢtir (Bruni ve ark., 2018).
Ekinezya türlerinin tıbbi olarak kullanımına imkân veren, biyoaktif içeriğini oluĢturan bileĢikler temelde iki ana gruba ayrılmaktadır. Bunlar; alkamitler gibi lipofilik bileĢikler (Bauer ve Remiger, 1989), çoğunlukla kafeik asit türevlerinden oluĢan polisakkaritler ve glukoproteinleri de kapsayan hidrofilik bileĢiklerdir (Pellati ve ark., 2004; Thude ve Classen, 2005). Bu bileĢikler, türlerde antioksidan, antiviral, antibakteriyel, antienflamatuvar (iltihap önleyici) (Merali ve ark., 2003; Sharma ve ark., 2010), immunostimulatör (bağıĢıklık sistemini uyarıcı) ve yara iyileĢtirici (Bauer ve Remiger, 1989; Kligler, 2003; Zhai ve ark., 2009) etkileri oluĢturan fitokimyasallardır.
Kafeik asit türevleri (kikorik asit, ekinakosit, klorojenik asit, kaftarik asit, kafeik asit, sinarin), E. purpurea L., E. pallida ve E. angustifolia türlerinin karakteristik fenolik bileĢikleridir. Kafeik asit türevlerinin kanıtlanmıĢ bağıĢıklık sistemini uyarıcı,
yara iyileĢtirici ve antioksidan aktivitelerinin yanı sıra (Hu ve Kitts, 2000; Ardjomand-Woelkart ve Bauer, 2016), antiviral (Hudson ve Vimalanathan, 2011), antihiyalüronidaz (Facino ve ark., 1993) ve yüksek serbest radikal temizleme özelliği (Pellati ve ark., 2004) de dâhil olmak üzere birçok biyoaktif fonksiyona sahip oldukları belirlenmiĢtir. Kafeik asit türevlerinden özellikle kikorik asit yukarıda bahsedilen biyoaktif fonksiyonlara ilaveten, anti-diyabet (Azay-Milhau ve ark., 2013), anti-obezite (Xiao ve ark., 2013) ve anti-HIV-AIDS virüsü (Chhipa ve ark., 2014) etkileriyle de tıbbi kullanımı açısından oldukça önemli bir yere sahiptir (Ramezannezhad ve ark., 2019).
Dünyada son yıllarda sentetik maddelerin olumsuz etkilerinin ortaya çıkması ve tüketici bilincinin oluĢmasıyla birlikte bitkisel kökenli doğal ürünlere olan talep artmakta olup, doğal gıda, ilaç ve kozmetik pazarı ciddi oranda büyüme göstermektedir (Temel ve ark., 2018). BaĢta Avrupa ülkeleri olmak üzere birçok ülke özellikle ilaç ve kozmetik sanayinin hammaddelerini oluĢturan bitkileri üreterek ve iĢleyerek ekonomilerine büyük katkılar sağlamaktadır. Bu bitkiler arasında, artan tıbbi değeri ve çok yönlü kullanım alanları sayesinde ekinezyanın popülerliği giderek artmıĢ ve son yıllarda Avrupa ve Amerika‘nın en çok satan tıbbi bitkisi haline gelmiĢtir (Smith ve ark., 2015).
Ekinezya türleri, tıbbi etkinliği tek bir kimyasal maddeye atfedilemeyen, bu nedenle de tüm metabolitlerin tam spektrumunu elde etmek için bitkisel üretimi gereken tıbbi bitkilerdir (Pellati ve ark., 2004; Jones ve ark., 2009a). Ayrıca ekinezya türlerinin temel biyoaktif içeriğini oluĢturan bileĢiklerin sentezinin zor, maliyetli ve zaman alıcı olması ve artan tüketime karĢılık bitkisel üretiminin yetersiz kalması nedeniyle doğada yetiĢen bitkilerden de bu bileĢiklerin elde edildiği belirlenmiĢtir (Abbasi ve ark., 2012). Pek çok tıbbi bitkide olduğu gibi, ekinezya türlerinde de doğadan toplanan bitkilerde, istenen standart ve kalite sağlanamadığı gibi bitki neslinin tükenmesi tehlikesi de ortaya çıkmaktadır. Ekinezyanın bitkisel üretimi ise, bitki materyalinin mikroorganizmalarla bulaĢması, çevreden kaynaklanan kirlilik, aktif bileĢenlerin değiĢkenliği ve biyokimyasal analizler için saf, standartlaĢtırılmıĢ bitki materyalinin olmaması gibi çeĢitli sebeplerden dolayı yetersiz düzeyde kalmaktadır (Raman ve ark., 2004).
Tüm bu problemlerin önüne geçmek için sekonder metabolitlerin üretiminde biyoteknolojik yöntemler, özellikle bitki hücre ve doku kültürleri alternatif bir yöntem olarak görülmektedir. Sekonder metabolitleri, bitki hücre ve doku kültürleri yoluyla kültür koĢulları optimizasyonu sağlayarak üretmek; bitkinin kültürü esnasında karĢılaĢılan çevresel etkenleri (iklim, coğrafi zorluklar, mevsimsel kısıtlamalar) ortadan
kaldırılmakta, daha az arazi kullanımı sağlanmakta, bitkinin doğadan toplanarak neslinin yok olması engellenmekte, bitkilerde düĢük miktarlarda bulunan ekonomik açıdan değerli metabolitlerin yeterli miktarda üretilebilmesi ve üretimde homojenite, standart kalite ve verimliliğin oluĢturulması gibi avantajlar sağlamaktadır (Rao ve Ravishankar, 2002). Tüm bu avantajlar sayesinde sekonder metabolitlerin üretimi klasik yöntemlere kıyasla daha hızlı, basit, güvenilir ve öngörülebilir bir Ģekilde yapılabilmektedir.
Ekinezya türlerinden bitki hücre ve doku kültürleri yoluyla sekonder metabolitlerin üretilmesi için öncelikle etkili ve tekrar edilebilen bir rejenerasyon sisteminin oluĢturulması, sonrasında ise sekonder metabolit içeriğinin artırılmasına yönelik optimum in vitro kültür koĢullarının ve farklı elisitör uygulamalarının araĢtırılması gerekmektedir. Bugüne kadar yapılan araĢtırmalar, genellikle türlerde kallus ve/veya sürgün kültürleriyle etkili rejenerasyon sisteminin kurulması (Choffe ve ark., 2000b; Coker ve Camper, 2000; Koroch ve ark., 2002; Zebarjadi ve ark., 2011), mikroçoğaltım sistemi ile kitlesel üretimin sağlanması (Harbage, 2001; Jones ve ark., 2007; Li ve ark., 2013) ve bu üretimden standart kalite ve verimlilikte sekonder metabolitlerin elde edilmesi üzerine olmuĢtur (Taha ve ark., 2009; Butiuc-Keul ve ark., 2012). Özellikle bu türlerde sekonder metabolit üretimini artırmaya yönelik in vitro kültür koĢullarının optimizasyonu ile farklı elisitör uygulamalarının etkilerinin araĢtırıldığı çalıĢmalar yeterli düzeyde bulunmamaktadır.
Bu tez çalıĢmasında, E. pallida ve E. purpurea L. türlerinde, kallus ve sürgün
kültürlerinin oluĢturulması ile etkili bir rejenerasyonun sisteminin belirlenmesi, in vitro Ģartlar altında sekonder metabolit içeriğinin artırılmasına yönelik kültür koĢullarının optimizasyonu ve farklı elisitör uygulamalarının araĢtırılması amaçlanmıĢtır. Her iki türe ait dört faklı eksplant tipinde (yaprak, yaprak sapı, kotiledon, kök), farklı tip ve konsantrasyonlarda büyüme düzenleyicileri içeren besin ortamlarında kallus uyarımı sağlanarak her eksplant tipi için en iyi kallus geliĢimini sağlayan büyüme düzenleyicisi kombinasyonu tespit edilmiĢtir. Bu eksplantlardan farklı kültür sürelerinde (4-6-8-10 hafta) elde edilen kalluslarda sekonder metabolit miktarları belirlenmiĢtir. Büyüme düzenleyicisi ve kültür süresi bakımından optimizasyon yapılan kallus dokularında, besin ortamında yer alan beslenme faktörlerinin (azot miktarı, karbon kaynağı ve miktarı) değiĢiminin ve abiyotik elisitör olarak PEG‘in (kuraklık) sekonder metabolit üretimi üzerine etkileri incelenmiĢtir. Her iki türün nodal segmentleri, çoklu sürgün çoğaltımını teĢvik etmek amacıyla farklı tip ve konsantrasyonlardaki büyüme
düzenleyicilerini içeren besin ortamlarında kültüre alınmıĢ, en etkili büyüme düzenleyicisi/düzenleyicileri belirlenmiĢ ve her iki türde, elde edilen sürgünlerde farklı tip ve konsantrasyonlardaki büyüme düzenleyicilerinin sekonder metabolit miktarlarına etkileri tespit edilmiĢtir. Aynı zamanda, indirekt sürgün uyarımını teĢvik etmek ve tam bir rejenerasyonu sağlayabilmek için elde edilen kalluslar, farklı büyüme düzenleyicilerini içeren ortamlarda kültüre alınmıĢ ve organogenesis gerçekleĢtirilmiĢtir. Son olarak, in vitro koĢullarda elde edilen sürgünler farklı konsantrasyon ve tipte oksin içeren ortamlarda köklendirilerek mikroçoğaltım süreci tamamlanmıĢtır.
2. KAYNAK ARAġTIRMASI
2.1. Ekinezyanın Taksonomisi, Morfolojisi ve YetiĢtiriciliği
Asteraceae/Compositae familyasına ait, gen merkezi Kuzey Amerika olan ekinezya türleri ilk kez McGregor tarafından, 1968 yılında taksonomik olarak sınıflandırılmıĢ ve ekinezya cinsi 9 tür altında toplanmıĢtır. McGregor (1968)‘a göre bu türler; E. angustifolia DC, E. atrorubens Nutt., E. laevigata (Boyton & Beadle) Blake,
E. pallida (Nutt.) Nutt., E. paradoxa (Norton) Britton, E. purpurea (L.) Moench, E. simulata McGregor, E. sanguinea Nutt., E. tennesseensis (Beadle) Small Ģeklinde
isimlendirilmiĢlerdir. Daha sonra Binns ve ark. (2002a) tarafından moleküler, morfometrik ve fitokimyasal varyasyona dayanarak revize edilen Echinacea cinsi taksonomisi, 4 tür altında toplanmıĢ olup, Mc Gregor‘un belirttiği dokuz türün beĢi (angustifolia, sanguinea, tennesseensis, simulata, paradoxa) E. pallida türünün varyeteleri olarak tanımlanmıĢtır (ÇalıĢkan ve OdabaĢ, 2011). Yakın zamanda Zhang ve ark. (2017) tarafından kloroplast genom verilerine göre de bir sınıflandırma yapılmıĢ olmakla birlikte, McGregor'un (1968) geleneksel taksonomisi hala yaygın Ģekilde kullanılmaktadır.
Ekinezya türleri, aynı familyada yer aldığı ayçiçeği, aspir, yer elması gibi bitkilerle benzer morfolojik özellikler göstermektedir. Çok yıllık otsu bitkiler olan ekinezya türlerinde, gövde dik geliĢir ve tüylüdür; nadiren dallanma gösterir, yapraklar alt kısımlarda bir yaprak sapıyla, uç kısımlarda ise direkt olarak gövdeye bağlanır, gövdenin en uç kısmında çiçek tablaları bulunur. Ekinezya türleri, dikenimsi çiçekcikleri ve koni biçimindeki çiçek tablasıyla karakterize edilmekte olup; çiçek tablası; fertil ve steril çiçekler ile çanak ve taç yapraklardan oluĢmaktadır. Yabancı döllenen bitkiler olup, genel olarak kazık köklüdürler (Li, 1998; Mistríková ve Vaverková, 2007).
E. pallida ve E. purpurea L. türleri arasında morfolojik olarak bazı farklılıklar
bulunmaktadır. E. pallida türünde bitki boyu 40 ile 90 cm arasında değiĢirken, E.
pupurea‘da bitki boyu 60 ile 180 cm arasındadır. E. pallida‘da kök yapısı ana kazık kök
iken E. purpurea‘da dallanmıĢ kazık kök sistemi görülmektedir. E. pallida‘da yapraklar mızrak Ģeklinde olup, E. purpurea‘da türün en belirgin özelliği olarak ovalden geniĢ mızrağa kadar değiĢen, düzensiz tırtıklı Ģekillerde yapraklar mevcuttur. Çiçek rengi her
iki türde de koyu mordan beyaza kadar değiĢmektedir (McKeown, 1999; Demirezer ve ark., 2007).
Ekinezya serin iklim bitkisi olmasına rağmen, sıcak yaz aylarına ve kuraklığa karĢı yüksek tolerans göstermekte olup, genellikle fakir, kayalık, iyi drene olmuĢ alkali ve nötr pH‘ya yakın topraklarda yetiĢmektedir. Bitkinin yetiĢtiriciliği tohumların doğrudan tarlaya ekilmesiyle, fidelerin yetiĢtirilerek tarlaya ĢaĢırtılmasıyla ve kök çeliği ile yapılabilmekte olup, en yaygın yetiĢtirme yöntemi ise maliyetli olmasına karĢın diğer yetiĢtirme yöntemlerine kıyasla daha yüksek verim sağlamasından dolayı fide ile üretimdir (ÇalıĢkan ve OdabaĢ, 2011).
2.2. Ekinezyanın Tarihsel ve Popüler Kullanımı
Ekinezya bitkisinin geleneksel tıpta kullanımı çok eskilere dayanmakta olup, 1870‘lerde ilk kez tıp dünyasına tanıtan isim Dr. Meyer olmuĢtur (Hobbs, 1994). Bitkinin kullanılıĢını Amerika yerlilerinden öğrenen Dr. Meyer, E. angustifolia köklerinden hazırladığı droglara kan temizleyici anlamına gelen "blood purifier" adını vermiĢtir. Ġlacını romatizma, migren, ağrı, yılancık, yaralar, hazımsızlık, bitki zehirlenmeleri, zehirli yılan sokması, sifilis, kangren, malarya, difteri, hemoroit gibi birçok hastalığın tedavisinde kullanmıĢtır. Dr. Meyer'in ilacı 1887'de Dr. John King ve Ecz. John Uri Lloyd'un dikkatini çekmiĢ ve böylece ekinezya üzerindeki ilk bilimsel çalıĢma King ve Lloyd tarafından yapılmıĢtır. BaĢlangıçta yalnızca E. angustifolia kökleri kullanılırken daha sonra E. pallida kökleri de kullanılmıĢtır (Mat, 2004; ÇalıĢkan ve OdabaĢ, 2011).
1930‘larda antibiyotiklerin ortaya çıkması ve yüksek oranda tedavi özelliğine sahip olması nedeniyle ekinezyaya olan ilgi düĢmeye baĢlamıĢtır. 1960-1970‘lerde ise bilim adamları bağıĢıklık sisteminin kanser hücrelerinin denetlenmesinden sorumlu olduğunu tespit etmiĢler ve bağıĢıklık sistemini güçlendirici bitkiler üzerinde araĢtırmalarını artırmıĢlardır. Aynı zamanda antibiyotik kullanımından ortaya çıkan sorunlar da insanları bitkisel kaynakları kullanmaya yönlendirmiĢtir (Schar, 1999). Böylece, ekinezya tekrar gündeme gelmiĢ ve birçok araĢtırma yapılmıĢ, tıbbi önemi klinik çalıĢmalarla kanıtlanmıĢ ve halen araĢtırmalar yapılmakta olup, ticari ürünler çıkarılmıĢ ve ekonomik değer kazanmıĢtır.
Son yıllarda ise bitkisel kökenli doğal ürünlere talebin artmasıyla birlikte ekinezya ürünlerinin kullanımı da dikkat çekici bir Ģekilde artmıĢtır. Ekinezya
türlerinden elde edilen pek çok ürün, baĢta Amerika ve Avrupa ülkeleri olmak üzere, bitkisel drog olarak kullanılmakta olup, dünya bitkisel ilaç pazarında önemli bir paya sahiptir (Barrett, 2003). Ticari ekinezya preparatları birkaç farklı formda (tablet, kapsül veya Ģurup) olabildiği gibi tüm bitkiyi ya da bitkinin sadece belli parçalarını (kök, çiçek tablası, üst kısımlar) içerebilmektedir (Barnes ve ark., 2005). Doğrudan preslenmiĢ meyve sularından, tüm bitkinin etanolik özütlerine veya kurutulmuĢ köklere, yapraklara ve çiçeklere kadar çeĢitli ticari ürünler tüm dünyada eczanelerde, sağlık ve gıda marketlerinde bulunmaktadır (Barrett, 2003; Bruni ve ark., 2018) (ġekil 2.1).
Ekinezya, Amerika‘da 2013 yılında bir önceki yıla göre %95‘lik artan satıĢ oranıyla, yılın en çok satan sekizinci bitkisi olurken (Lindstrom ve ark., 2014); 2014 yılı itibariyle satıĢ oranı 2013‘e göre %79 oranında artmıĢ ve en çok satan üçüncü bitki olmuĢtur (Smith ve ark., 2015). Avrupa‘da ise ekinezyadan elde edilmiĢ 280‘den fazla ticari ürünle en çok satan tıbbi bitki haline gelmiĢtir (ÇalıĢkan ve OdabaĢ, 2011; Yu ve ark., 2013).
Doğadan toplanarak elde edilen ekinezya türlerinin artan tüketimle birlikte yetiĢtiriciliği yaygınlaĢmıĢ ve ekim alanları artmıĢtır. BaĢlıca Amerika ve Avrupa‘da kültürü yapılan ekinezyanın ekim alanları Kuzey Amerika ve Avrupa‘nın ötesinde Güney Amerika, Avustralya ve dünyanın diğer bölgelerine yayılmıĢtır (Yu ve Kaarlas, 2004). Avrupa'da ekinezyanın önde gelen üreticileri Almanya, Ġsviçre, Hollanda, Ġtalya ve Ġspanya'dır (Abbasi ve ark., 2007a). Ülkemizde ise doğal florada bulunmayan ekinezya türlerinin baĢta Konya olmak üzere Ġç Anadolu Bölgesi koĢullarına adaptasyonunun belirlenmesi amacıyla çeĢitli çalıĢmalar yürütülmüĢ ve ekonomik olarak yetiĢtirilebileceği belirlenmiĢtir (YeĢil ve Kan, 2015). Bununla birlikte bitkinin özellikle küresel ısınmanın en fazla görüldüğü Ġç Anadolu Bölgesi‘nde yetiĢtirilmesi Türkiye‘nin hemen her bölgesinde de yetiĢtirilebileceği potansiyelini ortaya çıkarmaktadır (Çelik, 2016).
ġekil 2.1. Ekinezya türlerine ait bazı ticari ürünler
2.3. Ekinezyanın Kimyasal BileĢimi
Ekinezya türlerinin kimyasal bileĢimi temelde fenolik bileĢikler (kafeik asit türevleri), alkamitler (alkilamitler), glukoproteinler ve polisakkaritler olmak üzere dört grup biyoaktif bileĢenden oluĢmaktadır (Bruni ve ark., 2018). Ayrıca ekinezya türlerinin flavonoit, alkaloit ve uçucu yağ bileĢenlerini içerdiği de çeĢitli çalıĢmalarda belirlenmiĢtir (Mazza ve Cottrell, 1999; Miller ve Yu, 2004). Her bir bileĢik grubu farklı farmakolojik etkilere sahiptir (Barrett, 2003).
Ekinezyanın E. angustifolia, E. pallida ve E. purpurea olmak üzere üç türü tıbbi olarak değerlendirilmektedir. Bu türlerin kimyasal bileĢimleri açısından birbirinden farklılıkları olup; karakteristik bileĢen, miktar ve lokalizasyonu bitkinin yaĢına, bitki kısımlarına (Çizelge 2.1) ve çevresel faktörlere bağlı olarak değiĢiklik göstermektedir (Binns ve ark., 2002b; Toselli ve ark., 2009).
Çizelge 2.1. E. purpurea, E. angustifolia ve E. pallida‘da biyoaktif bileĢiklerin lokalizasyonu
E. purpurea E. angustifolia E. pallida
Çiçek Tablası Kafeik Asit Türevleri
Kafeik Asit Türevleri Kafeik Asit Türevleri
Alkamitler Alkamitler Alkamitler
Ekinakosit Ekinakosit
Yaprak Sapı Kikorik Asit Alkamitler Kikorik Asit Alkamitler ?
Yaprak
Kikorik Asit Kikorik Asit Kikorik Asit
Kaftarik Asit Kaftarik Asit Alkamitler
Sinarin Sinarin
Kök Alkamitler Ekinakosit Ekinakosit
Sinarin Sinarin Alkamitler
*Kalın yazılmıĢ bileĢikler yüksek konsantrasyonlarda, italik yazılan bileĢikler ise eser miktarda bulunmaktadır (Parsons ve ark., 2018).
Ekinezyanın yararlı etkisine neden olan tüm fitokimyasal mekanizmalar halen büyük ölçüde bilinmemekle birlikte, üst solunum yolu enfeksiyonlarında bağıĢıklık sistemini uyarıcı aktiviteleri açıklamaya yönelik çok sayıda araĢtırma mevcuttur (Toselli ve ark., 2009). Yapılan bu araĢtırmalarda, ekinezyanın bağıĢlık sistemini, T hücresi üretimini, fagositozu, lenfositik aktiviteyi ve hücresel solunumu (antioksidan) uyararak (Barrett, 2003) ayrıca hiyalüronidaz enzim salgılanmasını önleyerek sağladığı belirlenmiĢtir (Bergeron ve ark., 2002). Ekinezyanın, canlılarda doğuĢtan gelen bağıĢıklık fonksiyonunu aktivite ettiği, aktif bileĢenlerinin vücuttaki birçok hücrenin çekirdeğinde bağıĢıklık uyarıcı ve düzenleyici etkisi olduğunu belirtilmiĢtir (Zhai ve ark., 2007). Ayrıca, sindirim fonksiyonlarını güçlendirmesi ve vücutta bulunan fagosit hücreleri çoğaltarak kanda bulunan patojenlere karĢı etkili olduğunu belirten araĢtırmalar da bulunmaktadır (Roesler ve ark., 1991).
Ekinezya cinsinin en önemli bileĢenlerini fenolik bileĢikler, bunlar içinde de kafeik asit türevleri oluĢturmaktadır. Kikorik asit, ekinakosit, klorojenik asit, kaftarik asit, kafeik asit ve sinarin, ekinezyada en sık rastlanan kafeik asit türevleri olup (ġekil 2.2) miktarları türlere ve bitki kısımlarına göre değiĢmektedir (Barnes ve ark., 2005) (Çizelge 2.2). Örneğin, klorojenik asit, tüm ekinezya türlerinde minimum miktarda bulunurken, kikorik ve kaftarik asit ise E. purpurea'nın karakteristik bileĢenleri; sinarin ise sadece E. angustifolia köklerine özgü bir bileĢik olarak belirlenmiĢtir (Wills ve Stuart, 1999). Ekinakosit, E. pallida ve E. angustifolia'nın karakteristik bileĢeni iken E.
purpurea türünde bulunmamaktadır (Pietta ve ark., 1998). Ayrıca, bu bileĢiklerin
dağılımının, tek bir bitkinin organları ve dokuları arasında da farklı olduğu bildirilmiĢtir (Lin ve ark., 2011). E. pallida'nın yaprakları kikorik ve kaftarik asitlerce daha zengin, köklerinde ise ekinakosit daha baskındır. E. angustifolia'da en yüksek sinarin ve ekinakosit içeriği köklerde tespit edilmiĢtir (Kabganian ve ark., 2003). E. purpurea‘da
ise fenolik profil daha az çeĢitlilik göstermekte, kikorik ve kaftarik asit çoğunlukla köklerde ve çiçeklerde bulunmaktadır (Qu ve ark., 2005). Özellikle kikorik asit miktarı bitkinin geliĢme evresiyle yakından ilgili olup, çiçeklenme döneminde ya da tamamen olgunlaĢmıĢ bitkilerde daha yüksek miktarlarda elde edilmiĢtir (Letchamo ve ark., 1999; Stuart ve Wills, 2000)
Çizelge 2.2. Farklı Ekinezya türlerinde kafeik asit türevlerinin miktarı (Murthy ve ark., 2014)
Bitki Türü Bitki Kısmı BileĢen ve Miktarı (%) Özellik
E. purpurea Toprak üstü Kikorik asit (1.2–3.2) Ekinakosit mevcut değildir.
E. purpurea Kök Kikorik asit (0.14–2.05) Ekinakosit mevcut değildir.
E. angustifolia Kök Ekinakosit (0.5–1.0)
Sinarin
Sinarin E. angustifolia‘nın karekteristik bileĢiğidir.
E. pallida Kök Ekinakosit (0.5–1.0) Ekinakosit E. pallida‘nın temel bileĢiğidir.
Kafeik asit türevlerinin her biri önemli farmakolojik özelliklere sahip olmakla birlikte, özellikle kikorik asit ve ekinakosit bu grubun en önemli bileĢikleridir (Murthy ve ark., 2014). Kikorik asitin, HIV-1 (insan immün yetmezlik virüsü) integraz enzimini engellemesi ve fagosit aktiviteyi teĢvik etmesi sonucu immunostimulatör ve antiviral aktivitelere sahip olduğu belirlenmiĢtir (Bauer ve ark., 1989; Lin ve ark., 1999). Ayrıca, kikorik asit ve ekinakosit reaktif oksijen türlerine karĢı kollojeni koruma, antihiyalüronidaz ve yüksek serbest radikal temizleme aktiviteleri sayesinde antioksidan ve antienflamatuar etkiler göstermekte, böylece UV ıĢınların deride oluĢturduğu zararları koruyucu ve tedavi edici olarak kullanılabilmektedir (Facino ve ark., 1995; Speroni ve ark., 2002; Mistrikova ve Vaverkova, 2006). Klorojenik asitin bağırsakta glikoz emilimini inhibe ederek Ģeker hastalığını önlediği ve N-nitroso bileĢiklerinin oluĢturduğu kanserojenik etkileri hafifleterek antikanserojenik etkiler yaptığı bildirilmektedir (Kono ve ark., 1995; Shibata ve ark., 1999; Kasai ve ark., 2000). Sinarinin ise sindirim sistemini düzenleyici koleretik ve kolagog özelliklere sahip olduğu tespit edilmiĢtir (Speroni ve ark., 2002).
Sekil 2.2. Ekinezya türlerinde bulunan kafeik asit türevlerinin kimyasal yapıları
2.4. Ekinezya Türlerinde Doku Kültürü ÇalıĢmaları
ÇalıĢmamızın amaçlarından biri ekinezyada etkili ve tekrar edilebilen bir rejenerasyon sistemi oluĢturmak olduğu için, bugüne kadar farklı ekinezya türlerinde yapılan araĢtırmalar sonucu belirlenen hücre ve doku kültürü protokolleri incelenmiĢ ve referans olarak çalıĢmamızda değerlendirilmiĢtir.
E. purpurea L. steril fidelerinden alınan yaprak sapı eksplantları, farklı
konsantrasyonlarda BAP veya TDZ büyüme düzenleyicilerinin IAA, NAA veya 2,4-D ile kombinasyonlarını içeren besin ortamlarında kültüre alınmıĢtır. Kültür süresi sonunda, epidermis dokudan direk somatik embriyoların oluĢumu ve kallus dokularından sürgünlerin geliĢmesi olmak üzere iki farklı rejenerasyon formu gözlemlenmiĢtir. En yüksek eksplant baĢına sürgün sayısı (8.1 adet) 2.5 µM BAP içeren besin ortamlarında elde edilmiĢ, ayrıca BAP içeren besin ortamlarında eksplantların
Kaftarik asit Klorojenik asit
Kafeik asit Sinarin
Ekinakosit
yüzeyinde geç kotiledon evrede somatik embriyolar gözlemlenmiĢtir. BAP ile NAA veya 2,4-D büyüme düzenleyicilerinin kombinasyonlarını içeren ortamlarda rejenerasyon uyarımı gerçekleĢmediği tespit edilirken, 1 µM IAA ve 0.5 µM TDZ içeren ortamda eksplant baĢına ortalama sürgün sayısı 4.9 adet olmuĢtur. Bu çalıĢmanın sonucunda E. purpurea‘nın yaprak sapı eksplantlarından rejenerasyon sistemi oluĢturulmuĢ olup böylece ticari anlamda yüksek kalitede kitlesel üretimi kolaylaĢtıracak bir protokol belirlenmiĢtir (Choffe ve ark., 2000b).
E. purpurea’nın yaprak eksplantlarının kallus kültürlerinden organogenesis
aracılığıyla etkili bir bitki rejenerasyon sisteminin araĢtırıldığı çalıĢmada, farklı konsantrasyonlarda (0, 0.44, 2.22, 4.44, 8.88, 17.76, 31.08 µM) BAP ve (0, 0.054, 0.54, 2.69 µM) NAA büyüme düzenleyicileri kombinasyonlarını içeren MS besin ortamı kullanılmıĢtır. En yüksek sürgün rejenerasyon yüzdesi (%100) ve eksplant baĢına sürgün sayısı (7.7 adet) 4.44 µM BAP ve 0.054 µM NAA içeren MS besin ortamında elde edilmiĢtir. Elde edilen sürgünler, farklı konsantrasyonlarda (0, 0.49, 2.46, 4.90, 9.8 µM) IBA içeren MS besin ortamında köklendirilmiĢ, en yüksek köklenme oranı (%97) büyüme düzenleyicisi içermeyen besin ortamında elde edilmiĢtir. Bu çalıĢma, E.
purpurea‘nın yaprak eksplantlarının adventif sürgün rejenerasyonu için uygun olduğunu
ve bu yöntemle bitkinin mikroçoğaltımının rahatlıkla yapılabileceğini göstermektedir (Koroch ve ark., 2002).
E. purpurea‘nın hipokotil eksplantlarından farklı konsantrasyonlarda IAA, IBA
ve NAA oksinlerini içeren MS besin ortamında kök organogenesis uyarımının araĢtırıldığı çalıĢmada; IAA büyüme düzenleyicisinin IBA‘ya göre daha az etkili olduğu, NAA‘nın ise kök organagenesis uyarımında etkisiz olduğu bulunmuĢtur. Bu çalıĢma, steril bitki materyalinden mikroçoğaltım sisteminin kurulmasını sağlamanın yanı sıra endüstriyel anlamda yüksek kalitede E. purpurea kök dokularının kitlesel üretimine öncülük sağlamaktadır (Choffe ve ark., 2000a).
E. pallida‘nın yaprak eksplantlarından adventif sürgün uyarımı sağlayarak, bitki
rejenerasyonunun araĢtırıldığı çalıĢmada, kallus ve sürgün kültürleri farklı konsantrasyonlarda (0, 4.4, 8.9, 17.8, 26.6, 35.5 µM) BAP ve (0, 0.11, 0.27 µM) NAA büyüme düzenleyicileri kombinasyonlarını içeren MS besin ortamında elde edilmiĢtir. En yüksek sürgün rejenerasyon yüzdesi (%63) ve eksplant baĢına sürgün sayısı (2 adet), 26.6 µM BAP ve 0.11 µM NAA içeren besin ortamında tespit edilirken, rejenerent sürgünlerin köklendirilmesi IBA içeren/içermeyen MS besin ortamında baĢarılı olmuĢtur. Bu çalıĢma, E. pallida‘nın yaprak eksplantlarının adventif sürgün
rejenerasyonu için uygun olduğunu ve bu yöntemle bitkinin mikroçoğaltımının rahatlıkla yapılabileceğini göstermektedir (Koroch ve ark., 2003).
E. purpurea‘nın yaprak eksplantlarının farklı konsantrasyonlarda BAP ve NAA
içeren WPM besin ortamında kültüre alındığı çalıĢmada, yapraktan direk sürgün uyarımı araĢtırılmıĢtır. 2.5 mg/l BAP ve %5 hindistan cevizi sütü içeren besin ortamında direkt sürgün uyarımı gerçekleĢmiĢ ve en yüksek sürgün sayısına (15 adet) ulaĢılmıĢtır. 0.5 mg/l NAA ve 2.5 mg/l BAP içeren besin ortamında ise en yüksek kallus uyarım oranı (% 53.33) elde edilmiĢtir. Elde edilen sürgünler, farklı konsantrasyonlarda (0, 0.5, 1.0, 1.5, 2.0 mg/l) IBA içeren besin ortamında köklenmeye alınmıĢ, en yüksek köklenme oranı (%100) 1.5 mg/l IBA konsantrasyonunda elde edilmiĢ ve sonrasında bu bitkilerin baĢarılı bir Ģekilde aklimatizasyonu yapılmıĢtır (Mechanda ve ark., 2003).
E. purpurea‘da kallus oluĢumu ve direkt sürgün uyarımı ile rejenerasyon
protokolünün araĢtırıldığı çalıĢmada, steril fidelerden alınan hipokotil eksplantları 1.0-3.0 mg/l NAA + 1.0-2.0 mg/l KIN ve 0.5-1.0 mg/l 2,4-D + 1.5-2.0 mg/l KIN içeren MS besin ortamlarında kültüre alınmıĢtır. Kültür süresi sonunda eksplantlardan, NAA+KIN içeren ortamlarda çoğunlukla direkt sürgün ve kök uyarımı gerçekleĢirken, 2,4-D+KIN içeren ortamlarda ise çoğunlukla kallus uyarımı sağlanmıĢtır. NAA+KIN içeren ortamlarda elde edilen eksplantlardan olgun bitki üretimi gerçekleĢtirilmiĢ, en yüksek bitki üretim oranı (%100) 1.0 mg/l NAA+ 1.0 mg/l KIN içeren ortamda sağlanmıĢtır (Coker ve Camper, 2000).
E. purpurea‘da seri üretim için TDZ uyarımlı mikroçoğaltım protokolünün
araĢtırıldığı çalıĢmada, 2,4-D veya dikamba içeren MS besin ortamlarında yaprak eksplantlarından sadece kallus geliĢimi ve kök organogenesisi elde edilmiĢ, besin ortamına TDZ ilavesi ile kallus kültürlerinden rejenerasyon sağlanmıĢtır. En yüksek rejenerasyon oranı 2.5 μM ve daha üzeri TDZ içeren besin ortamlarında elde edilmiĢtir (Jones ve ark., 2007).
Seçkin bireylerin germplazmanın korunması ve hızlı klonal çoğaltım amacıyla üç önemli ekinezya türünün (E. purpurea, E. pallida ve E. angustifolia) mikroçoğaltım protokolünün araĢtırıldığı çalıĢmada, besin ortamına BAP ilavesi sürgün çoğaltımını artırırken, kök oluĢumunu üç türde de engellemiĢtir. Sürgün çoğaltım oranı, düĢük miktarda BAP içeren ortamlarda daha az sayıda elde edilmiĢtir. Alt kültüre alma sıklığını 4 haftadan 2 haftaya düĢürmek, alt kültür baĢına sürgün sayısını 1.4‘den 1.8‘e sürgüne ve 12 hafta sonunda alt kültür baĢına üretilen toplam sürgün sayısını 2.8'den 23.9'a yükseltmiĢtir. Köklenme, E. purpurea sürgünlerinde yüksek oranda (%97), E.
angustifolia (%32) ve E. pallida (%3) sürgünlerinde ise oldukça düĢük oranda
oluĢmuĢtur. E. pallida ve E. angustofilia türlerinde yapılacak kök uyarımı araĢtırmaları ile bu türlerin hastalıksız germplazının mikroçoğaltımı ve koruma altına alınması etkili bir Ģekilde yapılabilecektir (Harbage, 2001).
Üstün tarla performansı gösteren altı farklı E. purpurea genotipinde, adventif sürgün reienerasyonu ve aksiller sürgün çoğaltımı aracılığıyla çoklu sürgün üretimi için en uygun BAP konsantrasyonlarının belirlenmesinin araĢtırıldığı çalıĢmada, altı genotip ve farklı eksplant tipleri (yaprak, yaprak sapı, kök) arasında en uygun BAP konsantrasyonları bakımından büyük farklılıklar gözlemlenmiĢtir. En uygun BAP konsantrasyonunda, altı genotipten beĢinde aksillar sürgün çoğaltımı, adventif sürgün rejenerasyonundan daha etkili bulunmuĢtur. Elde edilen verilere dayanarak, çalıĢılan genotipler arasında üçünde etkili bir mikroçoğaltım sistemi belirlenmiĢtir (Li ve ark., 2013).
E. purpurea’da etkili bir mikroçoğaltım protokolünün araĢtırıldığı çalıĢmada, 0.3
mg/l BAP ve 0.01 mg/l NAA içeren MS besin ortamında, bitkinin steril fidelerinden alınan yaprak, yaprak sapı ve kök eksplantlarından adventif sürgünler elde edilmiĢ, elde edilen adventif sürgünler 0.01 mg/l NAA içeren MS besin ortamında köklendirilmiĢtir. Farklı olgunluktaki steril fideler alınan eksplantların farklı adventif sürgün oluĢturma yeteneğine sahip olduğu, 2.5 aylık fideler alınan eksplantlarda en yüksek adventif sürgün frekansına ulaĢıldığı tespit edilmiĢtir. Bu mikroçoğaltım protokoli; ile bir yıl içinde en az bir milyon bitkinin elde edilebileceği belirtilmiĢtir (Dahanayake ve ark., 2011).
Ticari açıdan önemli dört ekinezya türünde (E. angustifolia, E. pallida, E.
paradoxa ve E. purpurea) kitlesel üretim için etkili in vitro protokollerin
geliĢtirilmesinin amaçlandığı çalıĢmada, aksillar sürgün çoğaltımı ve somatik embriyogenesis araĢtırılmıĢtır. Steril nodlar, farklı konsantrasyonlarda (0.5, 1, 2, 4 μM) BAP, KIN, IBA ve PBA içeren MS besin ortamlarında kültüre alınmıĢ, E. pallida ve E.
purpurea türlerinde 4 haftalık kültür süresinin sonunda, 2 μM BAP içeren ortamda
çoklu sürgün üretiminin 3 kat arttığı tespit edilmiĢtir. Elde edilen sürgünlerin köklenmesi, oksin içeren ortamlara kıyasla büyüme düzenleyicisi içermeyen MS besin ortamında daha yüksek oranda (sırasıyla %56, %50, %48, %94) gerçekleĢtirilmiĢtir. Tüm türlerin hipokotil eksplantları, dikamba ve 2,4-D içeren besin ortamlarında kültüre alınmıĢ ve tamamında direk somatik embriyogenesis (sırasıyla %75, %85, %25, %20) elde edilmiĢtir. Elde edilen somatik embriyolar sitokinin içeren MS besin ortamında
çimlendirilmiĢ, büyüme düzenleyicisi içermeyen MS besin ortamında ise 6 haftalık kültür süresinin sonunda sağlıklı fideler elde edilmiĢtir. Bu araĢtırmada belirlenen in
vitro protokoller, ticari açıdan önemli ekinezya türlerinde hızlı çoğaltım
gerçekleĢtirilmesinin yanı sıra genetik transformasyon çalıĢmaları için de imkan sağlayacağı düĢünülmektedir (Lakshmanan ve ark., 2002).
Zebarjadi ve ark. (2011) tarafından yapılan çalıĢmada, E. purpurea‘nın kotiledon ve hipokotil eksplantlarından organogenesis aracılığıyla in vitro bitki rejenerasyonunun optimizasyonu araĢtırılmıĢtır. Eksplantlar farklı konsantrasyonlar ve kombinasyonlarda BAP ve NAA içeren MS besin ortamında 5 hafta boyunca kültüre alınmıĢtır. Kültür süresi sonunda, en yüksek kallus oluĢum oranı eksplant tipine bağlı olarak farklılık göstermiĢ, kotiledon eksplantında 0.2 mg/l BAP (%97), hipokotil eksplantında ise 0.2 mg/l BAP ve 0.6 mg /L NAA (%91) içeren besin ortamlarında elde edilmiĢtir. Kalluslardan sürgün rejenerasyon yüzdesi açısından eksplant tipleri arasında farklılık görülmemiĢ, hem kotiledon hem hipokotil eksplantlarında en yüksek sürgün rejenerasyon yüzdesi (sırasıyla %31.5 ve %32.5) ve eksplant baĢına sürgün sayısı (sırasıyla 5.2 ve 5.3), 0.4 mg/l BAP içeren besin ortamından elde edilmiĢtir. Elde edilen sürgünler, 2 mg/l IBA içeren ve içermeyen MS besin ortamında köklendirilmiĢtir. Bu çalıĢmanın sonucunda, E. purpurea kotiledon ve hipokotil eksplantlarının kallus uyarımı ve sürgün oluĢumu için önemli bir potansiyele sahip olduğu ve mikroçoğaltımı için önemli bir protokol belirlenmiĢtir.
2.5. Ekinezya Türlerinde Doku Kültürü ile Sekonder Metabolit Üretiminin AraĢtırılması ile Ġlgili ÇalıĢmalar
Sekonder metabolit kavramı ilk defa Kossel (1891) tarafından primer metabolitlerin karĢıtı olarak tanımlanmıĢ, Theis ve Lerdau (2003) ise sekonder metabolitleri bitkiler tarafından üretilen, fotosentez ürünü olmayıp, birtakım fizyolojik mekanizmaların fonksiyonlarıyla oluĢan ve fotosentez ya da solunum gibi hayati fizyolojik olaylar için mutlak gerekli olmayan maddeler Ģeklinde tarif edilmiĢtir.
Sekonder metabolitler, bitkide savunma, korunma, hayatta kalma, nesillerini sürdürme gibi çevresel koĢullara uyum faaliyetleri esnasında üretilmekte olup bazıları geniĢ miktarlarda üretilirken, bazılarının üretimi ise sınırlı düzeyde kalmakta hatta bazı metabolitler sadece belirli türlerde üretilmektedir. Çoğunlukla bitkilerin belli
organlarında bulunurlar ve bitkinin belli bir geliĢim periyodu süresince üretilirler (Verpoorte ve ark., 1999; Sökmen ve Gürel, 2001).
Sekonder metabolitlerin kültürü yapılan bitkilerden elde edilmesi sırasında; metabolitlerin miktar ve kalitesinin iklimsel koĢullardan etkilenmesi, bitkilerde belli geliĢim evrelerinde ve çok az miktarda sentezlenmeleri, bitkilerin kültüre alınmalarında baĢarının düĢük oluĢu, ekonomik miktarlarda etkili madde üretimi için geniĢ tarım alanlarına ihtiyaç duyulması gibi çeĢitli zorluklarla karĢılaĢılmaktadır (Erkoyuncu ve Yorgancılar, 2016). Ayrıca sekonder metabolitler çoğunlukla doğal florada bulunan bitkilerin toplanması sonucu elde edilmekte, bu da bitkilerin nesillerinin zamanla yok olma tehlikesini ortaya çıkarmaktadır.
Sekonder metabolitlerin bitki hücre ve doku kültürleri ile üretilmesi, geleneksel metotlara kıyasla belirgin avantajlara sahiptir. Bu avantajlar; ilgili metabolitin çevresel faktörlerden bağımsız olarak kontrollü Ģartlarda üretilebilmesi, hastalık ve zararlılardan arındırılmıĢ bitkisel materyalin elde edilmesi, herhangi bir bitkinin, ister tropik ister subtropik kökenli olsun, in vitro kültüre alınabilmesi, kültür Ģartlarının sekonder metabolit üretimini artıracak Ģekilde optimize edilebilmesi (Rao ve Ravishankar, 2002) Ģekilde özetlenebilir. Sekonder metabolit ürünlerinin in vitro koĢullarda üretimi aynı zamanda kontrollü koĢullarda bu ürünlerin biyokimyasal ve metabolik süreçlerinin yakından incelenmesine olanak sağlamaktadır (Karuppusamy, 2009). Hücre hatlarının (varyantlarının), arzu edilen metaboliti yüksek oranda üreten kültürlerden seçimi ve bu hatlardan, özellikle de tek hücre kültürleri yapılarak üstün nitelikli bitkilerin rejenerasyonu, bitki hücre ve doku kültürü tekniklerinin sunduğu bir baĢka uygulama alanıdır (Sökmen ve Gürel, 2001).
Birçok çevre ve beslenme faktörünün sekonder metabolitlerin biyosentez yolunda etkili olduğu bilinmektedir. Bitki doku ve hücre kültürlerinde, kültür ortamında yer alan beslenme faktörleri; karbon, fosfor, azot kaynakları ve diğer makro elementler ile bitki büyüme düzenleyicileri, yani oksinler, sitokininler, gibberellinler ve brassionosteroidler hem sekonder metabolitlerin oluĢumuna, hem de büyümeye etki etmektedir. Bu faktörlerin her biri kültürü yapılan bitkiye, kültür tipine, hatta kültürün yaĢına göre farklı etkilerde bulunmaktadır (Gundlach ve ark., 1992; Sökmen ve Gürel, 2001; Bhagyalakshmi ve ark., 2004). Kültür ortamındaki besin elementlerinin ve büyüme düzenleyicilerin kompozisyonu, hem biyokütle artıĢını sağlamak hem de istenilen metabolitlerin birikmesi amacı ile optimize edilmelidir. Bu anlamda, bitki
büyüme düzenleyicileri biyosentetik yolların uygun bir Ģekilde teĢvik edilmesi amacıyla oldukça önemlidir.
Jones ve ark. (2009b) yaptıkları araĢtırmada, E. purpurea‘da in vitro koĢullarda yetiĢtirilen bitkilerde kaftarik asit ve kikorik asitin miktarlarını artırmak amacıyla GA3
ile triazollerin (paklobutrazol, unikonazol) etkilerini incelemiĢlerdir. ÇalıĢmada, farklı konsantrasyonlarda (50, 100 ve 150 μM) GA3, paklobutrazol ve unikonazol ayrı ayrı ve
kombinasyon halinde uygulanmıĢ, elde edilen in vitro bitkilerin kök ve sürgünlerindeki kaftarik ve kikorik asit miktarları HPLC ile belirlenmiĢtir. En yüksek kaftarik (10 mg/g) ve kikorik asit (36 mg/g) miktarları in vitro bitkilerin köklerinde ve 100 μM GA3 ve 100
μM paklobutrazol içeren besin ortamında tespit edilmiĢtir. Ayrıca uygulamaların in
vitro bitkilerin köklerinde kikorik asit ve kaftarik asit miktarlarını artırdığı ancak
sürgünlerdeki kaftarik asit ve kikorik asit miktarlarına çok etki etmediği belirlenmiĢtir. Abbasi ve ark. (2012) tarafından yapılan çalıĢmada, E. purpurea L. saçak kök kültürlerinde, GA3‘ün kafeik asit türevlerinin üretimini artırmadaki etkinliğinin
belirlenmesi amaçlanmıĢtır. 0.005 ila 1.0 μM arasında değiĢen 8 farklı konsantrasyonda GA3‘ün kullanıldığı araĢtırma sonucunda, 0.025 μM GA3 uygulamasında, en yüksek
kikorik asit (33 mg/g), kaftarik asit (8 mg/g), kafeik asit (0.01 mg/g) ve klorojenik asit (7 mg/g) miktarları elde edilmiĢ, ayrıca PAL ve serbest radikal temizleme aktivitelerinde de artıĢlar gözlemlenmiĢtir. AraĢtırmada GA3‘ün E. purpurea L.
bitkilerinde sekonder metabolit üretimini artırmak için çok iyi bir ajan olduğu, fakat konsantrasyonların çok iyi belirlenmesi gerektiği vurgulanmıĢtır.
Wu ve ark. (2006), E. angustifolia‘nın adventif kök kültüründe, farklı oksin tipleri (IAA, IBA, NAA) ve konsantrasyonları (1.0, 2.0, 4.0, 6.0 mg/l) ile farklı sakkaroz (%1, %3, %5, %7, %9), amonyum/nitrat içeriği (0:40, 0:35, 0:30, 5:25, 10:20, 15:15, 20:10, 25:5, 30:0), besin ortamı gücü (1/4, 1/2, 3/4, 1/1, 3/2, 2/2) ve pH (4, 5, 6, 7, 8, 9) seviyelerinin, biyokütle artıĢı ve toplam fenol ve flavonoidlerin birikimi üzerine etkilerini araĢtırmıĢlardır. AraĢtırmanın sonucunda, en fazla biyokütle artıĢı ile toplam fenol ve flavonoid içeriğine, %5 sakkaroz, 5:25 (mM) amonyum/nitrat, 2 mg/l IBA içeren, pH:6.0 olan 1/2 MS besin ortamında ulaĢılmıĢtır. Besin ortamı içeriği optimizasyonunun sekonder metabolit üretimine önemli etkide bulunduğu tespit edilmiĢtir.
E. purpurea L.‘nın advendif kök kültürlerinde kafeik asit türevlerinin üretimini
artırabilmek amacıyla besin ortamı optimizasyonunu araĢtırıldığı baĢka bir çalıĢmada, farklı kuvvetlerde (¼ MS, ½ MS, ¾ MS ve MS) besin ortamlarında adventif kök
kültürlerinin üretimi sağlanmıĢ, 5 haftalık kültür süresinin sonunda adventif köklerdeki biyokütle, fenolik ve flavonoidler ile kafeik asit türevlerinin miktarları tespit edilmiĢtir. En yüksek kaftarik asit (4.35 mg/g), klorojenik asit (4.87 mg/g) ve kikorik asit (29.06 mg/g) ile optimum biyokütle, fenolik ve flavonoid miktarları ½ MS ortamında elde edilmiĢtir. ÇalıĢmanın bir sonraki aĢamasında, ½ MS besin ortamı, kültür süresi içerisinde 2. ve 3. haftalarda yenilenmiĢ, ancak bu tazeleme iĢlemi, adventif köklerin biyükütle, fenolik ve flavonoid miktarlarında düĢüĢe sebep olurken kafeik asit türevlerinin miktarlarında ciddi bir değiĢiklik oluĢturmamıĢtır (Wu ve ark., 2007b)
Bitki doku kültürlerinde, sekonder metabolitlerin üretimi genellikle hücre çoğalması ve farklılaĢmasına bağlı olarak artıĢ göstermektedir (George ve ark., 2008). Özellikle, belli bir organ (kök, sürgün veya embriyo benzeri yapılar) oluĢturmak üzere farklılaĢan kültürlerde sekonder metabolit birikiminin daha fazla olduğu gözlenmiĢtir (Sökmen ve Gürel, 2001). Özellikle bu amaçla kallus kültürleri, adventif kök kültürleri ve saçak kök kültürleri in vitro koĢullarda sekonder metabolitlerin üretiminde sıklıkla kullanılmaktadır.
E. purpurea L.‘nın yaprak eksplantlarından Agrobacterium rhizogenes aracılığla
elde edilen saçak kök kültürlerinin kafeik asit üretme kapasitelerinin araĢtırıldığı çalıĢmada, en yüksek biyokütle 30 g/l sakkaroz içeren MS besin ortamında 40 günlük kültür süresi sonunda elde edilmiĢtir. Elde edilen saçak kök kültürlerinde yapılan HPLC analizleri sonucunda önemli kafeik asit türevlerinin varlığı tespit edilmiĢ; kikorik asit (19.21 mg/g), kaftarik asit (3.56 mg/g) ve klorojenik asit (0.93 mg/g) miktarları belirlenmiĢtir (Liu ve ark., 2006).
Ekinezya'nın üç türünde (E. purpurea L., E. pallida ve E. angustifolia),
Agrobacterium uygulamasıyla saçak kök elde edildiği araĢtırmada, saçak kök
kültürünün optimizasyonu hem büyüme hem de alkamit üretimini eĢ zamanlı olarak arttırmak için uygulanmıĢtır. %3 sakkaroz içeren ½ B5 ortamı, saçak kök üretiminde, test edilen diğer ortama oranla iki kat fazla etkili olduğu tespit edilmiĢtir. Alkamit üretimini jasmonik asit ilavesi birkaç kat artırırken, IBA ilavesi yanıt vermemiĢtir. Bu çalıĢmanın sonuçları, Ekinezya'nın saçak kök kültürlerinin alkamitlerin in vitro üretimi için etkinliğini ve optimum verim için uygun Ģartları ortaya koymuĢtur (Romero ve ark., 2009).
E. angustifolia ve E. pallida türlerinin kallus ve rejenerent sürgünlerinde, kafeik
asit türevlerinin üretimini ve birikimini artırmak için etkili bir protokolün araĢtırıldığı çalıĢmada, farklı konsantrasyonlarda (1.0, 3.0, 5.0 mg/l) 2,4-D ve 1.0 mg/l BAP içeren
MS besin ortamında yaprak, yaprak sapı ve kök eksplantlarından kallus ve farklı konsantrasyonlarda (1.0, 3.0, 5.0 mg/l) BAP ve 1.0 mg/l IAA içeren MS besin ortamında ise rejenarent sürgünlerin üretimi gerçekleĢtirilmiĢtir. Her iki türde de, kallus ve rejenerent sürgünlerin en yüksek değeri sırasıyla yaprak, yaprak sapı ve kök eksplantlarında elde edilmiĢtir. Kallus üretimi için 3.0 mg/l 2,4-D ve 1.0 mg/l BAP, rejenerant sürgünlerin çoğalması için 1.0 mg/l IAA ve 3.0 mg/l BAP büyüme düzenleyicilerini içeren besin ortamları optimum olarak belirlenmiĢtir. Elde edilen kalluslarda ve rejenarent sürgünlerde kafeik asit türevleri (kaftarik, kikorik ve klorojenik asit) HPLC ile tespit edilip, belirlenmiĢtir. En yüksek kaftarik asit, klorojenik asit ve kikorik asit miktarı (sırasıyla 3.94, 5.94 ve 24.7 mg/g), sırasıyla yaprak ve yaprak sapı eksplantlarına kıyasla, kök eksplantlarından uyarılmıĢ sürgünlerde kaydedilmiĢtir (Taha ve ark., 2009).
Üç farklı ekinezya türünde (E. purpurea L., E. pallida ve E. angustifolia), ekinezya ekstraktlarının temel aktif bileĢiklerden biri olan kikorik asit üretimi için biyokütle elde edilmesi amacıyla in vitro mikroçoğaltım protokollerinin araĢtırıldığı çalıĢmada; ½ MS besin ortamında tohumlar çimlendirilip steril fideler elde edilmiĢtir. Farklı konsantrasyonlarda NAA, IBA, 2-izo-penteniladenin (2iP) ve BAP büyüme düzenleyicilerini içeren MS besin ortamında sürgün çoğaltımı gerçekleĢtirilmiĢtir. En fazla sürgün sayısı E. purpurea L. türünde (7.1 sürgün/nodal eksplant) 0.1 mg/l 2iP ve 0.1 mg/l IBA, E. pallida (5.4 sürgün/ nodal eksplant) ve E. angustifolia (3.9 sürgün/nodal eksplant) türlerinde ise 1 mg/l 2iP ve 0.1 mg/l IBA içeren besin ortamlarında elde edilmiĢtir. Ayrıca, her üç türün yaprak eksplantları kallus uyarımı için 2.0 mg/l 2,4-D ve 0.1 mg/l BAP içeren besin ortamında kültüre alınmıĢ, elde edilen kalluslarda kikorik asit miktarı belirlenmiĢtir. In vitro koĢullarda elde edilen sürgünler ve kallus dokularındaki kikorik asit miktarı HPLC ile tespit edilmiĢtir. Her üç türde de en yüksek kikorik asit seviyeleri sürgünlerde (E. purpurea L. türünde 0.58 mg/g, E.
pallida türünde 0.73 mg/g ve E. angustifolia türünde 0.39 mg/g) belirlenmiĢtir
(Butiuc-Keul ve ark., 2012).
Sekonder metabolitlerin bitkilerdeki en önemli rollerinden biri stres tepkilerine yanıt olarak geliĢtirilmiĢ olmalarıdır (Grassmann ve ark., 2002). Bitki hücre ve doku kültürlerinde sekonder metabolit üretimini artırmak, aynı zamanda yüksek konsantrasyonlarda üretimi kısa sürede sağlamak amacıyla biyotik ve abiyotik elisitörler (uyarıcılar) kullanılmaktadır (Barz ve ark., 1988). Elisitörün etkisinin, in vitro kültür
tipine, uygulanan elisitörün konsantrasyonuna ve süresine bağlı olarak değiĢiklik gösterdiği bildirilmiĢtir (Gadzovska ve ark., 2013).
E. purpurea L.’nın adventif kök kültürlerinde elisitör olarak nitrik oksidin
sekonder metabolit üretimine etkisinin araĢtırıldığı çalıĢmada, farklı konsantrasyonlarda (0, 50, 100 ve 250 μM) nitrik oksit üreticisi olan sodyum nitroprusit (SNP) çözeltileri besin ortamına ilave edilmiĢtir. En yüksek kaftarik, klorojenik ve kikorik asit miktarları (sırasıyla 3.71, 4.95, 34.89 mg/g kuru ağırlık) 100 μM SNP uygulamasında elde edilirken, 250 μM SNP uygulamasının ise pro-oksidan etki oluĢturarak kafeik asit türevlerinin üretimini olumsuz etkilediği belirlenmiĢtir. Ayrıca, en yüksek fenolik ve flavonoid miktarları da yine 100 μM SNP uygulamasında elde edilmiĢ ve optimum konsantrasyon olarak belirlenmiĢtir (Wu ve ark., 2007d).
Cui ve ark. (2013) yaptıkları çalıĢmada, E. angustifolia’nın adventif kök kültürlerinde, farklı besin ortamı gücü (1/4, 1/2, 3/4, 1/1 ve 2/1 MS) ve sakkaroz konsantrasyonlarının (% 0, %1, %3, %5, %7, %9) biyokütle artıĢı ve kafeik asit türevlerinin birikimi üzerindeki etkilerini araĢtırmıĢlardır. En yüksek biyokütle (46.83 g/l) ile klorojenik asit (2.26 mg/g), ekinakosit (4.66 mg/g), sinarin (1.57 mg/g) ve kikorik asit (1.57 mg/g) miktarları, %5 konsantrasyonda sakkaroz içeren ¼ MS besin ortamında elde edilmiĢtir. ÇalıĢmanın bir sonraki aĢamasında belirlenen besin ortamı gücü ve sakkaroz konsantrasyonunda elde edilen adventif köklere, 1 hafta boyunca 100 µM metil jasmonat elisitör olarak uygulanmıĢ ve biyokütle ile kafeik asit türevlerinin miktarları belirlenmiĢtir. Biyokütle (50.88 mg/l) ile klorojenik asit (5.54 mg/g), ekinakosit (12.30 mg/g), sinarin (6.25 mg/g), kikorik asit (5.45 mg/g) olarak tespit edilmiĢ ve elisitör uygulamasıyla biyokütle ve kafeik asit türevlerinin miktarlarında artıĢ gözlemlenmiĢtir.
E. purpurea L. hücre süspansiyon kültürlerinde, biyotik ve abiyotik stres
faktörlerini kullanarak, kafeik asit ve alkamit üretiminin artırılmasının amaçlandığı baĢka bir çalıĢmada, biyotik ve abiyotik elisitörler olarak, (0, 25, 50 ve 100 mg/l) yeast ekstrakt, (0, 25, 50 ve 100 mg/l) kitosan, (0, 5, 25 ve 50 g/L) sorbitol, (0, 5, 25 ve 50 μM) CdCl2 ve (0, 5, 25 ve 50 μM) AgNO3 kullanılmıĢtır. Hasat edilen hücrelerde
alkamit ve kafeik asit içerikleri belirlenmiĢtir. 50 mg/l yeast ekstrakt uygulamasında, alkamit ve kafeik asit miktarı, kontrole kıyasla sırasıyla 1.3 ve 0.5 kat artmıĢtır. 25 g /L sorbitol ve 100 mg/l kitosan uygulamalarında, kontrole kıyasla alkamit içeriği 0.8 ve 1.5 kat artarken, kafeik asit miktarının 2.5 ve 3.1 kat arttığı belirlenmiĢtir. En yüksek alkamit miktarları CdCl2 ve AgNO3 uygulamalarında (sırasıyla, 105 ve 127 μg/g)