1
TC
Ege Üniversitesi Tıp Fakültesi
Nöroloji Anabilim Dalı
Prof. Dr. Neşe Çelebisoy
SPİNOSEREBELLAR ATAKSİLERDE YUTMA İŞLEVİNİN
ELEKTROFİZYOLOJİK OLARAK DEĞERLENDİRİLMESİ
UZMANLIK TEZİ
Dr. İlay Hilal KILIÇ
İZMİR- 2016
TEZ YÖNETİCİSİ
TEŞEKKÜR
Tez çalışmam sırasında bilgi ve deneyimlerinden yararlandığım; büyük bir fedakarlık
ve sabırla her zaman ve her konuda bana destek olan çok saygıdeğer hocam Prof.Dr.İbrahim
Aydoğdu’ya
Uzmanlık eğitimim süresince katkılarından dolayı tüm saygıdeğer Nöroloji Anabilim
Dalı öğretim üyelerine,
Asistanlık sürecinde aynı yolda omuz omuza yürüdüğüm, bu zorlu süreci güzel anılarla
tamamlamama neden olan değerli asistan arkadaşlarıma,
Tez çalışmam süresince desteğini hissettiğim başta Nörofizyoloji Bilim Dalının
güleryüzlü teknik çalışanları olmak üzere tüm klinik çalışanlarına,
Eğitimimde ve tez çalışmamda katkısı olan tüm hasta ve hasta yakınlarına,
Merak etmeyi, sormayı ve sorgulamayı, özgürce düşünmeyi öğreten; hayatlarını
mutluluğuma adayan, asla hakkını ödeyemeceğim sevgili annem ve babama,
Başarılarıyla, hayattaki duruşlarıyla bana örnek olan ve hiçbir zaman elimi bırakmayan
sevgili ablalarıma,
3
İÇİNDEKİLER
1. GİRİŞ VE AMAÇ ……… 8
2. YUTMANIN ANATOMİSİ VE NÖROFİZYOLOJİSİ ………9
2.1.Yutmada Görevli Anatomik Yapılar ……….9
2.2.Yutma İlişkili Yapıların İnnervasyonu ………..10
2.3.Yutmanın Normal Nörolojik Kontrolü ……….11
2.4.Yutmanın Nörolojik Kontrolünden sorumlu diğer alanlar ………14
3. SEREBELLUMUN YUTMADA GÖREVİ ………15
3.1.İnsanda Serebellum ve Normal Yutma ………..16
4. DİSFAJİ ve SONUÇLARI ………..18
4.1.Penetrasyon ve Aspirasyon ………18
4.2.İnsanda Serebellum Ve Disfaji………..19
5.SOLUNUMUN ANATOMİSİ VE NÖROFİZYOLOJİSİ ……….20
5.1.Solunumla ilişkili santral merkezler………..20
i.Dorsal respiratuar grup ………..21
ii.Pontin respiratuar grup……….21
iii.Ventral respiratuar grup ………...22
6.YUTMA VE SOLUNUMUN KOORDİNASYONU………..23
7.SPİNOSEREBELLAR ATAKSİ ………..27
7.1.SCA’da Klinik Bulgular ………30
7.2.SCA’da Görüntüleme Bulguları ………33
7.3.SCA’da Elektofizyolojik Bulgular ……….34
7.4. SCA’da yutma ilişkili beyin sapı nükleuslarında dejenerasyon ……….34
7.5.SCA’da Disfaji ve Disfajiye Yönelik İncelemeler ………36
8.MATERYAL-YÖNTEM……….37
8.1.Klinik Değerlendirme ………..37
8.1.1.Spinoserebellar Ataksiye Yönelik Klinik Değerlendirme ………..37
i.Ataksi Fonksiyonel Derecelendirmesi ……….38
ii.SCA Hastalarının Tiplendirilmesi ………38
8.1.2.Disfajiye Yönelik Klinik Değerlendirme ……….38
i. Disfajinin Sorgulanması ………38
ii. Disfaji Klinik Bakısı ……….38
8.2.1.Tek Yutma Analizi (Single Bolus Analiz-SBA) ……….40
8.2.2.Disfaji Limiti (DL) ………..41
8.2.3.Bardaktan Su İçme (BSİ) ………..43
8.2.4.BSİ-Solunum Eş Zamanlı Değerlendirme (Sol-BSİ) ……….. 43
8.2.5.Spontan Solunum Kaydı ………..44
8.2.6.Emir-Apne Testi ……….45
8.3.Verilerin Analizi ………..46
9.BULGULAR ………..46
9.1.Demografik/Klinik Bulgular ………46
9.2.Disfaji Limiti (DL) ………...48
9.3.Diğer Elektrofizyolojik Sonuçlar ………..51
9.3.1.Tek Bolus Analizi-Single Bolus Analiz (SBA) ………51
9.3.2.Bardaktan Su İçme (BSİ) ………..57
9.3.3.Solunumla Eş Zamanlı Bardaktan Su İçme (Sol-BSİ) ……….60
9.3.4.Spontan Solunum ……….64
10.TARTIŞMA………....68
ÖZET ……….73
KAYNAKLAR ………76
EKLER ………..89
Ek 1: Spinoserebellar Ataksi Hasta Formu
Ek 2: Ataksi Fonksiyonel Derecesi
5
ŞEKİLLER DİZİNİ
Şekil 2.1: Oral kavite ve farinksin anatomisi
Şekil 2.2: Yutmanın santral patern jeneratörü (CPG)
Şekil 3.1:Sağlıklı insanda yutma esnasında serebellar aktivasyon
Şekil 5.1: Solunumla ilişkili bazı beyin sapı bölgeleri A. Koronal kesit, B. Horizontal kesit. Şekil 5.2: Çeşitli beyin sapı lezyonlarıyla ilişkili anormal solunum paternleri
Şekil 6.1.: Solunum, öksürük, yutma ve kusma CPG’leri arasındaki dinamik etkileşimin şematik
görünümü.
Şekil 9. 1: SCA olgusuna ait patolojik DL örneği
Şekil 9.2: SCA alt gruplarında ve normal kontrollerde ortalama disfaji limiti (DL) değerlerinin
karşılaştırması
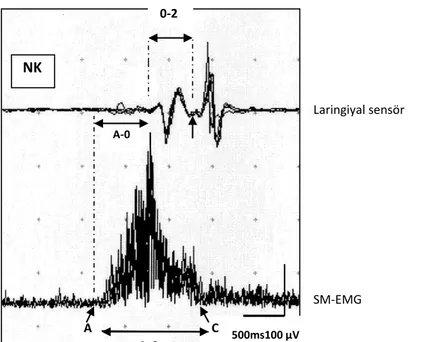
Şekil9.3: NK ve SCA olgusunun SBA traselerinin karşılaştırması
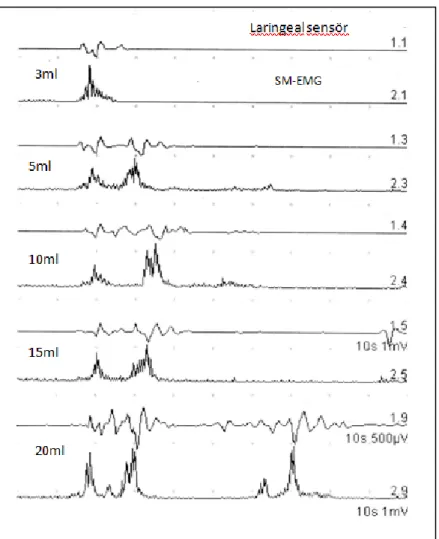
Şekil 9.4: SCA alt gruplarında ve NK’larda SBA parametrelerinin karşılaştırması Şekil 9.5: 3ml tek yutma analizinde SM değişkenliğinin NK ile karşılaştırması Şekil9.6: 3ml tek yutma analizi testinde SM değişkenliği örnekleri
Şekil 9.7: Normal ve patolojik BSİ örneklerinin karşılaştırması
Şekil 9.8: SCA grubu ve NK grubunun 100ml BSİ parametrelerinin karşılaştırılması Şekil 9.9: DL normal, BSİ’si patolojik olan bir olgu örneği.
Şekil 9.10: Normal ve patolojik sol-BSİ örneklerinin karşılaştırması.
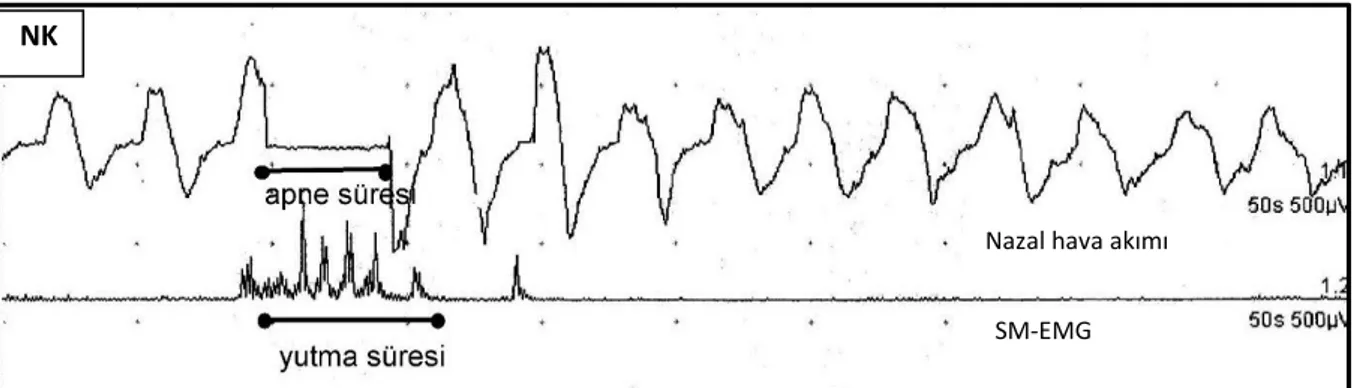
Şekil 9.11: NK ve SCA grubunda SolBSİ sırasında ortalama apne süresi ve apne içi yutma sayılarının
karşılaştırması
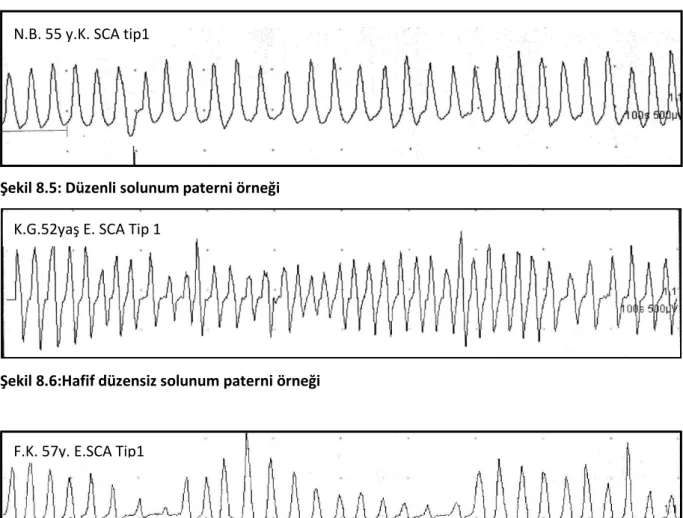
Şekil 9.12:SCA1 tanılı bir olguya ait düzenli solunum paterni örneği. Şekil9.13:SCA3 tanılı bir olguya ait ılımlı düzensiz solunum paterni örneği. Şekil 9.14 :SCA6 tanılı olguya ait şiddetli düzensiz solunum paterni örneği Şekil9.15:SCA1 tanılı bir olguya ait Cheyne-Stokes solunumu örneği Şekil 9.16:SCA1 tanılı olguya ait patolojik emir-apne testi örneği.
TABLOLAR DİZİNİ Tablo2.1:Yutmanın fazları ve ilişkili beyin sapı nükleusları
Tablo 7.1:SCA alt tipleri ve ilişkili tipik klinik, elektrofizyolojik, görüntüleme bulguları
Tablo 7.2:Tipik klinik bulgular ve ilişkili oldukları SCA türleri Tablo 7.3: SCA’da polinöropati
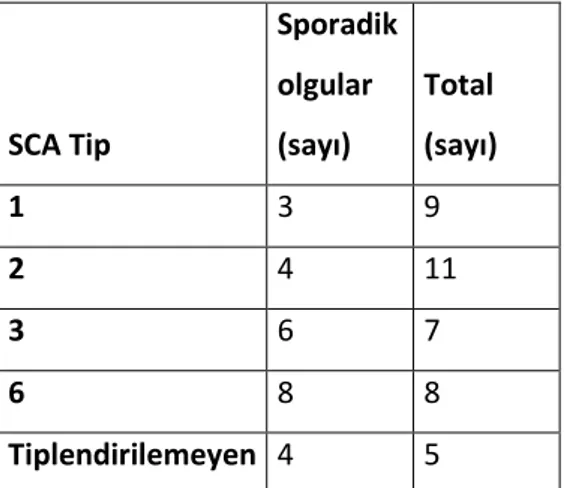
Tablo9.1: Çalışmaya alınan tüm gönüllülerin demografik verileri Tablo 9.2: SCA alt tiplerine göre sporadik olguların sayısı
Tablo 9.3: SCA grubunda saptanan klinik belirti ve bulguların oranları
Tablo 9.4: Sorgulama ve elektrofizyolojik yöntemlerle saptanan disfaji oranları,ortalama hastalık
süreleri ve subklinik disfaji oranları.
Tablo 9.5: 3ml tek yutma analizinde elde edilen veriler
Tablo 9.6: SBA sonuçlarına göre her bir parametre için üst veya alt limiti aşan olguların oranları Tablo9.7:Bardaktan su içme parametrelerinde bireysel patoloji oranları(%).
Tablo 9.8:Solunumla eş zamanlı bardaktan su içme testi sonuçları
Tablo 9.9:Solunumla eş zamanlı bardaktan su içme testi bireysel patoloji oranları
Tablo 9.10: NK ve SCA olgularının 50 sn ve 100 sn spontan solunum ve emir-apne testi sonuçları Tablo 9.11: SCA grubunda Spontan solunum paternine göre olguların oranları (%)
KISALTMALAR DİZİNİ ADCA: Otozomal dominant serebellar ataksi
AFD: Ataksi Fonksiyonel derecelendirmesi AP: Area postrema
BSİ:Bardaktan su içme BötC: Bötzinger complex
cVRG: Kaudal ventral respiratuar grup DD:Disfaji derecesi
DL:Disfaji limiti
DRG: Dorsal respiratuar grup DY:Disfaji yakınması
EEG:Elektroensefalografi EMG:Elektromiyografi
fMRI: Fonksiyonel manyetik rezonans görüntüleme iVRG: Ventral respiratuar grubun inspiratuar parçası MEP:Motor Uyartılmış Potansiyeller
MRG: Manyetik Rezonans Görüntüleme NAC: Nucleus ambiguous caudalis NDV: Nucleus dorsalis nervi vagi NK:Normal kontrol
NOI: Nucleus olivaris inferior NTS: Nucleus tractus solitarii
NV: Nucleus tractus spinalis nervi trigemini NXII: Nucleus nervi hypoglossi
7
OPCA: Olivopontoserebellar atrofi PET:Pozitron emisyon tomografisi preBötC: Pre‐Bötzinger kompleks SBA:Tek yutma analizi
SCA:Spinoserebellar Ataksi
SEP:Somatosensöriyel Uyartılmış Potansiyeller SM-EMG:Submental kas kompleksi yüzeyel EMG’si Sol-BSİ: Solunumla eş zamanlı bardaktan su içme UADRS:Ataxia Disorders Rating Scale
VEP:Görsel Uyartılmış Potansiyeller VFSS:Videofloroskopik yutma çalışması YSİ:Yutma solunum ilişkisi
1.GİRİŞ VE AMAÇ
Yutma, kompleks bir sensorimotor olaydır. Hem istemli hem de istemsiz devinimleri içerir. Belirli bir zaman sırası içinde dudaklar, dil, ağız tabanı, yumuşak damak, farinks, larinks, özefagus ve solunum kaslarının aktivasyonu ile oluşur (Zald ve Pardo 1999; Robbins J ve ark 1999) . İki temel amacı vardır: Oral kavitedeki yiyeceği mideye iletmek ve hava yolunu korumak. Yutma işlemi genel olarak bolusun bulunduğu bölüme göre oral, faringeal ve özofagial evre olarak ayrılmaktadır (Hamdy S ve ark 1999a).
Disfaji, terminolojik olarak, gıdaların ağızdan mideye hareket etme güçlüğüdür. Serebral korteksten yutmayla ilişkili kaslara kadar nöromüsküler yolağın herhangi bir yerindeki lezyona bağlı olarak oluşabilmektedir (Zald ve Pardo1999; Robbins J ve ark 1999). Nörolojik hastalıklar, özellikle orofaringeal disfaji olmak üzere, yutma problemi olan hastaların yaklaşık %70-80’inden sorumludur (Hamdy ve ark 1999b; Kern ve ark 2001a; Kern ve ark 2001b). Nörojenik disfaji bolus iletiminde aksamaya ve bolusun nazal kavite, larinks ve/veya trakea gibi yanlış yerlere kaçmasına neden olabilmektedir (Martin ve ark 2001; Martin ve ark 2004). Nörojenik disfajinin ana sonuçları aspirasyon ve buna bağlı gelişen pnömoni, malnütrisyon ve dehidratasyondur. Bunlar; inme, demans ve diğer ilerleyici nörolojik hastalıklarda ölümcül olabilecek komplikasyonlardır. Öte yandan kronik disfajinin maddi götürüsü de oldukça fazladır. Bu açıdan yüksek riskli hastaların disfaji yönünden araştırılması ve erken teşhisi önemlidir (Mosier ve ark 1999). Etkilenmiş bireylerde nörojenik disfajinin erken teşhisi yaşam kalitesi ve sağ kalım açısından da önemlidir. Nörojenik disfaji; amiyotrofik lateral skleroz (ALS), Parkinson hastalığı , demans, multiple skleroz ve bazı nöromüsküler hastalıklarda olduğu gibi sinsi başlangıçlı ve yavaş seyirli olabilmektedir. Bu kronik nörolojik hastalıklarda belirgin klinik disfaji ortaya çıkmadan önce subklinik bir dönem olabilmektedir. Disfajiyi erken, subklinik dönemde yakalamak, komplikasyonların önlenmesi açısından önemlidir. Bu durumun erken tespiti , rehabilitatif , tıbbi ve cerrahi tedavilere bir an önce başlamaya olanak sağlar (Mosier ve ark 1999b;Mosier ve ark 2001,Suzuki M ve ark 2003,Toogood ve ark 2005; Bear ve ark 2001; Ertekin ve ark 2003; Satow ve ark 2004; Jean ve ark 1997; Daniels ve ark 1997; Houk ve ark 2007) .
Sinir sisteminin çeşitli bölgelerinin, yutma işlevinde görevli olduğu öne sürülmüştür. Bunlar; presantral girus (MI), postsantral girus (SI), premotor alan (PMA), suplementar motor alan (SMA), anterior singulat korteks (ACC), operkulum, insula, prekuneus, kuneus, prefrontal alan, temporal korteks, serebellum, beyin sapı, frontal korteks, internal kapsül, asosiasyon alanları, talamus ve bazal ganglionlardır (Mussen ve ark 1930; Bernston ve ark 1973; Martner ve ark 1975; Weerasuriya ve ark 1979; Gibbs ve ark 1992; Bostan ve ark 2010; Grillner ve ark 2013; Harris ve ark 2005; Malandraki ve ark 2009; Grabski ve ark 2012). Serebellumun disfajideki ve disfaji ilişkili nörolojik bozukluklardaki rolü giderek daha fazla anlaşılmaktadır. Ancak pür serebellar lezyonlardan elde edilen az sayıdaki verilerle, bu rolün doğası halen iyi aydınlatılamamıştır (Shibamoto ve ark 2007). Serebellar
9 nörodejeneratif hastalıklarda disfajinin sıklığı ve doğasına yönelik klinik ve elektrofizyolojik araştırmalara ihtiyaç vardır. Ön planda serebellum ve bağlantılarının etkilendiği; genetik, nörodejeneratif bir hastalık olan Spinoserebellar ataksi (SCA)’li bireylerin elektrofizyolojik yöntemlerle incelenmesinin , hem bu hastalık grubundaki disfajinin doğası hakkında fikir vermesi, klinik ve genetik verilerin ışığında disfaji yönünden riskli grupların belirlenmesi , SCA’nın ana mortalite ve morbidite nedeni olan disfajinin erken tespiti, prognozun öngörülmesi ve gerekli müdahalelerin yapılmasına ; hem de yutma fizyolojisinde serebellum ve bağlantılarının fonksiyonları hakkında ipuçları elde etmeye olanak sağlayacağını düşünüyoruz. Bu amaçla spinoserebellar ataksi tanısı almış 33 hasta ve bu hastalara yapılan toplam 40 yutma EMG testi incelenmiş; veriler olguların klinik , disfajiye yönelik sorgulama ve muayene sonuçları eşliğinde gruplandırılmıştır. Grupların verileri hem kendi içinde hem de sağlıklı 26 gönüllüye yapılan yutma EMG testi sonuçlarıyla karşılaştırılarak analiz edilmiştir.
Bu çalışmada kullanılan yutma EMG testi, Ege Üniversitesi Tıp Fakültesi Hastanesi, Nöroloji kliniği, Klinik Nörofizyoloji Bilim Dalı’nda Ertekin ve ark. tarafından geliştirilmiş olup, daha önce çeşitli araştırmalarda kullanılmıştır (Ertekin ve ark 1995, 1996; Ertekin &Aydoğdu 2003, Aydoğdu ve ark 2011, Aydoğdu ve ark 2015).
2.YUTMANIN ANATOMİSİ VE NÖROFİZYOLOJİSİ 2.1.Yutmada Görevli Anatomik Yapılar
Yutma 30’dan fazla sinir ve kasın devreye girdiği hem istemli hem de refleks aktivitelerden oluşan bir davranıştır. Anatomik olarak oral kavite, fariks, larinks, özofagus ve ilgili yapılar görev almaktadır.
Oral kavite: Dudaklar, dişler, dil, sert damak, yumuşak damak, uvula, palatal arklar, tonsiller
plikalar, mandibula ve ağız tabanından oluşmaktadır. Burada ağza alınan gıdalar bolusa çevirilir.Bu yapılardan bazıları 2 fonksiyonel sfinkteri oluştururlar: Yumuşak damak ve dil tarafından oluşturulan ‘glossopalatal sfinkter’ ve yumuşak damak ve üst faringiyal konstriktör kaslar tarafından oluşturulan ‘palatofaringiyal sfinkter’. Palatofaringiyal sfinkter bolusun nazofarinkse kaçmasını önler (Cook 1991) . Ağız tabanında bulunan submental-suprahyoid kas kompleksi, orofaringiyal yutma sırasında ilk aktive olan kas grubudur. Mylohyoid, geniohyoid, digastrik ve stilohyoid kaslardan oluşur. Bu kaslar, hem hyoid kemiği kafa tabanına ve mandibulaya bağlar; hem de dil hareketlerine destek olur. Bizim çalışmamızda da bu kas grubundan EMG kayıtlaması yapılmıştır.
Farinks: Oral kaviteden iletilen bolus, orofarinkse gelir. Buradan da üst, orta ve alt faringeal
konstriktör kaslarla özofagusa iletilir. Stilofaringiyal, salpingofaringiyal ve palatofaringiyal asıcı kaslar yutma sırasında farinksin kısalması ve genişlemesi ile larinksin elevasyonuna yardımcı olurlar. Faringoözofagiyal bileşkede, başlıca krikofaringiyal kas (CP) tarafından oluşturulan, C şeklinde sirküler çizgili kas liflerinden oluşan ‘üst özofagiyal sfinkter’ yer alır. Bu yapı önde krikoid kıkırdağa
tutunur. Yutma dışında devamlı tonik aktivite göstererek özofagus içeriğinin geriye kaçışını ve solunum sırasında havanın özofagusa geçişini önler. Farinkste yutma öncesinde ve sonrasında materyal birikiminin olabildiği 2 sinüs bulunur. Bunlardan ‘sinus piriformis’ inferior konstriktor kasın tiroid kıkırdak lateral duvarına tutunduğu bölgede , ‘vallekula’ ise dil tabanı ile epiglottis arasında yer alır.
Larinks: Larinksin intrinsik kasları, epiglottis, ariepiglottik katlantılar, yalancı ve gerçek vokal
kordlar sayesinde yutma esnasında solunum yolu korunur.
Özofagus: Peristaltik hareketlerle bolusu mideye taşıyan tüp şeklinde bir yapıdır. Üst 1/3’lük
kısmı çizgili kaslardan, alt 1/3’lük kısmı düz kaslardan ve orta 1/3’lük kısmı hem çizgili hem de düz kaslardan oluşur. Üst özofagial sfinkter gibi, yutma haricinde tonik olarak kasılı duran ‘alt özofagial sfinkter’ yutma sırasında gevşeyerek, bolusun mideye geçişine izin verir.
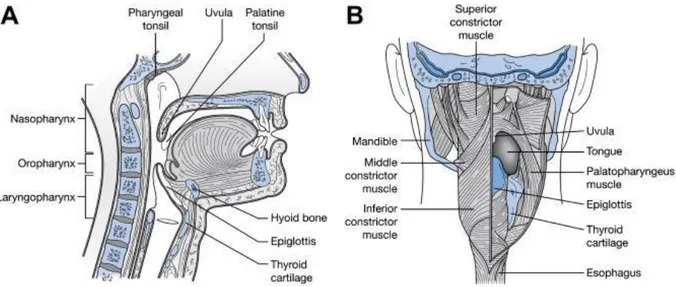
Oral kavite ve farinksin anatomisi şekil 2.1’de gösterilmiştir.
2.2.Yutma İlişkili Yapıların İnnervasyonu
Motor İnnervasyon: Dilin ekstrinsik kasları ansa servikalis, intirinsik kasları hipoglossal sinir
(XII) , stilofaringiyal kas hariç tüm farinks kasları, tensor veli palatini hariç tüm yumuşak damak kasları, krikofaringiyal kas ve larinks intrinsik kasları vagal sinir (X) ile innerve edilir. Çiğneme kasları mandibular sinir (V3), suprahyoid kaslar mandibular sinir (V3), fasiyal sinir (VII) ve ansa servikalis tarafından innerve edilir.
Duysal İnnervasyon: Oral kavitenin duyusu fasiyal sinir (VII) ve trigeminal sinir (V2)’le ; farinks
ve tonsiller plikaların duyusu glossofaringiyal sinir (IX)’le ; dil kökü, hipofarinks ve posterior larinksin duyusu superior laringiyal sinir (X)’le taşınır. Superior laringiyal sinir, yutmanın tetiklenmesinde önemlidir.
Yutma işlemi için periferden gelen duysal girdiler ve dudak, dil, çiğneme kasları, ağız tabanı, yumuşak damak, farinks, larinks ve özofugusun aktive olması gerekmektedir. Yutma, anatomik olarak birbirinden bağımsız ancak fonksiyonel ve zamansal olarak birbiriyle devamlılık gösteren; oral (hazırlık), faringeal ve özofagial evre olmak üzere 3 evrede gerçekleşmektedir (Magendie 1836). Orofaringiyal dönem fonksiyonel olarak bütünlük gösterir ve tek bir yutma için tanımlanmış total süre 0,6-1sn’dir. Özofagiyal dönem total süresi ise bilinçli insanda >10 sn tanımlanmıştır.
Normal yutma şu şekilde gerçekleşir: Öncelikle ağıza alınan bolus dilin üstünde ya da önünde durur.Dil ucu (oral dil), bolusu kaldırıp damağa doğru yükselterek, bolusun istemli olarak arkaya iletilmesini başlatır. Bolus ilerleyişi sırasında reseptörlerle temas ederek superior laringiyal ve glossofaringiyal sinirin afferent liflerini uyarır. Bu uyarı beyin sapında (medulla) nukleus traktus solitariusa ulaşır. Böylece yutmanın otomatik elementleri ortaya çıkmaya başlar. Pattern oluşturmuş outputlar, bir seri ardışık nöromusküler olayı başlatır. Yumuşak damak yükselerek nazofarinksi kapatır. Farinks kasılarak öne ve mediale yer değiştirir. Dil kökü (faringiyal dil) farinkse doğru
11 yaklaşarak bolusu sıkıştırır. Hyoid ve larinks yukarı ve öne doğru yer değiştirerek bolusun önünden çekilir. Epiglottis öne doğru eğilerek larinksi kapatır. Üst özofagiyal sfinkter gevşeyerek bolusun özofagusa geçişine izin verir (Gates ve ark 2006).
Şekil 2.1: Oral kavite ve Farinksin anatomisi (Moses ve ark. 2005)
Oral kavite ve farinks anatomisinin lateral (A) ve posterior (B)’den görünüşü gösterilmektedir (Moses KP 2005)
2.3.Yutmanın Normal Nörolojik Kontrolü a)Kortikal Düzey:
Yutma ihtiyacı farinksteki sensöriel girdilerin spinal kolumnadan beyin sapına; oradan da talamusa ulaşması; ve talamustan pariyetal korteksin ilgili spesifik duysal alanlarına projeksiyonu ile başlar (Bear ve ark 2001). Kortikal aktivasyon ve yutmayla ilgili araştırmaların çoğunda postsantral girusun hem istemli hem de otomatik yutmada aktive olduğu bildirilmektedir.(Zald ve ark 1999;Hamdy ve ark1999a,b Kern ve ark 2001a ;Martin ve ark 2001;Martin ve ark 2004;Mosier ve ark 1999a; Mosier ve ark 1999b; Mosier ve ark 2001;Suzuki ve ark 2003; Toogood ve ark 2005). Sensöriel parçada ; yüz, dil, faringeal, laringeal ve özofagial alanlar somatotopik olarak yerleşmiştir (Hamdy ve ark1999b, Mosier ve ark 1999b). Bu alan; dil, çene, palatal ve faringeal kasların hareketiyle oluşan orofaringeal stimülasyondan sorumludur ( Martin ve ark 2001). Çeşitli çalışmalar; nükleus traktus solitarius (NTS)’a pek çok direkt projeksiyonu olan (Mosier ve ark 1999b ), girus postsantralisin lateral parçasının uyarılmasının yutmayı tetikleyebildiğini göstermiştir (Hamdy ve ark1999b). Bazı çalışmalar postsantral girusun, hem insula hem de presantral girusla bağlantıları sayesinde, yutmanın motor kontrolünü de düzenlediğini belirtmektedir (Hamdy ve ark1999a; Ertekin ve Aydogdu 2003). Bu bağlantılar kas kontraksiyonunun şiddetinin algılanmasını ve gerekirse değiştirilmesine olanak sağlamaktadır (Martin ve ark 2004). Bu, hem istemli hem de otomatik yutmanın düzenlenmesinde duysal geri bildirimlerin önemini ortaya koymaktadır (Martin ve ark 2001).
Ayrıca; kortikal aktivasyon ve yutmayla ilgili araştırmaların büyük bölümü anterior singulat korteks, suplementar motor alan ve premotor alanların istemli yutma sırasında aktive olduğunu göstermiştir (Zald ve Pardo 1999; Hamdy ve ark 1999a,b; Kern ve ark 2001;Martin ve ark 2004; Mosier ve ark 1999a,b; Mosier ve ark 2001;Suzuki ve ark 2003).
Hem yutma hem de dil hareketleri, suplamenter motor alan ve postsantral girus arasında paralel olarak işlenmektedir (Satow ve ark 2004). Anterior singulat korteks, yutmaya ilişkin önemli bir alandır ve yutmanın erken döneminde aktive olduğu görülmüştür (Hamdy ve ark 1999a,b ;Ertekin ve ark 2003). Yutma haritalaması çalışmaların büyük çoğunluğu anterior singulat korteksin istemli yutmada aktive elduğunu belirtmektedir (Hamdy ve ark 1999 a,b; Kern ve ark 2001 a,b; Martin ve ark 2001; Martin ve ark 2004;Mosier ve ark 1999 a,b; Mosier ve ark 2001;Suzuki ve ark 2003; Toogood ve ark2005). Anterior singulat korteks, sensorimotor bir planın seçilmesinde (Mosier ve ark 2001) olduğu kadar hareket başlangıç modunun, hareketin tipinin ve sıklığının kodlanmasında da önemli rol oynamaktadır (Martin ve ark 2004).Yutmanın premotor aşaması için aktive olmuş ve bazal ganglionlardan inputlar almış olan suplamenter motor alan; bunun sonucunda presantral girusu uyarır (Bear ve ark 2001). Suplamenter motor alan ve presantral girus arasında önemli bir bağlantı vardır. Ayrıca ikisi de postsantral girustan, posterior paryetal korteksten, talamustan ve serebellumdan inputlar almaktadır (Rangarathnam ve ark 2014).
b)Beyinsapı Düzeyi:
Yutma için temel nöronal ağ medulla oblangatadaki yutma merkezi ve çevresinde bulunmaktadır. Bu alan, bilateral yerleşmiş dorsal meduller bölgedeki nukleus traktus solitarius (NTS) ve çevresindeki ventromeduller retikuler formasyon (VMRF) ile ventral bölgedeki nukleus ambigiuus’tan (NA) oluşmaktadır. Periferik yutma alanlarından kalkan duysal afferentler (V,VII,IX ve X’uncu kraniyal sinirler vasıtasıyla) ve kortikal inici yollar NTS’de kaudorostral olarak sonlanmaktadır ( Jean ve ark. 2001). NTS ve VMRF’ye gelen afferent girdiler burada süzülür. Uygun bulunan afferent impulslar internöronal havuza geçer. Çift yönlü olarak afferent ve efferent düzeylerle iletişim içinde olan, yutmanın motor evresini başlatabilen ve organize edebilen bu internöronal havuz yutmanın ‘Santral Patern Jeneratör’üdür (CPG) (Ertekin ve Aydogdu 2003). Burada işlenen impulslar NA’ya projekte olan efferentleri oluşturur. NA’nın aksonları da yutma ile ilişkili değişik düzeylerdeki motor nükleuslarda (IX, X ve XI.) sonlanım gösterir.Bulbusta yer alan yutmanın CPG’ü 3 bölgeden oluşmaktadır:
1-Dorsal Yutma Grubu (DSG): NTS ve etrafındaki internöronlardan oluşan bölgedir. Yutmanın
CPG’nün afferent işleyişten sorumlu olan bu alanı kortikal-subkortikal yollardan gelen inici sinyalleri ve periferden gelen duysal girdileri alır. ‘Jeneratör Nöronlar’ olarak da adlandırılan bu bölge; yutmanın tetiklenmesi, ardışık ve ritmik yutma paterninin organizasyonundan sorumludur (Jean 1972; Jean 1978).
13
2-Ventral Yutma Grubu (VSG) : Venrolateral medullada, NA’nın üzerinde yerleşmiştir .
Yutmanın CPG’nün efferent işleyişten sorumlu bu alanı; NA’tan motor çıktıları alıp ilgili kraniyal motor nukleusların motor nöron havuzuna dağıtır. ‘Switching Nöronlar’ olarak da adlandırılan bu bölge; DSG tarafından salınan nöral paterni birden çok motor nöron havuzuna sıralı olarak dağıtır (Jean 1983).
3-Motor Nöron Havuzundaki İnternöronal Ağ: Ventrolateral medullada yer alan ve yutmanın
CPG’de önemli rol alan VSG’nun dışında V. ve XII. kranial sinirleri ve etrafını içeren bir bölgede yutmanın internöronları tespit edilmiştir. Afferent ve efferent düzey arasında bulunan bu nöronların premotor nöron görevi gördüğü ve yutmanın longitudinal ve bilateral organizasyonundan sorumlu olduğu düşünülmektedir (Jean 2001).
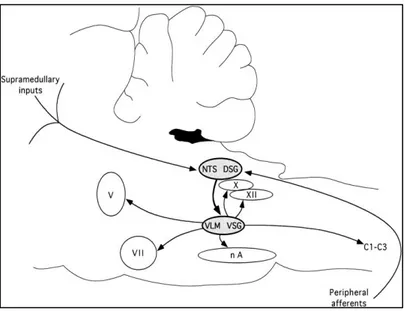
Pontin İnternöronlar ise ponsun rostral parçasında, V. Kranial sinir motor nükleusunun üstünde, prinsipal duysal trigeminal nukleus düzeyinde yerleşiktir. Bu pontin nöronların orofaringeal reseptörlerden gelen verileri santral sinir sisteminin daha üst merkezlerine aktarmakla görevli olduğu ve yutmanın CPG’üne dahil olmadığı düşünülmektedir (Jean 2001). Yutmanın organizasyonunda görevli beyin sapı bölgeleri şekil2.2 de , yutmanın fazları ve ilişkili beyin sapı bölgeleri tablo 2.2’de gösterildi.
Şekil 2.2: Yutmanın santral patern jeneratörü (CPG)
CPG, medulla oblangatada 2 ana nöron grubundan oluşmaktadır: Dorsal yutma grubu (DSG), NTS ve etrafındaki internöronlarda lokalizedir, venral yutma grubu (VSG) nukleus ambiguus (NA)’a komşu ventrolateral medullada lokalizedir. DSG, ardışık ya da ritmik yutma paterninin tetiklenmesi, şekillendirilmesi ve zamanlamasında görevli jeneratör nöronları içerir. VSG ise yutma güdüsünü yutma ilişkili çeşitli motor nöron havuzlarına dağıtan dönüştürücü (switching) nöronları içerir. Yutma ve elementer reflekslerden sorumlu bu yolak; periferik afferent lifler, dorsal ve ventral yutma grubu nöronları ve motor nöronlardan oluşmuş oligosinaptik bir devre (loop) ‘dir (Jean A.2001).
Tablo2.1:Yutmanın fazları ve ilişkili beyin sapı nükleusları
Yutmanın fazı Fonksiyonu İlişkili beyin sapı nukleusu
Hazırlık fazı Oral duysal değerlendirme, çiğneme, lingual arama ve lingual transfer işlevleri
Mezensefalik, prinsipal, spinal ve motor trigeminal nükleuslar;fasial nükleus; hipoglossal nükleus
Lingual faz Yiyecek ve içeceklerin orofarinkse iletilmesi
Mezensefalik, prinsipal, spinal ve motor trigeminal nükleuslar;fasial nükleus;nükleus ambiguus;hipoglossal nükleus; pigmente ve parvoselüler soliter nükleus; parvoselüler retiküler nükleus ve intermediat retiküler zon Faringiyal faz Havayolunu koruyarak
yiyecek ve içeceklerin farinksten geçirilmesi
Fasial nükleus; nükleus ambiguus;hipoglossal
nükleus;medial,pigmente ve komissural soliter nükleuslar; parvoselüler retiküler nükleus ve intermediat retiküler zon Özofagiyal faz Yiyecek ve içeceklerin
mideye peristaltik transportu
Nükleus ambiguus; nükleus dorsalis nervi vagi
2006 Blackwell Publishing Ltd,Neuropathology and Applied Neurobiology,32, 635–649
2.4.Yutmanın Nörolojik Kontrolünden sorumlu diğer alanlar:
İnsula ve operkuler korteksin yutmadaki rolünü aydınlatmaya yönelik çalışmalar devam etmektedir. İnsula, frontal, temporal ve paryetal operkulumun birleştiği bölgenin hemen altında, derinde, lateral sulkusta yer alır (Daniels ve ark 1997). Nörogörüntüleme çalışmalarının çoğu, insular korteksin hem istemli hem de otomatik yutmada görevli olduğunu belirtmektedir. Yine pek çok çalışma, insular kortekste sağ lateralizasyon olduğunu ileri sürmektedir (Zald ve ark 1999; Hamdy ve ark 1999 a,b; Kern ve ark 2001 a,b; Martin ve ark 2001; Mosier ve ark 1999 a,b; Suzuki ve ark 2003;Toogood ve ark 2005). İnsulanın postsantral girus, primer motor alan, suplamenter motor alan ,bazal ganglionlar, talamus, singulat, NTS, operkulum ve tat,koku,limbik, otonomik alanları içeren pek çok kortikal ve subkortikal yapıyla bağlantılı olduğu bildirilmiştir (Martin ve ark 2001;Mosier ve ark 2001;Daniels ve Foundas 1997). İnsula, disfaji yakınması olan unilateral inme hastalarında en sık rastlanan lezyon alanıdır (Daniels ve Foundas 1997). Operkuler korteks, insulanın üzerini örter ve yutma işleminde görevli olduğu bildirilmiştir. Operkuler korteks, Martin ve ark(2004)’ın uyarıldığında tam bir yutma oluşturabileceğini söylediği 4 kortikal alandan biridir. Operkulum; çiğneme ve diğer
15 yutma ilişkili oral hareketlerin zamanlamasını kontrol ve idare etmek gibi, yutmanın zamansal organizasyonunun yapılmasından sorumludur (Martin ve ark 2004) Aynı zamanda burası ağız ve farinks bölgesinin duyusunu alması nedeniyle de önemlidir (Martin ve ark 2001; Ertekin ve Aydogdu 2003).
3.SEREBELLUMUN YUTMADA GÖREVİ
Serebellum, motor çıktıları denetler ve kişinin yapmak istediğiyle yaptığı hareketi karşılaştırarak gerekli düzeltmeleri yapar. Ponsta bulunan pontin nukleuslar; suplamenter motor alan, premotor alan, presantral girus, postsantral girus ve posterior paryetal korteksin aksonlarından projeksiyonlar alıp bu bilgileri serebelluma taşır. Serebellum, bu bilgileri hareketin yönü, zamanlaması ve gücü açısından işleyerek talamus aracılığıyla motor kortekse geri gönderir (Bear ve ark 2001). Serebellumun motor koordinasyon, zamanlama, sıralama ve proprioseptif geri bildirimden sorumlu olduğu düşünülürse (Houk ve ark 2007) yutma ilişkili fonksiyonlarda da destekleyici rolünün olması şaşırtıcı değildir. Ancak litratürde nörojenik yutma bozukluğuyla ilgili çelişkili veriler bulunmaktadır. İnsan dışı primatlarla ilgili birkaç çalışma, yutma ilişkili motor hareketleri daha iyi kavramamızı sağlamıştır. Mussen (1930), serebellumun yutmayla ilişkili olduğunu ortaya koyan ilk araştırıcıdır. Vermisin ventral bölümlerinin uyarılmasıyla farinks ve oral kavite kaslarının kasıldığını belirtmiştir. Bernston ve ark (1973), bundan çok sonra kedilerde , fastigial nukleustan süperior serebellar pendinküle kadar uzanan bölgelerin uyarılmasıyla iyi organize olmuş yeme ve tımarlanma davranışı olduğunu bildirmiştir. Martner(1975), daha spesifik olarak fastigial nukleusun uyarılmasıyla kedilerde tımarlanma ve çiğneme için önemli oral davanışlar gözlemlemiş ve serebellumun kedilerde çiğneme hareketlerinde rolü olduğunu ileri sürmüştür. Daha sonraları, fastigial nukleustaki nöronların beslenme davranışıyla ilgili diğer bölgelerle de irtibat halinde olduğunu belirtmiştir. Kedilerde serebellumun elektriksel uyarımının yutma ilişkili kortikal çıktılar üzerinde inhibitör etkisinin gösterilmesi de serebellumun bu bölgeler arasında koordinasyon sağladığı görüşünü desteklemektedir (Weerasuriya ve ark 1979). Gibbs (1992) ise serebellumdaki nukleus interpositus ve süperior serebellar pedinkül lezyonu olan tavşanlarda çene hareketleri ve göz kırpma yanıtları üzerinde çalışmış ve destekleyici veriler elde edememiştir. Bazı yeni çalışmalar, insan dışı primatlarda yutma ilişkili sensöri motor hareketlerde serebellumun olası rolü hakkında dolaylı olarak fikir sağlamıştır. Örneğin, Bostan ve ark. (2010 a,b) cebus maymunlarında bazal ganglionlarla serebellum arasında disinaptik bağlantılar göstermiş ve bu ikisi arasında iletişim sağlayan yolaklar olduğunu ileri sürmüştür. Bazal ganglionların, olasılıkla serebellumun da katkısıyla, primatlarda yutma ilişkili ince motor hareketlerde önemli rolü olduğuna dair yayınlar giderek artmaktadır (örn., Grillner ve ark 2013). Serebellumun serebral korteksle bağlantısının da önem arzettiği düşünülmektedir. Serebellar output sonucunda serebral kortekste geniş bir alanda aktivasyon olduğu bildirmiş ve serebellumun yutma gibi daha kompleks işlevlerde görevli olduğu belirtilmiştir (Bostan ve ark 2013) .
İnsanlarda serebellar fonksiyonlara yönelik çalışmalar, serebral korteks ve serebellum arasında monosinaptik bağlantılar olmamasından dolayı geleneksel anterograd ve retrograd takip yöntemlerinin etkin bir şekilde kullanılamaması nedeniyle, oldukça sınırlıdır. Bu yüzden serebellumun normal yutmadaki rolü hakkındaki bilgilerimiz nörogörüntüleme verileri ve çeşitli lezyonlara bağlı anormal pattern örneklerine dayanmaktadır (Rangarathnam ve ark 2014).
3.1.İnsanda Serebellum ve Normal Yutma
Son 15 yılda araştırıcılar, yutma sırasında nöronal aktivasyon patternlerini incelemek için fMRI ve PET gibi görüntüleme tekniklerini kullanmışlardır. Prospektif nörogörüntüleme çalışmalarından birinde Zald ve Pardo (1999) sağlıklı gönüllülerde sekresyonların yutulması esnasında bölgesel serebral kan akımı (rCBF)’nı Pozitron Emisyon Tomografisi (PET)’ni kullanarak incelemişlerdir. Sol inferior presantral girus, insula ve sol serebellumda (özellikle crus Ia/VI bölgesinde) belirgin bir aktivasyon gözlenmiştir. Araştırmanın devamında bu aktivite, başka sağlıklı bir grupta dil hareketleriyle ortaya çıkan aktiviteyle kıyaslanmış ve birbirinden çok farklı olduğu görülmüştür. Bu yüzden serebellumun, özellikle VI. lobülünün faringeal yutmada önemli olduğu belirtilmiştir.
2001’de Mosier ve Bereznaya , yutmanın hiyerarşik olarak düzenlenmiş görevler şeklinde değil, paralel ağlar(network)ın aktivasyonuyla gerçekleştiğini öne sürmüştür. Yutma sırasında fonksiyonel manyetik rezonans görüntüleme (fMRI)’yle elde edilen aktivasyonların strüktürel eşitlem modelleme (SEM) yöntemiyle yolak (pathway) analizleri yapılmış ve serebellumun istemli yutma sırasında primer motor korteks, inferior frontal girus, bazal ganglionlar ve talamusla fonksiyonel (anatomik değil) bağlantılar sergilediği tespit edilmiştir. Aktive olan yapılar, bağımsız birimler halinde çalışmaktadır ve her birimin yutma sırasında ayrı bir görevi vardır. Serebellum, bu birimlerden birini oluşturur ve olasılıkla oral-lingual ve faringeal kaslar için ileri besleme (feed-forward) mekanizmaları ve zamanlama, sıralama, internal koordinasyonun kontrolününde rol oynamaktadır (Mosier ve Bereznaya 2001). 1999’da Zald ve Pardo’nun çalışmaları da bunu desteklemektedir.
Suzuki ve ark (2003) 11 sağlıklı gönüllüde istemli sekresyon yutma esnasında fMRI tekniğiyle yaptığı incelemede özellikle arka bölümlerinde belirgin olmak üzere her iki serebellar hemisferde aktivasyon göstermiştir (Şekil 3.1) . 2005’te Harris ve ark istemli yutma sırasında rCBF’nin değerlendirildiği bir PET çalışmasında sol serebellar hemisferde aktivasyon gözlemlemişlerdir. Presantral ve postsantral giruslar, insular girus, talamus, globus pallidus, anterior singulat girus, supplamenter motor alan, superior temporal girus ve substantia nigra’da güçlü bir aktivasyon saptanmıştır. 2009’da Malandraki ve ark yutmanın belli aşamaları sırasındaki beyin aktivasyonunu araştırmışlardır. Yutma gerçekleşmeden yutmanın planlanması, dilin damağa değdirilmesi, boğaz temizleme hareketi ve 3ml sıvı yutma komutları verilmiştir. Bu esnadaki nöral aktivasyon fMRI ile incelenmiştir. 3ml yutma işlemi serebellumun anterior ve posterior bölgelerinde çeşitli alanlarla beraber bazı kortikal ve subkortikal yapılarda da aktivasyona neden olmuştur. Larinksin kapanmasını
17 sağladığı düşünülen boğaz temizleme hareketi, bilateral serebellumun posterior bölümlerinde aktivasyon sağlamıştır. Dilin damağa değdirilmesi ve yutmanın planlanması esnasında serebellar aktivasyon gözlenmemiştir. Bu bulgular, serebellumun yutmanın belli aşamalarında görevli olması bakımından önemlidir. Grabski ve ark (2012) da serebellumun supralaringeal ve laringeal motor kontroldaki rolünü araştırmıştır. Yaptıkları fMRI çalışmasında, artikülasyon hareketleri ve sesli harf (‘i’) çıkarma sırasında serebellar hemisferlerin süperior parçasında aktivasyon saptanmıştır. Araştırıcılar hem konuşma sırasındaki artikülatör hareketlerle; hem de çiğneme, yutkunma gibi konuşma dışı hareketlerle benzer sonuçlar elde etmiş ve serebellumun yutma esnasında oral motor yapıların koordinasyonundan sorumlu olabileceğini öne sürmüştür. Shibamoto ve ark (2007) sadece kapsül yutma sırasında serebellumda aktivasyon gözlemlemiş, diğer bolus içeriklerinde ise öyle bir şey saptamamışlardır. Bu sonuçlar, yutma mekanizmasında farklı içeriklerin farklı derecelerde motor koordinasyon gerektirdiği şeklinde yorumlanmıştır. Jayasekeran ve ark. (2011) bir çalışmada serebellumun transkranial manyetik stimülasyonunun (TMS) faringeal konstriktör kaslar üzerine etkisini araştırmıştır. Bu çalışmada 16 sağlıklı gönüllünün serebellar hemisferleri ve orta hatta vermisi uyarılmıştır. Sonuçta kortikal stimülasyona kıyasla daha az olmakla birlikte, serebellar stimulasyondan sonra da faringeal bölgelerde belirgin motor uyarılmış potansiyeller (MEP) elde edilmiştir. Ayrıca kortikal uyarım öncesinde serebellar uyarım yapıldığında, kortikal stimulasyon sonrası motor yanıtlarda belirgin fasilitasyon gözlenmiştir. Bu, serebellumun, yutma esnasında kortikal ateşlemeyi yoğunlaştırdığı ya da modüle ettiğini düşündürmektedir. Bu çalışma, kortikal stimulasyona ek olarak serebellar stimulasyonun da uygulandığı TMS’nin rolünü anlamamıza katkı sağlamaktadır. Mottolese ve ark (2013), insan serebellumunda motor temsil alanlarını haritalandırmaya çalışmıştır. Bu çalışmada serebellum dışındaki bir tümör operasyonu esnasında, serebellumun posterior bölgelerine elektriksel uyarı verilmiş ve lobül VI’ün yüz ve ağız bölgesini temsil ettiği görülmüştür. Bu da yine lobül VI’ün yutmayla ilişkili olabileceğini düşündürmektedir.
Şekil 3.1:Sağlıklı insanda yutma esnasında serebellar aktivasyon (Suzuki ve ark 2003)
4.DİSFAJİ ve SONUÇLARI
Disfaji, yani anormal yutma çok çeşitli nedenlerle ortaya çıkabilmektedir (Kuhlemeier1994; Castell 1987). Oral kavite, farinks, larinks, özofagus ya da özofagial sfinkterin işlevsel ya da yapısal bozuklukları disfajiye yol açabilir. Disfaji; dehidratasyon, malnütrisyon, pnömoni ya da hava yolu obstrüksiyonu gibi ciddi komplikasyonlara yol açabilir. Üzerinde duracağımız orofaringeal disfajinin başlıca bulgu ve belirtileri şöyledir: Beslenme sırasında veya sonrasında ıslak ses; beslenmeden önce, esnasında ya da sonrasında öksürük, boğulacak gibi olma; oral sekresyonlarda artış; yutmayı başlatmada zorluk; gıdanın boğazda takılma hissi; drooling; kilo kaybı; kronik alt hava yolu sekresyonları; yavaş yeme gibi beslenme davranışlarında değişiklik; pnömoni; seste kabalaşma; dispne; nazal regürjitasyon (Logemann 1996).
4.1.Penetrasyon ve Aspirasyon
Yutma sırasında havayolunun korunması elzemdir ve yetersizliğinde ciddi sonuçlar doğurabilir. Ağıza alınan ya da özofagustan regürjite olan materyalin larinkse kaçması ancak vokal kord düzeyinin altına inmemesidir. Aspirasyonda ise materyal vokal kordları geçer. Laringeal penetrasyon bazen normal bireylerde de gözlenebilir. Mikroskopik düzeyde aspirasyonlar bile olabilir. Ancak floroskopi ya da endoskopi ile saptanabilecek boyutta aspirasyon olması patolojiktir ve artmış aspirasyon pnömonisi ya da hava yolu obstrüksiyonu riskiyle ilişkilidir (Marik 2001) . Aspirasyon, yutmadan önce,esnasında ya da sonrasında olabilir.Azalmış hiyolaringeal elevasyon, epiglottik kapanmada bozukluk, laringeal vestibülde tam olmayan kapanma veya vokal katlantıların düzgün
19 kapanmaması havayolu korunmasını sekteye uğratabilir. Bu sorunlar genelde yutma sırasında aspirasyona yol açar. Yutma öncesinde olan aspirasyon, genelde ya sıvının farinkse erken geçişinden (ağızda tutamamaya bağlı) ya da sıvı farinkse ulaştıktan sonra laringeal kapanmada gecikmeden kaynaklanmaktadır.Yutma sonrasında görülen aspirasyon ise çoğunlukla yutma sonrasında farinkste biriken rezidüden kaynaklanmaktadır.Buradaki materyal, yutma sonrasında solunum geri geldiğinde inhale edilebilmektedir. Aspirasyonun ; kaydedeğer bir etki olmamasından havayolu obstruksiyonu ya da ciddi aspirasyon pnömonisine kadar değişebilen sonuçlar doğurabilir. Aspirasyona normal yanıt güçlü bir refleks öksürük ya da boğaz temizleme hareketi (BTH)’dir.Ancak ciddi disfajili bireylerde genellikle laringeal duyu da etkilenmiştir (Garon ve ark 1996) . Sessiz aspirasyonlar ya da görünür bir yanıtın olmadığı aspirasyonlar disfaji merkezlerine başvuran olguların %25-30’unda bildirilmiştir (Garon ve ark 1996;Leder ve ark1998;Smith ve ark1999) .Aspire edilen materyalin miktarı, aspiratın havayolundaki derinliği, aspiratın içeriği (asidik materyaller kimyasal pnömonite sebep olarak akciğerlere en çok zarar verenlerdir), oral hijyen ve bireyin pulmoner temizleme mekanizmaları gibi çeşitli faktörler aspirasyonun etkilerini belirler (Palmer ve ark 2000) .
4.2.İnsanda Serebellum Ve Disfaji
Az sayıda çalışmada serebellar lezyonlu olgularda disfaji bildirilmiştir. Disfajinin olası patternleri göz önüne alındığında bu sonuçlar oldukça yetersizdir. Alberts ve ark (1992) lezyon lokalizasyonuyla disfaji semptomlarını karşılaştrmaya çalışmıştır. Yaptıkları 47 olguluk kohort çalışmalasında, 11 olguda serebellar lezyon- 6’sı büyük vasküler, 5’i küçük vasküler- saptanmıştır. Yatak başı klinik yutma testi ve videofloroskopik yutma testinin kombine edilerek kullanıldığı değerlendirmede bu 11 hastanın 7’sinde (4’ünde büyük damar ve 3’ünde küçük damar lezyonu) aspirasyon saptanmıştır. Min ve ark (1999) değerlendirdikleri 31 serebellar lezyonlu hastanın yalnızca 3’ünde disfaji semptomları bildirmiştir. Bunlardan birinde vertebral arter proksimal segment enfarktı, birinde in situ arter dal oklüzyonu ve birinde de intrakranial oklüziv hastalıkla beraber VI. alan lezyonu saptanmıştır. Zuketto ve van Gijn (2002), kompressif cerrahi sonrası semptomları tamamen düzelen, belirgin yutma güçlüğü olan serebellar ektopili bir olgu bildirmişlerdir. Disfajiyle prezente olan posterior serebellar arter anevrizmalı birer olgu bildiren Massey ve ark (1984) ve Nagahiro ve ark. (1993)’nın çalışmaları da destekleyici niteliktedir. Bu araştırıcılardan hiçbiri disfajinin şekli ya da şiddeti gibi ayrıntılara yer vermemiştir. Perie ve ark (1999) , sol serebellar hematom ve sustentoriel üçüncü ventrikül kanaması olan bir olgu sunmuştur. Bu olgunun videofloroskopik yutma testinde, yutmayı başlatmakta gecikme, bozulmuş laringeal kapanma, oral ve hipofaringeal rezidü ve aspirasyon saptanmıştır. 2006’da, Kumral ve ark. anterior ve posterior inferior serebellar arter lezyonu olan olgularda disfaji bildirmiştir. Ancak yakın zamanda yapılan bazı çalışmalar aksini iddia etmektedir. Flowers ve ark. (2011) akut iskemik inme sonrası nöroanatomik öngördürücülerle ilgili bir
meta analiz yayınlamışlardır. 17 makale incelenmiş ve meta analize dahil edilmiştir. Toplamda 656 olgu incelenmiş ve sonuç olarak serebellar lezyon sonrası disfaji insidansı % 0 bulunmuştur. Moon ve ark (2012) tarafından farklı beyin bölgelerinde inme geçirmiş 76 hastada, lezyon lokalizasyonuyla disfaji derecesi arasındaki bağlantı araştırılmıştır. Sadece medüller bölge lezyonları, yutmanın faringeal evresiyle ilgili bozukluklarla, özellikle de aspirasyonla bağlantılı bulunmuştur. Valleküler rezidü, parietotemporal inmelerle orta derecede bağlantılı bulunmuştur. Serebellar lezyonlarla disfaji semptomları arasında anlamlı bağlantı saptanmamıştır. Ancak 76 olgudan oluşan bu grupta sadece 2 kişide izole serebellar lezyon bulunduğuna dikkat etmek gerekir. Bu kadar az hasta sayısıyla anlamlı korelasyon beklenemez. Birkaç çalışmada serebellopontin (SP) köşe tümörü olan hastalarda (örn.; Thapa ve ark 2003;Wu ve ark 2011) ve SP köşe tümörü rezeksiyonu yapılmış hastalarda disfaji semptomları değerlendirilmiştir(ör.; Starmer ve ark 2012). Ancak tümör yerleşiminden dolayı serebellar doku dışında başka bölgeleri de etkilemesi nedeniyle bize spesifik olarak serebellumun disfaji üzerine etkisi hakkında bilgi verememektedir. Isono ve arkadaşları (2013) bir çalışmada spinoserebellar ataksi tip 3(SCA3) ile tip 6 (SCA6)’lı bireylerin retrospektif olarak videofloroskopik yutma testlerini inceleyerek disfaji patternlerini kıyaslamışlardır. Başlıca serebellar lezyonlu bireylerde (SCA 6) de, yaygın lezyonu olanlar (SCA 3) kadar şiddetli olmasa da, disfaji olabileceğini telkin eden bulgular elde etmişlerdir. Ancak sonuçların bireysel değişkenlik gösterebileceği unutulmamalıdır.
5.SOLUNUMUN ANATOMİSİ VE NÖROFİZYOLOJİSİ
Solunum, yaşam için mutlak gerekli gaz değişimini sağlayan kompleks sensorimotor bir fonksiyondur. Respiratuvar kemorefleksler, hava yolu reseptorleri ve akciğerler gibi çeşitli yapılardan gelen afferent geri bildirimler ışığında şekillenen özelleşmiş nöronal ağ düzeneği aracılığıyla otomatik kontrol altındadır. Bu nöronal ağda bulunan respiratuvar nöronlar , gaz alış-verişinden sorumlu pompa kaslarını ve hava akımı regulasyonundan sorumlu valf kaslarını koordine etmek üzere özelleşmiştir(Armand L ve ark. 2009). Mekanik olarak solunum , (inspirasyon sırasında) inspiratuar kasların kasılması ve (ekspirasyon sırasında) akciğerin pasif olarak eski haline dönmesinin yol açtığı intrapulmoner basıncın değişmesiyle sağlanır. Difragma ve eksternal interkostal kaslar inspiratuar kaslardır.Diafragma C3-5 spinal segment ön boynuz motor nöronlarının aksonlarından köken alan frenik sinirle innerve edilir. Eksternal interkostal kaslar T1-12 spinal segment ön boynuz motor nöronlarının aksonları olan eksternal interkostal sinirler tarafından innerve edilir. Rektus abdominis ve internal interkostal kaslar ise ekspiratuar kasları oluşturur. Bu kaslar solunum frekansının artmasıyla aktive olur. Rektus abdominis kası L1-5, internal interkostal kaslar ise T1-12 spinal segmentlerinden innervedir.
Solunum, ritmik bir aktivitedir.Bu ritmisiteyi medulla ve ponsta yerleşmiş respiratuvar merkezler sağlar. Medullada yerleşmiş pek çok respiratuvar nöronun aksonları spinal kord boyunca
21 aşağı doğru iner ve servikal, torakal, lomber bölgelerde yerleşmiş olan internöronlar ya da motor nöronlarla sinaps yapar.Medüller nöronlardan inputlar alan spinal nöronlar inspirasyon ya da ekspirasyonda görevli sinirleri oluşturur. İnspiratuvar kaslar kasılıp intrapulmoner negatif basınç oluşturduğunda atmosferden akciğere doğru hava akımı gerçekleşir. Solunumun derinliği ve frekansı önemlidir çünkü; arteriel kandaki oksijen, karbondioksit ve hidrojen iyon homeostazının sağlanmasında rol oynar. Akciğer hacmindeki değişikliğe ya da arteriel oksijen/karbondioksit/hidrojen iyon konsantrasyonuna duyarlı çeşitli reseptörler; solunumun derinliğini ve frekansını ayarlayan medüller respiratuvar nöronlara geri bildirimde bulunur. Ayrıca uyanıklık derecesi ( ör;uykuda ya da uyanık olmak gibi), sıcaklık, ağrı ve duygular da solunumu etkiler.Son olarak solunum istemli olarak korteksin de kontrolü altındadır (ör; konuşma, nefes tutma). Solunumun istemli yolakları; medüller respiratuvar merkezleri bypass ederek doğrudan spinal kordda bulunan respiratuvar motor nöronları uyarır (Mateika 2007).
5.1.Solunumla ilişkili santral merkezler:
Memelilerin alt beyin sapında bilateral organizasyonlu 3 solunum grubu tanımlanmıştır (Ballanyi 2004):
i.Dorsal respiratuar grup: Primer görevi; kemoreseptörler ve mekanoreseptörlerden gelen
verilerin medullada yerleşik olan respiratuar kontrol merkezine iletilmesidir. DRG tamamen inspiratuvar nöronlardan oluşmaktadır.DRG, 9. ve 10. Kranial sinirler ve spinal kord vasıtasıyla solunum ilişkili mekanoreseptörler ve kemoreseptörlerden afferent uyarılar alır. Daha üst serebral merkezlerden de inici afferent uyarılar almaktadır. DRG nöronları inspiriumdan hemen önce deşarj olurlar.Burada bulunan inspiratuvar nöronlar 2 çeşittir. Bunlardan biri (Iα) akciğerlerin şişmesiyle inhibe olurken , diğeri (Iβ) eksite olur.DRG nöronları aktivitelerini servikal spinal kordaki frenik motor nöronlara iletir. Ayrıca DRG inspiratuvar nöronları VRG ve PRG’ taki ekspiratuvar nöronları inhibe eder (Mateika 2007).
ii.Pontin respiratuar grup: Solunumun çeşitli fazlarında aktive olan solunum kas gruplarının
(pre/post) inspiratuar- ekspiratuar sinaptik nöronal aktivite paterninin orkestra şefidir. Pontin respiratuar grup; diyafragmanın motor kontrolü ve respiratuar ritim modulasyonundan sorumlu medial-lateral parabrakial ve Kollicker-Fuse nukleuslarından oluşur. Medial parabrakiyal nukleusta ekspiratuvar nöronlar ve lateral parabrakiyal ve Kolliker-Fuse nukleuslarında inspiratuar nöronları bulubur. PRG’nin medullayla resiprokal bağlantıları vardır. PRG’de artmış aktivite, medulladaki inspiratuvar nöronların aktivitesini kısaltır. Böylece inspirasyon kısalıp ekspirasyon daha erken gerçekleşir.Faz değişimi (phase switching) olarak adlandırılan bu aktivite solunum frekansını arttırır. Ponsun alt bölümü inspiratuvar nöronlar üzerinde eksitatuvar etki gösterir ve aktivitesi pnömotaksik merkez tarafından suprese edilir. Yine de pnömotaksik merkezin ve akciğer gerilim (stretch) reseptörlerinin uyarıları olmasa bile tam inspirasyon sonrasında inspirasyon durur. Pontin
merkezlerden hiçbir uyarı gelmediğinde medüller merkezler yavaş, ritmik ‘iç çekme’ (‘gasping’) şeklinde bir solunum paterni üretir (Mateika 2007).
iii.Ventral respiratuar grup: VRG; rostral ve kaudal gruba ayrılmıştır. VRG’un rostral
parçasında Botzinger kompleks, pre-Botzinger kompleks ve parafasial respiratuar grup bulunur.VRG, DRG’takine benzer davranış gösteren inspiratuvar ve ekspiratuvar nöronlar içerir.VRG içinde yer alan NA’ta başlıca eksternal interkostal sinirleri,11. kranial siniri, laringeal kasların motor nöronları, bronşiollerin ve kalbin parasempatik nöronlarını uyaran premotor inspiratuvar nöronlar yer alır.NRA(nukleus retro ambigualis)’nın rostral parçası (rNRA) inspiratuvar nöronlardan oluşurken, caudal parçası (cNRA) ekspiratuvar nöronlardan oluşmaktadır.VRG ekspiratuvar nöronları, ekspirasyon aktif hale gelince, ekspiratuvar kasları (abdominal ve internal interkostal) aktive eder; aynı zamanda da ekspirasyon sırasında inspiratuvar nöronları baskılamak için inhibitör sinyaller gönderir. Bu da inspiratuvar fazı sonlandırıp inspirasyonu durdurur (inspiratuvar off-switch). BotC(Bötzinger kompleks), NA’un rostralinde yer alır ve glossofaringeal ve vagal motor nöronlar dışında nerdeyse tamamen ekspiratuvar nöronlardan oluşmaktadır. NTS aracılığıyla duysal inputlar alır ve DRG,VRG ve spinal korddaki (frenik motor nöronlar) inspiratuvar nöronlar üzerinde inhibitör etkisi bulunmaktadır (Mateika 2007). Pre-BotC(Pre-Bötzinger kompleks), solunumun santral patern jeneratörünün bulunduğu bölge olarak tanımlanmaktadır. Pre-BotC, BotC ‘in kaudalinde yer alır ve sinaptik inputlar olmaksızın spontan olarak primer respiratuar ritim oluşturabilen pacemaker hücreler içerir. Pre-BotC, inspirasyon ilişkili internöronal ve motor aktivitelerin oluşturulmasından sorumludur. Buna karşın pons ve kaudal preBotC arasında yerleşik olan parafasial respiratuar grup (pFRG),ekspiratuar abdominal kasları uyaran preinspiratuar (ve postinspiratuar) aktiviteleri başlatır (Ballanyi 2004; Onimaru ve Homma 2003).
Karotid ve aortik cisimde periferik kemoreseptörler bulunur. Bu alanlarda 2 çeşit glomus hücresi bulunur.Tip 1 glomus hücreleri karotid cisimde bulunan oksijene duyarlı periferik kemoreseptörlerdir. Hipoksik uyarıyı karotid sinüs sinirine (CSN) aktarır. CSN’e gelen sinyaller NTS’un çeşitli (medial, dorsomedial, lateral ve kommissural) bölgelerine aktarılır. Daha sonra dorsal respiratuar grubun (DRG) ana nukleusu olan NTS; pontin respiratuar grup ve ventral respiratuar grubu (VRG) da içeren diğer respiratuar nöronlarla bağlantıya geçer. Botzinger komplekste bulunan ekspiratuvar nöronlar, hem medullada hem de spinal kordda bulunan inspiratuvar nöronlara inhibitör sinyaller göndererek inspiriyumu durdurur ve ekspirium pasif olarak başlar.Pre-Botzinger kompleks ve parafasial respiratuar grubun respiratuar ritmogenezden sorumlu olduğu düşünülmektedir. Buna ek olarak respiratuar nöronlar; suprapontin nukleus, mezensefalon, diensefalon, hipotalamus, serebellum, ve serebral korteks gibi beyin sapının rostralindeki alanların da etkisi altındadır. Solunumla ilişkili bazı beyin sapı bölgeleri şekil 5.1’de gösterilmiştir.
23
Şekil 5.1: Solunumla ilişkili bazı beyin sapı bölgeleri A. Koronal kesit, B. Horizontal kesit
(Mutolo ve ark, 2007; Meessen ve Olszewski’nin Atlası,1949; Shek ve ark.’nın Atlası, 1986)
Bazı beyin sapı lezyonlarında, etkilenen yapılara bağlı olarak çeşitli solunum paternleri tanımlanmıştır (Şekil 5.2).Bunlar kısaca şöyledir:
• Cheyne-Stokes Solunum: Tidal volümün önce arttığı ardından azaldığı ve kişinin apneye girdiği sikluslardan oluşan solunum paternidir. Talamus, mezodiensefalik bileşke lezyonları ya da yaygın serebral hasarda bu solunum paterni görülebilir.
• Santral Nörojenik Hiperventilasyon: Solunum sayısının dakikada 40’ı geçtiği çok yüksek frekanslı ancak düzenli bir solunum paternidir. Başlıca pontomezensefalik lezyonlarda görülebilmektedir. • Apneik Solunum: Her inspirasyonu bir apnenin takip etiiği solunum paternidir. Pons alt yarısında lateral tegmentum lezyonlarda görülebilmektedir.
• Cluster Solunum: Frekansı ve amplütüdü düzensiz, gruplar halinde inspirasyon-ekspirasyonun izlendiği bununla birlikte gruplar arasında apnenin görüldüğü solunum paternidir. Alt pontin ya da üst medüller lezyonlarda görülebilmektedir.
• Ataksik (Biot) Solunum: Son derece düzensiz bir solunum ritmidir .Dorsomedial medüller lezyonlarında görülebilmektedir.
A. Cheyne-Stokes solunumu B. Santral nörojenik hiperventilasyon. C.Apneik solunum. D, Cluster
solunum E. Ataksik (Biot) solunum (Plum F ve Posner JB,1980)
6.YUTMA VE SOLUNUMUN KOORDİNASYONU
Yutma ve solunum işlevleri için aynı kaslar ve anatomik olarak aynı pasaj (farinks) kullanılmaktadır. Bu yüzden alınan gıdaların aspirasyon riskinin en aza indirilmesinde vokal kordların adduksiyonu ve larinks elevasyonu kadar yutmanın solunumla koordineli bir şekilde yürütülmesi de önemlidir. Yutma-solunum ilişkisi pek çok çalışmada incelenmiştir. Farklı çalışmalarda farklı yöntemler kullanılsa da (Martin ve ark 1994, Martin-Harris, 2006 ve Martin-Harris, 2008, McFarland ve Lund, 1995, Nishino ve ark 1985, Nishino ve Hiraga, 1991, Preiksaitis ve ark 1992, Preiksaitis ve Mills, 1996, Selley ve ark 1989 ve Smith ve ark 1989), yutma-solunum ilişkisine dair karakteristik bulgular açıkça ortaya konmuştur:1) solunum siklusu içinde yutmanın zamanlaması rastgele değildir; yutmaların çoğu ( ≥%80 ) ekspiratuar fazda başlar, 2) her tekli (single bolus) yutmaya 1-2 saniye süreli kısa bir apne eşlik eder; 3) genelde yutma sonrasında ekspirasyon gerçekleşir ve bir sonraki solunum döngüsü gecikir.
Beyin sapı respiratuvar nöral ağı; yutma, derin nefes alma (augmente solunum), ekspiratuar refleks, aspirasyon refleksi, asfiksik yanıt, apne, hapşırma, laringeal adduksiyon ve öksürük gibi çeşitli havayolu koruyucu davranışlarda rol alır (Baekey ve ark 2001; Baekey ve ark 2004; Bolser ve ark 2006; Ertekin ve ark 2003; Saito ve ark 2003). Tüm bu havayolu koruyucu davranışlar için ;solunum patterninin normal solunumdan koruyucu reflekse özgü koordineli nöromusküler patterne dönüşmesi gerekmektedir. Bu reflekslere ait her bir spesifik davranışın beyin sapındaki nöronal jeneratörünün, bir mekanizmayla respiratuar nöronal ağın kontrolünü ele geçirdiği varsayılmaktadır (Bolser ve ark 2006).Bu mekanizma, davranışsal kontrolün montajı - behavioral control assemblies- (BCA) olarak tanımlanmaktadır. Beyin sapı kontrol sistemlerinin kararları doğrultusunda, farinksten ve larinksten gelen duysal girdilerin bombardımanı altındayken bile, her bir davranış düzenli ve koordineli bir şekilde yürütülür( Pitts ve ark 2010; Pitts ve ark 2011).
Yutmanın solunum paternini şekillendirdiği Jafari ve ark. (2003) tarafından gösterilmiştir.Buna göre yutma; inspiratuar aktivitede kesilme, kısa bir ekspirasyon ve ardından submental EMG aktivitesi sırasında apne; bolusun farinks boyunca hareketinin göstergesi olan hipoglossal basınç artışı, bunu takiben ekspiratuar hava akımı ve ventilatuar solunum paterninin geri gelmesiyle sonuçlanır. Böylece normal solunum paterni rekonfigüre edilerek üst hava yolunu aspirasyondan koruyan, aynı zamanda da bolusun farinks ve özofagusa geçişine izin veren bir yutma-solunum paterni oluşturulmuş olur. Dick ve ark. (1993), süperior laringeal sinirin (SLN) yutma ortaya çıkaracak şiddette uyarılmasının, ekspirasyonun geç fazında uzamaya yola açarak inspirasyonu inhibe ettiğini bildirmişlerdir. Saito ve ark (2002) SLN uyarımının yapay bir yutma oluşturduğunu ve frenik sinir inspiratuar aktivitesini inhibe ederek eş zamanlı olarak hipoglossal sinir (XII. K.S.) aktivitesini aktive ettiğini göstermişlerdir.
Yutma-25 solunum paterni için inspiratuar inhibisyonun, bolusun larinkse kaçmasını (aspirasyon) önleyip farinksi geçmesine yetecek kadar süren bir yutma apnesinin elzem olduğu görülmektedir. Bu nedenle yutma-solunum paterninde; beyin sapı yutma patern jeneratörünün aktivasyonu respiratuar nöral ağ üzerinde etki eder.Bu etkiyle respiratuar nöral paternin solunum kaslarına giden uyarısı değişerek yutma apnesi oluşturulur. Böylece respiratuar nöral ağ yeniden şekillendirilmiş (rekonfigüre edilmiş) olur. Solunumun nörojenezinde rol oynayan elemanların çoğu aynı zamanda öksürme, kusma gibi solunum paterninde modülasyon gerektiren başka reflekslerde de görevlidir(Baekey ve ark 2001; Baekey ve ark 2004; Shannon ve ark 2000). Birbirinden tamamen farklı 2 davranışın nörojenezinde rol oynayan ortak elemanlar Lindsey ve ark.(1992) ve Shannon ve ark.(2000) tarafından rekonfigürasyon olarak tanımlanmaktadır. Rekonfigürasyon kavramı, fonksiyonel olarak bağlantılı bir nöral ağın; deşarj patternlerini yeniden düzenleyerek ve motor outputlar üzerine etkilerini değiştirerek, birden fazla davranış oluşturabildiğini öne sürmektedir. Rekonfigürasyon, sistemin yeniden organize olarak farklı bir davranış oluşturabilme yetisi kazanmasıdır (Bolser ve ark 2006). Ek olarak solunum fazlarının zamanlamasına yönelik incelemeler, tekrarlayan öksürük sırasında oluşan yutmanın, takip eden solunum fazının ekspirasyon süresini uzatarak modüle ettiğini göstermiştir ( Pitts ve ark 2010; Pitts ve ark 2011). Saito, ve ark.(2002a,b) XII. K.S. aktivitesiyle eşzamanlı olarak ; solunum fazı zamanlamasında inspiratuar faz süresinin inhibisyonu ve ekspiratuar fazın uzamasının, respiratuar nöral ağdaki ekspiratuar nöronların depolarize ve repolarize olduğu fonksiyonel döngüyle sağlandığını bildirmektedirler. Ayrıca; bu ağdaki bazı nöronların yutma uyarısına yanıt vermediğini de belirtmektedirler (Saito ve ark 2002a). Bu sonuçlar; yutma BCA’sının, respiratuar ağ nöronlarını düzenleyen, respiratuar kontrol sisteminde yutmaya uygun nöral programın devreye sokulmasını sağladığını göstermektedir. Yutmanın solunum patterni üzerindeki nöral kontrol mekanizmalarını ortaya koymak için deney hayvanlarının medullalarında mikroelektrod kaydı yöntemlerinin kullanıldığı çeşitli araştırmalar yapılmıştır. Öncelikle nöronlar, solunum sırasındaki aktivitelerine göre sınıflandırılmıştır (Shannon ve ark 1998) .İnspiratuar fazda uyarım yapan nöronlar I, ekspiratuar fazda uyarım yapan nöronlar E, ekspirasyondan inspirasyona geçiş fazında uyarım yapan nöronlarsa E/IE olarak belirtilmiştir. Respiratuar fazlara göre uyarım yapmayan nöronlara ise respiratuar olarak modüle olmayan -non respiratory modulated- (NRM) nöronları denmektedir. Ayrıca nöronlar yükseltici (AUG) veya indirgeyici (DEC) aktivite paternlerine göre de sınıflandırılmaktadır (Shannon ve ark 1998) . Nöron bazında yapılan kayıtlamalarda yutmanın E-DEC nöronlarının bir sub-populasyonundaki aktiviteyi arttırırken diğer E-DEC nöronlarındakini inhibe ettiği gösterilmiştir(Saito ve ark 2003). Aynı şekilde pek çok E-AUG respiratuar nöronunun farklı yutma yanıtı özellikleri vardır. Örneğin bazıları yutma motor aktivitesi esnasında inhibeyken, yutmalar arasındaki ekspiratuar dönemde aktive olur (Saito ve ark 2003). Bu yüzden E-DEC nöronlarının bir sub-populasyonu yutmanın motor fazında aktive olur, ekspiratuar faz boyunca aktif kalır ve inspirasyon sırasında inhibe olur. E-AUG populasyonunun başka bir sub-populasyonu yutmanın motor fazında inhibe olur ancak
yutmanın olmadığı geç ekspiratuar fazda aktive olur. Tek tek kayıtlanan inspiratuar nöronlar, yutmanın motor fazında inhibe olur ve geç ekspiratuar fazda bir sub-populasyonu inhibe olurken, I nöronlarının başka bir sub-populasyonu aktive olur (Saito ve ark 2003). Ertekin a ve Aydogdu (2003) beyin sapındaki nöral aktivite kayıtlamalarına dayanarak yutmanın solunum paterni üzerindeki kontrolünün; NTS’da yerleşik DSG ile ventral lateral medullada yerleşik VSG arasındaki etkileşimle sağlandığını öne sürmektedir. DSG, supramedüller merkezlerden ve periferik duysal uyaranlardan aktive edici girdiler alır. Daha sonra DSG, yutmanın motor paternini oluşturmak için VSG ile etkileşir. VSG’nin, yutma ve solunumla ilişkili kasları kontrol eden motor nöronlara efferent projeksiyonları vardır. Bu yutmanın solunum üzerindeki kontrolü kavramı, yutma BCA’sıyla sağlanmaktadır (Bolser ve ark 2006). Yutma BCA’sı, meduller respiratuar nöral ağdaki mevcut elemanların düzenlenmesini, rekonfigürasyonunu ve yutmaya özgü kaslara medüller motor outputlar gönderilmesini sağlar.
Özetle; yutmanın solunum üzerindeki kontrolünün; devreye giren yutma nöral elemanlarının mevcut respiratuar nöral ağ elemanlarının deşarj paternlerini değiştirerek onları yeniden görevlendirmesiyle olduğu öne sürülmektedir.Yutma santral patern jeneratörünün devreye girmesi de mevcut elemanları suprese ederek nöral ağdaki rolünü ortadan kaldırır ya da etkisizleştirir. Daha önce sessiz olan elemanların devreye sokulması a) premotor sistemlere yeni,farklı bir sürüm (drive) ekleyebilir b) temel elemanların yerine farklı aksonal bağlantıları olan elemanların ikame edilmesini sağlayabilir. Yutma, dorsal ve ventral yutma gruplarındaki yutma nöronlarının devreye girmesiyle başlatılır. Yutma patern jeneratörü bir kez başlatıldığında; sessiz halde bekleyen kasları devreye sokarak ve aktif haldeki kasları, özellikle solunum kaslarını, susturarak stereotipik bir motor pattern oluşturur. Yutmanın solunum üzerindeki kontrolü; her ikisinin de iç içe geçmiş nöral elemanları sayesinde solunumsal nöral çıktıların yeniden düzenlendiği , rekonfigüre edildiği bir nöral (network) ağdır. Bolus; farinkse, hem yutma hem de solunum için ortak olan alana giriş yaptığında, respiratuar güdümün sınırları içinde, respiratuar motor pattern yutmaya hizmet etmeye başlar. Yutmanın respiratuar nöral ağı rekonfigüre etmesi, havayolunun korunması için elzemdir ve diğre havayolu koruyucu sistemlerle entegre çalışır. Bu nedenle yutma, solunum, öksürük ve diğer havayolu koruyucu refleksler bir çeşit santral nöral motor sistem tarafından oluşturulur ve ortak elamanları paylaşırlar. Respiratuar ağ, çoğu havayolu korunmasında önemli çeşitli davranışlar için rekonfigüre olur,değişim gösterir. Ancak yutma ve havayolu koruyucu refleksler arasındaki bu önemli koordinasyon tam olarak anlaşılamamıştır. Yutma ve havayolu koruyucu sistemlerin; bu davranışları, havayolu güvenliğini sağlamak üzere, kusursuzca koordine eden BCA sistemiyle kontrol edildiği varsayılmaktadır (Davenport ve ark 2011). Sonuç olarak respiratuar CPG’ne ait bazı nöral elemanlar; farklı davranışlara hizmet eden başka kontrol merkezlerince de ortak olarak kullanılmaktadır. Yaşam boyu spontan olarak aktif olan respiratuar motor döngünün aksine, non respiratuar davranışların belli afferent inputlarla tetiklenmesi gerekir. Aynı zamanda; bu non-respiratuar davranışlar bir kez tetiklendiğinde stereotipik bir motor döngü başlatılır ve afferent geribildirimlere ihtiyaç duymaz.
27 Ancak tüm bu santral programlı motor döngüler , fizyolojik şartlara bağlı olarak afferent inputlarla module edilebilmektedir. Şekil 6.1’de birbirine antagonist fonksiyonlardan sorumlu ancak tamamen ya da kısmen aynı motor outputları kullanan solunum, öksürük, yutma ve kusma CPG’lerinin kesişimi gösterilmektedir. Bu multifonksiyonel pattern jeneratör devrelerinin nöromodulasyonu ile davranışsal esneklik sağlanabilmektedir. Bu kavram omurgasızlarda gösterilmiş ve memelilerde kabul görmüştür (Dickinson, 2006 ;Briggman ve Kristan, 2008).
Şekil 6.1.: Solunum, öksürük, yutma ve kusma CPG’leri arasındaki dinamik etkileşimin şematik görünümü. Afferent ya da santral inputlar, koruyucu ve ekspulsive refleksleri tetiklemek üzere
respiratuar CPG’yle etkileşir. Çoğu respiratuar nöron hem solunum hem de öksürmede rol oynar; bu da CPG’lerinin büyük oranda kesiştiğinin göstergesidir. Yutma ve kusma büyük oranda , solunum CPG’ünün dışında ,ayrı CPG’lere bağlıdır. Ancak bazı respiratuar nöronlar (şekilde taralı alanlarla gösterilmiştir) aynı zamanda non respiratuar davranışlara da hizmet ederek, CPG’leri arasında gösterilmiş parsiyel kesişimi açıklar. (Bianchi, A. L. ve Gestreau, C. 2009)
7.SPİNOSEREBELLAR ATAKSİ
Spinoserebellar ataksi (SCA) , eski adıyla otozomal dominant serebellar ataksi (ADCA); hem klinik hem de genetik yönden heterojen; otozomal dominant kalıtımlı nörodejeneratif bir hastalıktır. Nöropatologlar SCA’yı, serebellum ve onun afferent ve efferent bağlantılarının dejenerasyonuna bağlı olarak değişik derecelerde beyin sapı ve spinal kordun da etkilendiği bir çeşit herediter serebellar ataksi sendromu olarak tanımlamaktadır(Schöls ve ark 2004; Taroni ve DiDonato 2004).Prevalansı ülkeden ülkeye farklılık göstermekle birlikte tüm dünyada 0.9-3 in 100,000 civarındadır (Klockgether 2008) . Ancak bu alanda çok fazla çalımanın olmaması ve tanı almamış SCA olgularının olabileceği göz önünde bulundurulmalıdır. Ağustos 2016’da internet veri bankasında yapılan arama sonucuna göre (http://neuromuscular.wustl.edu/ataxia/domatax.html) genetik olarak tanımlanmış 40 SCA alttipi (SCA1-SCA43) bulunmaktadır. Bunlar tipik klinik özellikleriyle birlikte tablo 7.1 ’de görülmektedir.