ÜÇ FARKLI SICAKLIĞIN Acanthoscelides obtectus Say, 1931
(Coleoptera:Bruchidae)
ERGİNLERİNİN YAĞ ve YAĞ ASİDİ MİKTARINA ETKİSİ
EVRİM SÖNMEZ DOKTORA TEZİ BİYOLOJİ ANABİLİM DALI
T.C.
ONDOKUZ MAYIS ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
ÜÇ FARKLI SICAKLIĞIN Acanthoscelides obtectus Say,1931 (Coleoptera:Bruchidae) ERGİNLERİNİN YAĞ ve YAĞ ASİDİ
MİKTARINA ETKİSİ
EVRİM SÖNMEZ
DOKTORA TEZİ BİYOLOJİ ANABİLİM DALI
DANIŞMAN
PROF. DR. ADEM GÜLEL
T.C.
ONDOKUZ MAYIS ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
Bu çalışma, jürimiz tarafından 12/05/2010 tarihinde yapılan sınav ile Biyoloji Anabilim Dalında DOKTORA tezi olarak kabul edilmiştir.
Başkan: Prof. Dr. Adem GÜLEL
Üye :Prof.Dr.Halil KORKMAZ
Üye :Prof.Dr.İbrahim ÖZKOÇ
Üye :Doç.Dr.Ülya NURULLAHOĞLU
Üye :Yar.Doç.Dr.Eylem AKMAN GÜNDÜZ
ONAY:
Yukarıdaki imzaların adı geçen öğretim üyelerine ait olduğunu onaylarım. / /
Prof. Dr. Hasan GÜMÜŞ
ÜÇ FARKLI SICAKLIĞIN Acanthoscelides obtectus Say, 1931 (Coleoptera: Bruchidae) ERGİNLERİNİN YAĞ ve YAĞ
ASİDİ MİKTARINA ETKİSİ ÖZET
Üç farklı sıcaklıkta A. obtectus Say, 1931 (Coleoptera: Bruchidae) erginlerinde toplam yağ, toplam yağ asidi ile yağ asitlerinin çeşit ve miktarlarının yaşa ve eşeye bağlı olarak değişimi incelendi. Denemeler 15, 20 ve 30±2ºC ve % 65±5 nisbi nem içeren laboratuar şartlarında yapıldı. Denemeler sırasında herhangi bir fotoperyot rejimi uygulanmadı. Erginlerin sahip oldukları yağ asitlerinin çeşit ve miktarları gaz kromatografisi ile tayin edildi.
Çalışılan tüm sıcaklıklarda toplam yağ miktarı her iki eşeyde yaş ve sıcaklık artışına paralel olarak azaldı. Toplam yağ asidi miktarları genel olarak sıcaklık artışına paralel olarak artmıştır. Buna karşılık toplam yağ asidi miktarları yaş artışına bağlı olarak önce artmış sonra azalmıştır. A. obtectus erginlerinde kaprilik asit, kaprik asit, laurik asit, miristik asit, palmitik asit, palmitoleik asit, stearik asit, oleik asit, linolelaidik asit, linoleik asit, linolenik+11-eikosenoik asit, 11,14-eikosadienoik asit, cis-13,16-dokosadienoik asit ve cis-5,8,11,14,17-eikosapentaenoik asit olmak üzere 15 yağ asidi tespit edildi. Bunlardan oleik ve palmitik asit dominant yağ asitleri olarak belirlendi. Üç farklı sıcaklıkta genel olarak tespit edilen yağ asitlerinin çeşitleri sabit kalırken, yağ asitlerinin miktarının sıcaklığa, eşeye, yaşa ve yağ asidi çeşidine bağlı olarak değiştiği tespit edilmiştir.
Anahtar Kelimeler: Acanthoscelides, Acanthoscelides obtectus, sıcaklık, toplam yağ, yağ asitleri.
THE EFFECT OF THREE DIFFERENT TEMPERATURES ON THE LIPID AND FATTY ACID AMOUNT IN Acanthoscelides obtectus Say, 1931
(Coleoptera:Bruchidae) FOR ADULTS ABSTRACT
At three different temperatures the total lipids, total fatty acids and fatty acid types and amounts were investigated at differrent age groups in both sexes of A. obtectus Say, 1931 (Coleoptera: Bruchidae) adults. The study was carried out under laboratory conditions at 15, 20 and 30±2o
The amount of total lipids decreased in both sexes when age and temperature increased. However, amount of total fatty acids generally increased with temperature, but it firstly increased and then decreased with age. Fifteen different fatty acids were determined in A. obtectus adults. They were capryrlic acid, capric acid, lauric acid, myristic acid, palmitic acid, palmitoleic acid, stearic acid, oleic acid, linolelaidic acid, linoleic acid, linolenic+cis-11-eicosenoic acid, cis-11,14-eicosadienoic acid, cis-13,16-docosadienoic acid ve cis-5,8,11,14,17-eicosapentaenoic acid. Two of them oleic and palmitic acids were found to be dominant. The types of fatty acid present were the same at the three different temperatures but their amounts changed depending on temperature, sex, age and the fatty acid type.
C and 65±5 % relative humidity. No specific photoperiodic regimen was used throughout the study. The types and amount of fatty acids in adults were determined by gas chromotography.
Key words: Acanthoscelides, Acanthoscelides obtectus, temperature, total lipid, fatty acids
TEŞEKKÜR
Tez konumun belirlenmesinde yardımcı olan, çalışmalarım sırasında desteğini esirgemeyen, fikirleriyle çalışmalarıma yön veren danışman hocam Sayın Prof. Dr. Adem GÜLEL’e teşekkür ederim.
Tez izleme komitemde bulunan ve çalışmalarım süresince beni yönlendiren hocalarım Sayın Prof. Dr. İbrahim ÖZKOÇ ve Sayın Prof. Dr. Halil KORKMAZ’a,
Gaz kromatografisi analizleri sırasında laboratuarında çalışma fırsatı veren ve analizler sırasında desteğini esirgemeyen, fikirleri ile çalışmalarıma yön veren Veteriner Fakültesi’nden Sayın Prof. Dr. Abdurrahman AKSOY, Yrd.Doç. Dr. Oğuzhan YAVUZ, Yrd. Doç. Dr. Kürşad DAŞ, Arş. Görevlileri Dilek GÜVENÇ ve Enes ATMACA’ya,
Laboratuar analizleri sırasında tecrübelerini ve desteğini esirgemeyen Sayın Doç. Dr. Ülya NURULLAHOĞLU ve Arş. Gör. Rahile ÖZTÜRK’e,
Verilerin istatistiksel analizleri sırasında yardımcı olan Arş. Görevlileri N. Alp ERİLLİ ve Ufuk YOLCU’ya,
Laboratuar çalışmalarım ve tez yazım aşamasında yardımlarını, fikirlerini ve manevi desteğini hiç eksik etmeyen Biyoloji Bölümü Arş. Görevlileri Özgür Varer IŞITAN, Dr. Arzu GÜRSOY ve Ali BOZ’a, her türlü desteklerinden dolayı Biyoloji Bölümü öğretim üyelerine ve araştırma görevlilerine,
Çalışmalarım sırasında tez projeme destek veren Ondokuz Mayıs Üniversitesi Bilimsel Araştırma Projeleri Komisyonu’na,
Destek ve teşvikleri ile bana yardımcı olan Ramazan AKBULUT’a, sevgili anne ve babama teşekkür ederim.
İÇİNDEKİLER
1. GİRİŞ ... 1
2. GENEL BİLGİLER ... .11
2.1. Sıcaklığın Böceklere Etkileri ... ….11
2.2. Karbohidratların ve Proteinleri Böcekler İçin Önemi ... 14
2.3. Lipitler ... 17
2.4. Gaz Kromatografisi ... 29
3. MATERYAL ve YÖNTEMLER ... 31
3.1. Zararlı Stok Kültürlerinin Kurulması ... 31
3.2.Analizler için Numunelerin Hazırlanması ... 32
3.2.1. Toplam Yağ Miktarının Belirlenmesi ... 32
3.2.2. Yağ Asitlerinin Serbetleştirilmesi ve Toplam Yağ Asidi Miktarının belirlenmesi32 3.2.3. Yağ Asidi Metil Esterlerinin Hazırlanması ... 33
3.3.Gaz Kromatografisi ile Numunelerin Analizi ... 34
3.3.1. Gaz Kromatografi Cihazının (Shimadzu 17A GC-FID) Çalışma Koşulları ... 34
3.3.2. Miks (Karışım) Yağ Asitleri Standartının Hazırlanışı ... 35
3.3.3. Elde Edilen Kromatogramların Değerlendirilmesi ... 35
3.4. Elde Edilen Verilerin İstatistiksel Olarak Değerlendirilmesi ... 35
4. BULGULAR ... 36
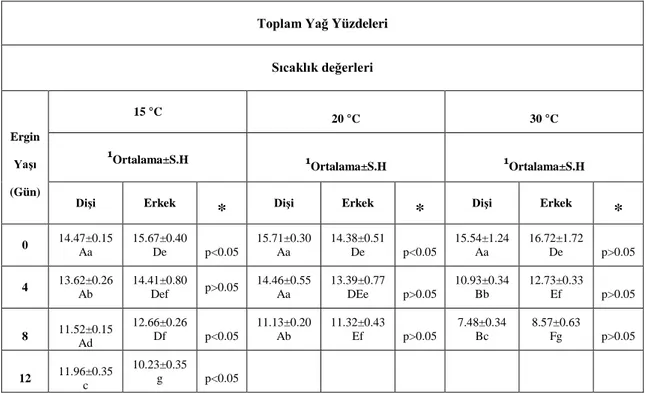
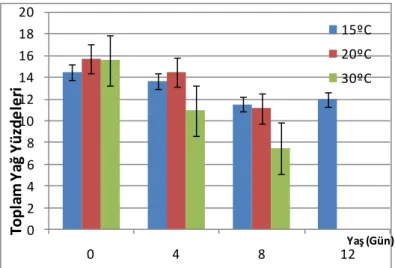
4.1. A. obtectus Erginlerinde Birey Başına Düşen Toplam Yağ Yüzdeleri ... 36
4.1.1. A. obtectus Erginlerinde Birey Başına Düşen Toplam Yağ Yüzdelerinin Yaşa Bağlı Olarak Değişimi ... 37
4.1.2. A. obtectus Erginlerinde Birey Başına Düşen Toplam Yağ Yüzdelerinin Sıcaklığa Bağlı Olarak Değişimi ... 38
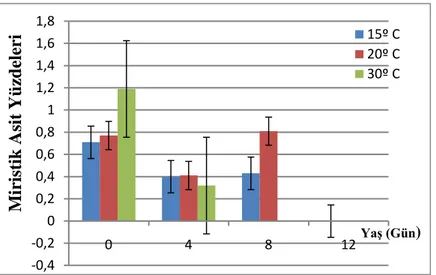
4.2. A. obtectus Erginlerinde Birey Başına Düşen Toplam Yağ Asidi Yüzdeleri ... 39
4.2.1. A. obtectus Erginlerinde Birey Başına Düşen Toplam Yağ Asidi Yüzdelerinin Yaşa Bağlı Olarak Değişimi ... 41
4.2.2. A. obtectus Erginlerinde Birey Başına Düşen Toplam Yağ Asidi Yüzdelerinin Sıcaklığa Bağlı Olarak Değişimi ... 42
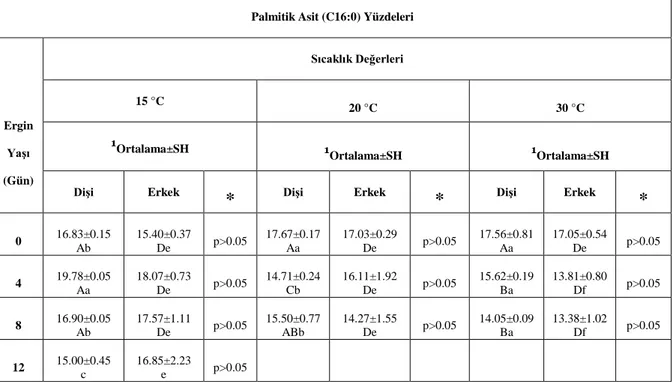
4.3. Üç Farklı Sıcaklıkta A. obtectus Erginlerindeki Yağ Asitlerinin Çeşit ve Miktarları43 4.3.1. A. obtectus Erginlerinde Yağ Asitlerinin Çeşit ve Miktarlarının Yaşa Bağlı Olarak Değişimi ………..43
4.3.2. A. obtectus Erginlerinde Yağ Asitlerinin Çeşit ve Miktarlarının Sıcaklığa Bağlı
Olarak Değişimi ... 63
4.3.3. Gaz Kromatografisi Analizlerinde Elde Edilen Kromatogramlar ... 75
5. TARTIŞMA ... 84
6. SONUÇ ve ÖNERİLER ... 96
7. KAYNAKLAR ... 97
ŞEKİLLER LİSTESİ
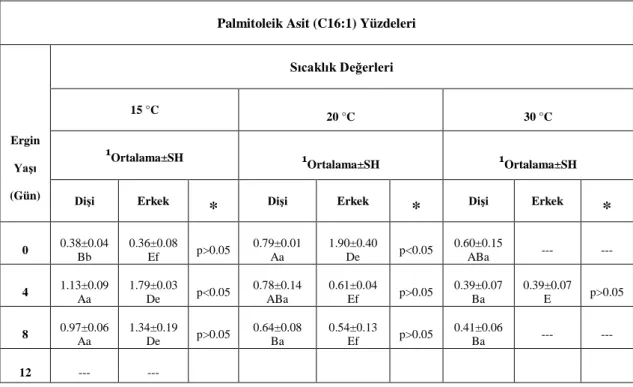
Şekil 2. 1. A. obtectus’un gelişim evreleri ... 25 Şekil 2. 2. A. obtectus larvalarının fasulye tanelerine verdiği zarar ... 26 Şekil 4.1. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen toplam yağ yüzdeleri ... 37 Şekil 4.2. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen toplam yağ yüzdeleri ... 37 Şekil 4.3. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen toplam yağ asidi yüzdeleri ... 40 Şekil 4.4. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen toplam yağ asidi yüzdeleri ... 41 Şekil 4.5. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen laurik asit yüzdeleri ... 45 Şekil 4.6. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen laurik asit yüzdeleri ... 45 Şekil 4.7. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen miristik asit yüzdeleri ... 47 Şekil 4.8. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen miristik asit yüzdeleri ... 47 Şekil 4.9. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen palmitik asit yüzdeleri ... 49 Şekil 4.10. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen palmitik asit yüzdeleri ... 49 Şekil 4.11. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen palmitoleik asit yüzdeleri ... 51 Şekil 4.12. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen palmitoleik asit yüzdeleri ... 51 Şekil 4.13. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen stearik asit yüzdeleri ... 53 Şekil 4.14. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen stearik asit yüzdeleri ... 53
Şekil 4.15. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen oleik asit yüzdeleri ... 54 Şekil 4.16. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen oleik asit yüzdeleri ... 55 Şekil 4.17. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen linolelaidik asit yüzdeleri ... 56 Şekil 4.18. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen linolelaidik asit yüzdeleri ... 57 Şekil 4.19. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen linoleik asit yüzdeleri ... 58 Şekil 4.20. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen linoleik asit yüzdeleri ... 59 Şekil 4.21. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen linolenik asit+cis-11-eikosenoik yüzdeleri ... 60 Şekil 4.22. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen linolenik asit+cis-11-eikosenoik yüzdeleri ... 61 Şekil 4.23. A. obtectus dişilerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen cis-11,14-eikosadienoik yüzdeleri ... 62 Şekil 4.24. A. obtectus erkeklerinde sıcaklığa ve yaşa bağlı olarak birey başına düşen cis-11,14-eikosadienoik yüzdeleri ... 63 Şekil 4.25. Supelco FAMEs mix yağ asitleri standardının kromatogramı ... 76 Şekil 4.26. 15°C'da dört günlük A. obtectus dişilerine ait yağ asidi kromatogramı ... 78 Şekil 4.27. 15°C'da dört günlük A. obtectus erkeklerine ait yağ asidi kromatogramı .... 79 Şekil 4.28. 20°C'da dört günlük A. obtectus dişilerine ait yağ asidi kromatogramı ... 80 Şekil 4.29. 20°C'da dört günlük A. obtectus erkeklerine ait yağ asidi kromatogramı .... 81 Şekil 4.30. 30°C'da dört günlük A. obtectus dişilerine ait yağ asidi kromatogramı ... 82 Şekil 4.31. 30°C'da dört günlük A. obtectus erkeklerine ait yağ asidi kromatogramı .... 83
ÇİZELGELER LİSTESİ
Çizelge 2. 1. Depolanmış ürün zararlılarının sıcaklıklara verdiği tepkiler ... 11 Çizelge 4.1. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen toplam yağ yüzdeleri ... 36 Çizelge 4.2. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen toplam yağ asidi yüzdeleri ... 40 Çizelge 4.3. A. obtectus erginlerinde tespit edilen yağ asitleri ... 43 Çizelge 4.4. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen laurik asit yüzdeleri ... 44 Çizelge 4.5. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen miristik asit yüzdeleri ... 46 Çizelge 4.6. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen palmitik asit yüzdeleri ... 48 Çizelge 4.7. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen palmitoleik asit yüzdeleri ... 50 Çizelge 4.8. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen stearik asit yüzdeleri ... 52 Çizelge 4.9. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen oleik asit yüzdeleri ... 54 Çizelge 4.10. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen linolelaidik asit yüzdeleri ... 56 Çizelge 4.11. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen linoleik asit yüzdeleri ... 58 Çizelge 4.12. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen linolenik asit+cis-11-eikosenoik asit yüzdeleri ... 60 Çizelge 4.13. A. obtectus erginlerinde sıcaklığa, yaşa ve eşeye bağlı olarak birey başına düşen cis-11,14-eikosadienoik asit yüzdeleri ... 62 Çizelge 4.14. Standart Yağ Asitlerine Ait Alıkonma Zamanları ve Alanları ... 77
1. GİRİŞ
Canlılar varlıklarını devam ettirmek için devamlı olarak çevreleri ile madde ve enerji alışverişi yapan açık sistemlerdir. Canlı ihtiyaç duyduğu birtakım maddeleri çevresinden alırken, bir kısmını gerçekleştirdiği metabolik faaliyetlerle kazanır (Carrol ve ark., 2003). Canlılar aleminde geniş bir yer tutan böcekler poikliotermik canlılardır. Bu nedenle sıcaklık bunların metabolik faaliyetlerinde enzimler üzerine olan etkisinden dolayı önemli rol oynar (Chen ve ark., 2005; Benkova ve Volf, 2007). Sıcaklık artışı belirli sınırlar içerisinde kaldığı zaman enzimatik aktiviteyi olumlu yönde arttırır (Fields, 1992; Somero, 1995; Have, 2008). Böceklerin ergin öncesi gelişimleri ve ergin hayatlarını sürdürmek için ihtiyaç duydukları sıcaklık dereceleri türlere göre değişebilir. Bu durum onların değişken iklim koşullarında hayatta kalabilmeleri için kazandıkları en önemli adaptasyonlardan birisidir (Gilbert ve Raworth, 1996; Amarasekare ve ark. 2008). Bu adaptasyon sayesinde böcekler coğrafik dağılımlarını ve davranışlarını değiştirerek değişen iklim ve yeni iklim koşullarına hızlı bir şekilde tepki verebilirler (Akbulut, 2000; Carrol ve ark., 2003; Xian-Hui ve Kang, 2005; Zhang ve ark., 2008).
Diğer canlılarda olduğu gibi aşırı düşük veya aşırı yüksek sıcaklıklar, böceklerde de önemli fizyolojik tepkilere neden olur (Carrol ve ark., 2003; McMillan ve ark., 2005 Coracini ve ark., 2007). Böceklerdeki beslenme (Chandrakantha ve ark., 1987; Logan ve ark., 2007), metabolizma (Kistler, 1982, Chown ve Nicolson, 2004; Morgan, 2004; Klowden, 2007), gelişim hızı (Kistler, 1982; Lale ve Vidal, 2003a; Lale ve Vidal, 2003b; Dupuis ve ark., 2006), ömür uzunluğu (Allsopp, 1981; Prasantha ve ark., 2002; Honek ve ark., 2003; Johnson ve Valero, 2003; Ferizli ve ark., 2004) ve üreme (Hagstrum ve Clarence, 1973; Kistler, 1982; Lale ve Vidal, 2003a; Keena, 2006) gibi faaliyetler sıcaklığa bağlı olarak önemli derecede değişir.
Fasulye zararlıları olan Callosobruchus maculatus (Coleoptera: Bruchidae) ve Acanthoscelides obtectus (Coleoptera: Bruchidae)’nın ölüm oranlarına farklı sıcaklık ve nem oranlarının etkilerinin ele alındığı çalışmada bu türlerin ölüm oranlarının sıcaklıkla doğru orantılı, nem oranı ile ters orantılı olarak arttığı, ayrıca bu iki türden A. obtectus’un düşük sıcaklığa toleransının daha fazla olduğu tespit edilmiştir (Prasantha ve ark., 2002).
Chandrakantha ve ark. (1987), dört farklı sıcaklıkta (20, 25, 30 ve 35 ºC) üç farklı fasulye türü (Vigna unguiculata, Phaseolus radiatus ve Dolichos lab lab) ile
beslenen C. maculatus erginlerinde verim ve larvalarda gelişim sürelerini incelemişlerdir. Çalışmalar sonucunda üç farklı besinde sıcaklık arttıkça larvaların besin tüketiminin azaldığı ve gelişim süresinin kısaldığı belirlenmiştir. Ayrıca yumurta üretiminin, yumurtaların ağırlığının ve enerji miktarının besin çeşidinden etkilenmediği fakat sıcaklığın artması ile yumurta üretiminin azaldığı tespit edilmiştir.
Kistler (1982), 20-50 ºC arasında değişen sıcaklıkların altı Coleoptera türünde (Stator generalis, S. limbatus, S. pruininus, S. sordidus, Mimomestes amicus ve Algarobius prosopis) metabolik hıza, gelişim hızına ve verimlerine etkilerini incelemiştir. Çalışılan tüm türlerin metabolik hızlarının, gelişim hızlarının ve verimlerinin türlere göre farklılık gösterdiği fakat tüm türlerde gelişim hızı ve metabolik hızın sıcaklık artışı ile arttığı ve yüksek sıcaklıklarda erginlerin kısa sürede öldüğü gözlenmiştir. Sıcaklığa bağlı olarak verimin türler arasında farklılık gösterdiği S. generalis hariç diğer çalışılan türlerde maksimum veriminin 30 ºC’da, S. generalis türünde ise 35 ºC’da olduğu tesbit edilmiştir.
C. maculatus ve C. subinnotatus erginlerinin ovipozisyon ve gelişimlerine dört farklı sıcaklık (25, 30, 35 ve 40 °C) ve üç farklı nem oranının (%30, 60 ve 90 r.h.) etkilerinin ele alındığı çalışmada; farklı sıcaklık ve nem oranlarının bu türlerin erginlerinde gelişim ve ovipozisyonu önemli derecede etkilediği bildirilmiştir (Lale ve Vidal, 2003a). C. maculatus erginleri en fazla yumurtayı 35 ve 40 °C’da bırakmış ve bırakılan yumurta sayısı sıcaklıkla doğru orantılı olarak artmıştır. C. subinnotatus erginleri ise en fazla yumurtayı 30 ve 35 °C’da bırakmıştır. Her iki türde en az yumurta sayısı ekstrem sıcaklıklarda ortaya çıkmıştır. Bırakılan yumurta sayısını farklı nem oranlarının önemli derecede etkilemediği gözlenmiştir. En fazla yumurta gelişimi 30 °C’da kaydedilmiştir. Bırakılan yumurtaların gelişim süreleri her iki türde sıcaklık ve nem tarafından önemli bir şekilde etkilenmiştir. C. maculatus ve C. subinnotatus’da yumurtadan ergine kadar geçen gelişim süresi 25 °C ve %30 r.h.’de, 30 ve 35 °C’a göre yaklaşık olarak iki kat daha uzun sürmüştür (Lale ve Vidal, 2003a).
Ferizli ve ark. (2004)’nın depo ürün zararlısı C. maculatus ile yaptıkları çalışmada düşük sıcaklıkların yumurta, larva ve erginler üzerine olan etkileri incelenmiştir. Yumurta, larva ve erginler 20, 40, 60, 80, 100, 120, 140, 160 ve 180 dakika -18 ºC’a maruz bırakılmışlardır. Larva ve yumurtaların 100 dakika ve üzerinde -18 ºC’a maruz bırakılmaları halinde gelişimlerini tamamlayamadıkları, bunun dışındaki diğer zaman aralıklarına tolerans gösterdikleri, tüm bu gelişim aşamalarında 180 dakika
-18 ºC’a maruz bırakma işleminin bu zararlının kontrolünde alternatif bir yöntem olarak kullanılabileceği öne sürülmüştür. Böcekler ergin öncesi gelişim basamaklarında bir evreden diğerine geçerken belirli bir sıcaklık derecesine ihtiyaç duyarlar. Dupuis ve ark. (2006), çok düşük sıcaklıkların A. obtectus larval aşamalarına ve erginleri üzerine etkilerini incelemişlerdir. Çalışma sırasında 2., 3. ve 4. evre larvaları, prepup, pup ve erginleri farklı zaman aralıklarında farklı sıcaklıklara (-10 ºC’da 2, 4, 6,12 saat ve 0 ºC’da 4, 8, 16, 32 gün) maruz bırakılmıştır. Farklı evrelerdeki larva, pup ve erginleri direkt 0 ºC’a maruz bıraktıklarında ölüm oranının 6. günden itibaren %50 civarında olduğu, direkt -10 ºC’a maruz bıraktıklarında 1. ve 2. evre larvalarının 4 saat sonra, diğer evrelerdeki larva ve pupaların ise 6 saat sonra öldükleri ve -22 ºC’da 16 saat sonunda tüm bireylerin öldüğünü tayin etmişlerdir.
Ponsonby ve Copland (1996), Chilocorus nigritus (Coleoptera: Coccinellidae) ile yaptıkları çalışmada yedi farklı sıcaklığın (18, 20, 22, 24, 26, 28 ve 30 ºC) bu türün gelişim aşamalarına (yumurta, larva ve pupa) etkilerini incelemişlerdir. 18 ºC’da 1. evre larvaların öldükleri ve genellikle tüm larval aşamaların gelişim sürelerinin artan sıcaklıkla beraber kısaldığı tespit edilmiştir. Yumurtadan ergin oluncaya kadar olan gelişim süresinin en kısa 30 ºC’da olduğu, ergin olan böceklerin hayatta kalma yüzdelerinin en fazla 28 ve 30 ºC’da gözlendiği, erginlerin eşey oranının sıcaklıktan etkilenmediği ve çalışılan her sıcaklıkta yumurtadan ergin oluncaya kadar geçen sürelerin sıcaklıklara göre değişiklik gösterdiği bulunmuştur.
Jackson ve Elliot (1988), Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) ile yaptıkları çalışmada 15-33 ºC sıcaklık aralıklarında yetiştirildiğinde sıcaklıkla beraber gelişim hızının arttığını ve tüm sıcaklıklarda eşeyler arasında gelişim hızında farklılık olduğunu, erkeklerin dişilerden daha hızlı geliştiğini, 33 ºC’da yumurta açılımı olmadığını, larvaların gelişemediklerini ve bu tür için optimum sıcaklığın 21-30 ºC olduğunu tespit etmişlerdir. Gastrophysa viridula (Coleoptera: Chrysomelidae) ile yapılan çalışmada dört farklı sıcaklığın bu türün yumurta, larva ile pupa gelişim süresi ve ömür uzunluğuna etkileri incelenmiştir. Artan sıcaklıkla beraber gelişim, preovipozisyon, ovipozisyon sürelerinin ve ergin ömür uzunluğunun kısaldığı tespit edilmiş ve eşeyler arasında fark gözlenmemiştir (Honek ve ark., 2003).
Hayvanlar aleminde proteinler, lipitler ve karbohitratlar çok değişik amaçlar ve faaliyetler için kullanılmaktadır (Stryer, 1975; Nelson ve Cox, 2005; Chown ve Nicolson, 2004; Morgan, 2004; Klowden, 2007). Böcekler büyüme, gelişme ve üreme
faaliyetlerini devam ettirmek için protein, karbohidrat ve lipit gibi temel besin maddelerine gereksinim duyarlar (Chippendale, 1972; Haynes, 1985;Tomic-Carruthers, 2007). Bu nedenle böceklerde de bu temel besin maddeleri ile ilgili araştırmalar her zaman ilgi çekici olmuştur. Çünkü bu konuda elde edilen bilgiler, bunların çok farklı metabolik işlevlerinin, hayvandaki homeostasisin devamlılığını, temel maddelerin metabolik ilişkilerini anlamamızda önemli rol oynamaktadır. Böcekler ihtiyaç duydukları maddelerin bir kısmını doğal yaşamlarında çevrelerindeki çeşitli kaynaklardan karşılarken, bir kısmını da gerçekleştirdikleri metabolik faaliyetler ile sentezlerler.
Chippendale (1972), Sitophilus oryzae (Coleoptera: Curculionidae) ile yaptığı çalışmada farklı besinlerle (nişasta, dekstrin, amilopektin, glikojen, amiloz, selüloz, inülin, mono ve disakkaritler) beslenmenin ömür uzunluğuna etkilerini incelemiştir. Polisakkaritlerle beslenen grubun hayatta kalma oranı ve ömür uzunluğunun diğer gruba göre daha yüksek olduğunu tespit etmiştir.
Böceklerin besin gereksinimleri türlere göre farklılıklar gösterdiği gibi (Nettles ve ark., 1972) aynı türde gelişim evrelerine, eşeye ve çevresel faktörlere göre de farklılıklar gösterir (Tomic-Carruthers, 2007). Anthonomus grandis (Coleoptera: Curculionidae) ile yapılan çalışmada diyapozda olanların olmayanlara göre trehaloz miktarları arasında çok büyük farklar olmadığı, 12-20 saat aç bırakılan erginlerde trehaloz miktarının oldukça düşük olmasına rağmen diyapozda olanlardan daha yüksek olduğu ve glikojenin metabolize edilme hızının erginlerde diyapozda olanlardan daha fazla olduğu tespit edilmiştir (Nettles ve ark., 1972).
Böceklerin besinsel ihtiyaçlarının beslenmeye bağlı olarak da değiştiği bilinmektedir (Canavoso ve ark., 2001). Bazı böcek türlerinde ergin evrede ihtiyaç duyulan besinlerin, yumurtadan veya larval evrede depolanan besin rezervlerinden karşılandığı ifade edilmiştir (Wigthman, 1978; Pant ve ark., 1979; Giron ve Casas, 2003; Behmer, 2006; Colinet ve ark., 2006). Callosobruchus analis ile yapılan çalışmada yeni ergin olmuş erginlerin enerji içeriklerinin larval aşamalarda depolanan lipitlerden dolayı diğer yaşlardaki erginlere göre çok daha fazla olduğu ve bu lipit miktarının yaşa bağlı olarak azaldığı tespit edilmiştir (Wigthman, 1978).
Gündüz ve ark. (2008), yumurta veya larva döneminde kazanılan besinlerin ergin evrede beslenmeyen ve ergin hayat süreleri kısa olan türlerde yeterli olabildiğini ileri sürmüşlerdir. Buna karşılık ergin yaşam süresi uzun olan türlerde yumurta veya
larva döneminde kazanılan besinler yeterli olmadığından bu türler yaşama ve üreme ile ilgili faaliyetlerini devam ettirebilmek için ihtiyaç duydukları protein, karbohidrat, lipit ve vitamin gibi maddeleri genellikle bitki özsuyu, nektar, polen veya konak hemolenflerinden karşılarlar (Wackers ve ark., 2007; Gündüz ve ark., 2008; Gindin ve ark., 2009). Nektarla beslenme genellikle Diptera, Lepidoptera, Hymenoptera ve bazı Coleoptera ordolarında yaygındır (Nicolson, 1998; Wackers ve ark., 2007).
Önemli enerji kaynağı olan karbohidratların, böceklerin ömür uzunluğunu ve üreme faaliyetlerini olumlu yönde etkilediği bilinmektedir (Chippendale, 1972; Haynes, 1985; Ellis ve ark., 2002; Kaakeh, 2005; Tomic-Carruthers, 2007). A.grandis (Coleoptera: Curcuilionidae) ile yapılan çalışmada %2.5 ile %10 sükroz ve %10 maltoz içeren besinle beslenen erginlerin ölüm oranlarının farklılık gösterdiği tespit edilmiştir. Ölüm oranının %10’luk sükrozla beslenen dişiler ile %10’luk maltozla beslenen erkeklerde daha düşük olduğu bulunmuştur (Haynes, 1985). Kaakeh (2005), Rynchophorus ferrugineus (Coleoptera: Curculionidae) ile yaptığı çalışmada doğal besin (şeker kamışı, palmiye yaprağı ve sapı) ve değişik doğal maddelerin karışımı ile hazırlanan besinle (yulaf, patates, ananas, yulaf-palmiye lifi, yulaf-patates, yulaf-ananas besinleri bakto agar, şeker, tuz ve maya) beslenmenin bu türün yumurtlama süresini, larval ve pupal sürelerini, ergin erkek ve dişilerin gelişim sürelerini ve hayat döngülerini önemli ölçüde etkilediğini tespit etmiştir. Doğal besinlerle beslenen grupların ömür uzunlukları ve ergin gelişim süreleri diğer gruptan daha uzun bulunmuştur.
Hylobius transversovittatus (Coleoptera: Curculionidae) ile yapılan çalışmada sükroz, nişasta ve üç farklı sindirilemeyen karbohidrat içeren yapay besinle beslenen erginlerin büyüme ve larvaların hayatta kalmalarına etkileri incelendiğinde; sükroz konsantrasyonunun artmasının larvaların hayatta kalma oranlarını düşürdüğü, besinlerinden sükroz çıkarıldığında veya yerine nişasta eklendiğinde 1. evre larvaların ölüm oranının arttığı, besinlerinde sindirilebilir karbohidrat miktarı arttırıldığında gelişimini tamamlayan larva oranının düştüğü tespit edilmiştir (Tomic-Carruthers, 2007).
Yüksek organizasyonlu hayvanlarda olduğu gibi böceklerde de enerji rezervi olarak kullanılan glikojen, genellikle yağ doku ve uçma kaslarında depolanır. Böceğin beslenme rejiminin kalitatif ve kantitatif bileşimi glikojenin depolanma düzeyini önemli derecede etkilemektedir (Sharma ve Sharma, 1980; El Atta, 2000). Sharma ve Sharma (1980), C. maculatus ile Zabrotes subfasciatus (Coleoptera: Bruchidae) ile yaptıkları
çalışmada Z. subfasciatus erginlerinin diğer türden daha fazla lipide sahip olduklarını ayrıca iki türün glikojen miktarlarının yaşa bağlı olarak azaldığını tespit etmişlerdir.
Bir tohum zararlısı olan Bruchidius uberatus (Coleoptera: Bruchidae) ile yapılan çalışmada farklı besinlerle (%1’lik sükroz, sadece su ve kontrol grubu) beslenmenin bu türün verimine, ömür uzunluğuna ve yumurta bırakma süresine etkileri incelenmiştir. Tüm deneme gruplarında %1’lik sükrozla beslenen grubun, hiç beslenmeyen kontrol grubundan ve sadece su ile beslenen gruptan daha uzun yumurta bırakma süresine sahip olduğu ve bırakılan yumurta ile açılan yumurta sayısının, verimin ve ömür uzunluğunun daha fazla olduğu tespit edilmiştir (El Atta, 2000).
Alphitobius diaperinus (Coleoptera: Tenebrionidae) ile yapılan çalışmada sentezlenen ürün miktarının sıcaklığa bağlı olarak değiştiği tespit edilmiştir. 28 ºC’da bulunan dişilerdeki prolin, asparjin ve serin amino asit miktarlarının erkeklerden daha düşük, tirozin ve lizin miktarlarının daha yüksek olduğu fakat 10 ºC’a maruz bırakılan erginlerde 28 ºC’dakilerin aksine total aminoasit miktarının arttığı tespit edilmiştir (Renault ve ark., 2006).
Birçok böcek eşeysel olgunluğa ulaşma ve yumurta üretimi için lipitlere gereksinim duymaktadır (Nemec, 2002). Böcekler lipitleri besin yoluyla alabildikleri gibi metabolik ilişkiler yoluyla vücutta depolanmış olan protein ve karbohidratlardan da sentezleyebilirler. Bu nedenle bunlardaki karbohidrat ve protein miktarları dolaylı da olsa sahip oldukları lipit ve yağ asitlerinin miktarını etkiler. Son yıllarda değişik araştırıcılar tarafından değişik ordolara dahil olan Lyctus planicollis, Tenebrio molitor, Dermestes maculatus, Mobosteira lobulifera, Photinus pyralis, Magicicada septendecim, Ostrinia nubilalis ile yapılan çalışmalarla böceklerin bazı doymuş, doymamış veya aşırı doymamış yağ asitlerini sentezleyebildiklerini ortaya çıkarılmıştır (Mauldin ve ark., 1971; Cohen, 1974; Stanley-Samuelsen ve ark.,1988; Khebbeb ve ark., 1997; Hoback ve ark.,1999; Aktümsek ve ark., 2000; Aliza ve ark., 2001; Arrese ve ark., 2001; Çakmak ve ark., 2005; Çakmak ve ark., 2007; Gereszek ve ark., 2008; Visser ve Ellers, 2008; Due ve ark., 2009).
L. planicollis (Coleoptera: Bostrichidae)’da gelişim süresince yağ asitlerinin çeşidinin değişmediğini fakat bunların miktarlarının gelişim evrelerine göre değişiklik gösterdiği tespit edilmiştir. Lipit içeriği embriyonik gelişim sırasında sabit kalırken, larvaların çıkışı sırasında azalmış, larval evrede tekrar artmıştır. Larval evrede artan lipit miktarı, larvadan pupa, puptan ergine geçiş sırasında kullanılmıştır. Düşük yağ içerikli
besinlerde larvalar, glukozdan linoleik asit ve sterolleri sentezleyemedikleri için gelişememişlerdir. Bu sonuca göre bu maddelerin bu tür için esansiyel olduğu sonucuna varılmıştır (Mauldin ve ark., 1971).
Rhynchophorus palmarum (Coleoptera: Curculionidae) larvalarının toplam yağ ve yağ asitlerinin incelendiği çalışmada derideki toplam yağ içeriğinin 35.16 (g/100g), sindirim kanalı içeriğindekinin 49.05 (g/100g) olduğu, deri ve sindirim kanalı içeriği yağ asidi çeşitlerinin miristik, miristoleik, palmitik, stearik, oleik ve linoleik asit olduğu ve en fazla miktarda palmitik (40.44 g/100g) ve oleik asit (46.71 g/100g) bulunduğu tespit edilmiştir (Due ve ark., 2009).
Böceklerin gelişim ve büyümelerinde özellikle etkili olan iki değişken faktör sıcaklık ve besindir. Genel olarak düşük sıcaklıklar böceklerde gelişim süresini uzatır, gelişim hızını ve verimi azaltıp, ergin büyüklüğünü artırır (Rodriguez-Saona ve Miller, 1999; Cymborowski, 2000; Petersen ve ark., 2000; Smith, 2002; Renault ve ark., 2003; Chen ve ark., 2005; Colinet ve ark., 2006; Stillwell ve ark., 2007). Buna karşılık kalitatif ve kantitatif olarak yetersiz olan besinler gelişim süresini uzatıp, gelişim hızı, ergin büyüklüğü ve verimi azaltırken, erginlerde bazı anatomik bozukluklara neden olmaktadır (Moczek, 1998; Takakura, 2004; Fox ve ark., 2006; Tomic-Carruthers, 2007; Bong ve ark., 2008; Gonzalaez-Teuber ve ark., 2008).
Sıcaklık ve beslenme birçok yönden canlıyı etkiler. C. maculatus (Coleoptera: Bruchidae) ile yapılan çalışmada düşük kaliteli besinlerle beslenen larvaların, yüksek kaliteli besinlerle beslenen larvalara göre erginleştikleri zaman daha küçük yumurtalar bıraktıkları ve daha kısa ömür uzunluklarına sahip oldukları tespit edilmiştir (Gonzalaez-Teuber ve ark., 2008). Hippodamia convergens (Coleoptera: Coccinellidae) ile yapılan çalışmada dört farklı sıcaklığın (18, 22, 26 ve 30 ºC) bu türün yumurta, larva ve pupa gelişim süresini etkilediği, ölüm oranının düşük sıcaklıkla beraber arttığı tespit edilmiştir. Pupa ve 4. evre larvalarının ölüm oranının en fazla 18 ºC’da olduğu, pupa ve ergin ağırlıklarının sıcaklıkla beraber değişiklik gösterdiği, en ağır pupa ve erginlerin 22 ºC’da belirlendiği ve bu erginlerin kanatlarının diğer sıcaklıklara göre daha büyük olduğu bulunmuştur (Rodriguez-Saona ve Miller, 1999).
Chen ve ark. (2005), Pseudacteon tricuspis’de ömür uzunluğu, erginlerde vücut büyüklüğü ve kanat uzunluğu üzerine sıcaklık, besin, eşey ve çiftleşmenin etkilerini incelemişlerdir. Çalışma sonucu farklı sıcaklıkta (20, 25, 28 ve 33 °C) şekerle beslenen erginlerin şekerle beslenmeyenlere göre, 20 °C’da hem beslenen hem de beslenmeyen
dişi ve erkeklerin diğer sıcaklıklarda yetiştirilenlere göre daha uzun ömür uzunluğuna sahip oldukları, ömür uzunluğunun sıcaklıkla ters orantılı olarak değiştiğini ve vücut büyüklüğünün ömür uzunluğuna olumlu katkısı olduğunu tespit etmişlerdir.
Canlılar, türe göre değişmekle birlikte metabolik aktivitelerini değiştirerek, vücut sıcaklıklarını düzenleyerek veya mikrohabitatlarını değiştirerek, sıcaklık değişimine belirli sınırlar içinde tolerans gösterebilirler (Cox ve ark., 2007).
Hızlı nüfus artışı diğer sosyoekonomik sorunlar ve ihtiyaçlar yanında tarım ürünlerine olan ihtiyacı da arttırmaktadır. Tarım ürünlerinde artış sağlamak için birim alandan elde edilen ürün miktarını artırmak amacıyla değişik yöntemler uygulanmaktadır. Fakat sadece elde edilen ürün miktarının artırılması yeterli değildir. Elde edilen ürünün kalite ve kantitesinin zarar görmeden saklanması da önemli bir sorun olmaktadır. Günümüzde karşılaştığımız biyotik ve abiyotik faktörlerin olumsuz etkileri, aşırı kentleşme gibi sorunlar tarım alanlarının ve elde edilen ürünlerin verimli bir şekilde değerlendirilmesini zorlaştırmaktadır. Biyotik faktörlerden biri olan zararlı böcekler orman, tarım alanları veya ürünlerin depolandığı ambarlarda büyük ölçüde kalite ve kantite azalmasına sebep olmaktadır (Ertürk ve ark., 2004).
Yemeklik baklagiller (fasulye, nohut, börülce, barbunya, bezelye ve mercimek), yüzyıllardan beri insan beslenmesinde önemli yer tutan ve bu nedenle yaygın olarak kullanılan tarım ürünlerindendir (Velten ve ark., 2007; Çetin ve ark., 2009). Üretici tarafından hasat edilen baklagiller kendileri veya bunları ticari amaçla alanlar tarafından tohumluk, yemeklik veya hayvan yemi olarak kullanılmaktadır. Fakat gerek üreticilerin depolarında gerekse ticari depolarda depolanan tüm baklagiller için küf mantarlarından kaynaklanan aflatoksin oluşumu dışında en büyük problemlerden birisi de ambar ve depolarda çoğalabilen ve yılda birçok döl verebilme yeteneğine sahip olan baklagil tohum böcekleridir (Schmale ve ark, 2002; Parsons ve Credland, 2003). Depolanmış ürünler 600’den fazla böcek türü tarafından istilaya uğrar ve bu istilacı türler ürünlere kalitatif ve kantitatif olarak zarar verirler (Rajendran ve Sriranjini, 2008). En önemli tohum zararlıları Coleoptera, Diptera, Lepidoptera, Hymenoptera ve Hemiptera ordolarına ait türlerdir (Silva ve ark., 2007). Bu ordolara ait türler yüksek adaptasyon yeteneğine sahip olduklarından insanlar ve diğer yollarla taşınan tohumlarla coğrafik olarak geniş bir dağılım alanına sahip olmuşlardır (Sing ve Argobast, 2008; Tuda ve ark., 2009).
Fasulye tohum böceği A. obtectus, kuru fasulye tanelerinin depolanması esnasında hem kalitatif hem de kantitatif olarak çok büyük ürün kayıplarına neden olan bir zararlıdır (Keats ve ark., 1998; Odagiu ve Porca, 2003; Porca ve ark., 2003a; 2003b; 2003c; Rojas-Rousse ve ark., 2007). A. obtectus yılda birden çok döl verir. Dişi erginler fasulye tanelerinin üzerine tek tek veya 4-20'lik kümeler halinde 200 kadar yumurta bırakırlar (Parsons ve Credland, 2003; Silva ve ark., 2007). Yumurtadan çıkan larvalar, baklagil taneleri içinde beslenmeleri süresince oyuklar meydana getirerek tanenin besin değerini düşürdükleri gibi dışkı ve larval vücut artıkları ile de kirletirler. Bu türde yumurtadan ergin oluncaya kadar geçen gelişim süresi 100-110 gün, ergin ömrü sıcaklık ve neme bağlı olarak değişmekle beraber genellikle 7-14 gündür (Yılmaz ve Elmalı, 2002; Ghidiu, 2005). Erginler için en uygun sıcaklıklar 27-29 ºC’dır. Daha düşük ve daha yüksek sıcaklıklar ömür uzunluğunu olumsuz yönde etkiler (Johnson ve Kistler, 1987; Tamer, 1996, http://www.agroatlas.ru/en/content/pests/Acanthoscelides_obtectus, 15.02.2010).
Tohum böceklerinin hem arazide hem de depolarda verdiği zararları önlemek için üreticiler ve depo sahipleri bu zararlılara karşı genellikle kimyasal mücadele yaparlar (Porca ve ark., 2003a,b; Nazzi ve ark., 2008). Zararlı kontrolünde kimyasal maddelere ağırlık verilmesi kimyasalların hedef dışı bir şekilde direkt olarak veya besin zinciri yoluyla hem insan hem de diğer canlılara verdiği zararlardan dolayı önceden beklenilenin aksine zamanla birçok problemin ortaya çıkmasına neden olmuştur. Bu nedenle kimyasalların birçoğunun kullanımı bırakılmış veya yasaklanmıştır. Son yıllarda ekolojik bilincin artması sonucu kimyasal mücadeleye alternatif olabilecek, entegre mücadele yöntemleri kullanılmaya başlamıştır. Entegre mücadele yönteminde doğal dengenin korunmasını sağlayan ve çevreye hiçbir olumsuz etkisi olmayan biyolojik mücadele yöntemleri önemli bir yer tutmaktadır (Aslan ve ark., 2006). Biyolojik mücadelede başarı, mücadele stratejilerinin ve zamanlamasının belirlenmesine bağlıdır. Bu durum hem zararlının hem bu zararlıya karşı kullanılacak olan biyolojik kontrol ajanının biyolojik özelliklerinin bilinmesi ile sağlanmaktadır. Bir tohum zararlısı olan A. obtectus ile yapılacak mücadelede de öncelikle zararlının biyolojisinin, biyolojik özelliklerini etkileyen çevresel faktörlerin bilinmesi gerekmektedir (Baldassari ve ark., 2005; Gaylord ve ark., 2008).
A. obtectus ile ilgili yaptığımız literatür taramasında besin çeşitlerinin ve farklı sıcaklık derecelerinin yumurta açılım oranına, ergin öncesi gelişim süresine,
preovipozisyon zamanına, oviopzisyonuna, verimine, ergin hayat uzunluğuna, ölüm oranına, farklı eşeylerin gelişim hızına ve vücut büyüklüğüne etkilerinin yeteri kadar araştırıldığı, fakat yağ asitleri ile ilgili olarak sadece erginlerin kutikulasındaki yağ asitlerinin ele alındığı, farklı sıcaklık değerlerinin değişik yaşlardaki erginlerde toplam yağ, toplam yağ asitleri ile yağ asitlerinin çeşit ve miktarına etkisinin ele alınmadığı görülmüştür (Szentesi, 1981; Pfaffenberger, 1985; Tucic ve ark., 1995; 1998; Yılmaz ve Elmalı, 2002; Odagiu ve Porca, 2003; Papachristos ve Stamopoulos, 2004; Tucic ve ark., 2004; Alvarez ve ark., 2006; Dupius ve ark., 2006; Romano ve ark., 2006; Sapunaru ve ark., 2006; Velten ve ark., 2007; Golebiowski ve ark., 2008; Seslija ve ark., 2008). Bu nedenle bu çalışmada yaşa bağlı olarak üç farklı sıcaklığın A. obtectus erginlerindeki toplam yağ, toplam yağ asitleri ile yağ asitlerinin çeşit ve miktarına etkilerinin değerlendirilmesi amaçlanmıştır. Elde edilen verilerin böceklerdeki lipit metabolizması konusundaki boşlukları doldurmaya ve A. obtectus ile yapılacak mücadele stratejileri ile zamanlamasının tayinine katkıda bulunacağı beklenmektedir.
2.GENEL BİLGİLER
2.1. Sıcaklığın Böceklere Etkileri
Canlılar için üç sıcaklık zonu (bölgesi) tanımlanır. Birincisi gelişiminin ve üremenin maksimum seviyede gerçekleştiği Optimum Zon, ikincisi canlının hayat döngüsünü tamamlayabildiği fakat biyolojik aktivitelerinin daha düşük olduğu optimum zonun altındaki veya üstündeki Suboptimum Zon’dur. Üçüncüsü ise canlının ölümüne neden olan ve suboptimum zondaki sıcaklık değerlerinin altında veya üstünde yer alan Letal Zon’dur (Fields, 1992). Fields (1992), depolanmış ürün zararlısı böceklerde optimum, suboptimum ve letal zon sıcaklık değerlerinin böcekleri nasıl etkilediğini tespit etmiştir (Çizelge 2.1)
Çizelge 2.1. Depolanmış ürün zararlılarının sıcaklıklara verdiği tepkiler (Fields, 1992).
Zon Sıcaklık (ºC) Etki
Letal 50-60 45 Dakikalar içerisinde ölüm Saatler içerisinde ölüm Suboptimum 35 33-35 Gelişimin yavaşlaması Gelişimin durması
Optimum 25-33 Maksimum gelişim oranı
Suboptimum 13-25 13-20 Gelişimin yavaşlaması Gelişimin durması Letal 5 -10,-15 -25,-15
Hareketliliğin durması, günler içerisinde ölüm
Haftalar veya aylar içerisinde ölüm
Böceğin donması, dakikalar içerisinde ölüm
Optimum sıcaklıkta birçok böcek larvası türüne öz sabit bir gelişme göstermektedir. Fakat optimum sıcaklıklarda meydana gelen değişiklikler böceğin metabolizmasını, üreme kapasitesini, beslenme alışkanlıklarını ve bunlara bağlı olarak da yayılışını etkilemektedir (Howe ve Currie, 1964; Tunaz, 2004; Xian-Hui ve Kang, 2005). Howe ve Currie (1964), sıcaklığın altı depo ürün zararlısının (A. obtectus, C. maculatus, C. chinensis, C. analis, Callosobruchus rhodesianus ve Z. subfasciatus)
gelişim hızı, ölüm oranı ve yumurtlamasına etkilerini incelemişler ve her tür için optimum sıcaklıkları belirlemişlerdir. A. obtectus’da yumurta açılımının 17,5-30 ºC arasında %80, 15 ile 35 ºC’da %70 oranında olduğu, 40 ºC’da hiçbir yumurtanın açılmadığı, yumurtaların ve larvaların en hızlı gelişimi için optimum sıcaklığın 30 ºC olduğu, ergin çıkışının ise 15 ºC’da çok az, 35 ºC’da hiç olmadığını tespit etmişlerdir. Ayrıca dişilerin gelişim süresinin ve ağırlıklarının erkeklerden daha fazla olduğu, eşey oranının sıcaklıktan etkilenmediği, en ağır bireylerin en düşük sıcaklıklarda tespit edildiği, artan sıcaklıkla beraber ağırlıklarının azaldığı, en fazla yumurtayı (ortalama 70) 25 ºC’da ergin oldukları günden 2-3 gün sonra bıraktıkları belirtilmiştir.
Üç farklı sıcaklıkta (20, 25 ve 30 ºC) yapılan çalışmada beş farklı fasulye genotipinin (Arc.1S, Arc.1, Arc.2, Arc.4 ve Carioca Pitoco) A. obtectus istilasına dirençlerinde sıcaklığın etkileri incelenmiştir. Arc.1S ve Arc.2 dışındaki diğer genotiplerin direncinin 25 ve 30ºC’da sıcaklıkla beraber azaldığı ve A. obtectus istila oranının arttığı, 20 ºC’da fasulye direncinin en fazla olduğu ve 20 ºC’ın depolanma için en uygun sıcaklık olduğu belirtilmiştir (Baldin ve Lara, 2004).
Böceklerin tolerans gösterebileceği sıcaklık sınırları içerisinde olmak kaydıyla genelde artan sıcaklıklar böceklerin yumurtlama oranını, yumurta sayısını ve larval gelişim aşamalarını hızlandırır ve ergin ömrünü kısaltır (Howe ve Currie, 1964; Lale ve Vidal, 2003b). C. maculatus ve C. subinnotatus ile yapılan çalışmada üç farklı sıcaklığa (40, 45 ve 50 ºC) farklı zaman dilimlerinde (1, 2, 4 ve 6 saat) maruz bırakılan bu türlerin yumurta bırakma, gelişim ve hayatta kalma oranları incelenmiştir. 30 ºC’da yetiştirilen ergin dişiler belirtilen zaman aralıklarında belirtilen sıcaklıklara maruz bırakılmışlardır. Her iki türde bırakılan yumurta sayısı ile elde edilen ergin sayısının sıcaklıkla ve sıcaklıklara maruz bırakılma süresi ile ters orantılı olarak azaldığı, en fazla 40 ºC en az 50 ºC’da yumurta bıraktıkları, erginlerde ölüm oranının en fazla 50 ºC’da görüldüğü ve C. maculatus’un yüksek sıcaklığa diğer türden daha duyarlı olduğu tespit edilmiştir. Ayrıca 50 ºC’a 4 ve 6 saat maruz bırakılan yumurtalarda gelişme olmadığı belirtilmiştir (Lale ve Vidal, 2003b).
Genellikle sıcaklık değişimine bağlı olarak böceklerde iki tip davranış şekli görülür. Birincisi sıcaklık arttıkça böcekte biyolojik tepki artar ve bu durum ölüm sınırına kadar sürer. İkincisi ise belli bir noktaya kadar sıcaklıktaki artış böceğin biyolojik tepkisini yükseltir fakat belirli bir noktadan sonra bu tepki azalır (Akbulut, 2000; Pervez ve Omkar, 2004; Keena, 2006).
Keena (2006), iki farklı bölgeden topladığı Anoplophora glabripennis (Coleoptera: Cerambycidae) erginleri ile yaptığı çalışmada, yedi farklı sıcaklığın (5, 10, 15, 20, 25, 30 ve 35 o
Propylea dissecta (Coleoptera: Coccinellidae)’da değişen sıcaklıkların (20-35 ºC) bu türün özellikle verimini ve yumurtlamasını etkilediği tespit edilmiştir. Bu tür için optimum sıcaklığın 27 ºC olduğu, 27 ºC’da artan metabolizma hızı ile birlikte gonadların daha erken olgunlaştığı ve en fazla yumurtanın bu sıcaklıkta bırakıldığı bulunmuştur. 27 ºC’dan sonraki sıcaklıklarda ise yumurtlamanın geciktiği, gonad olgunlaşmasının sıcaklıktan etkilendiği ve daha az yumurta bıraktıkları tespit edilmiş, bu türün veriminin, ovipozisyonunun yüksek ve düşük sıcaklıklardan önemli derecede etkilendiği belirtilmiştir (Pervez ve Omkar, 2004).
C) üreme, yumurta açılımı ve erginlerin hayatta kalma sürelerine etkilerini incelemiştir. Farklı sıcaklıkların, her iki bölgeden toplanan dişilerde verimi önemli derecede etkilediğini, verim bakımından iki bölgeden toplanan dişiler arasında farklılık olduğunu ileri sürmüştür. Düşük sıcaklıkta (10 ºC) tutulan erginlerin daha yavaş hareket ettiğini, daha az beslendiğini, çiftleşmediklerini ve yumurtlamadıklarını, 5 ºC’da tutulanların birçoğunun öldüğünü fakat 15-25 ºC’da tutulanların normal davranışlar sergilediğini tespit etmiştir. Erginlerin sıcaklık arttıkça özellikle 30 ºC’da çok hızlı hareket ettikleri, çok fazla beslendikleri ve uçtukları, 35 ºC’da ise bu davranışların tekrar azaldığı, çiftleşme ve yumurta bırakma davranışlarının gözlenmediği tespit edilmiştir.
Düşük sıcaklıklar da yüksek sıcaklıklar gibi böceklerin birçok metabolik faaliyetini etkiler (McAvoy ve Kok, 1999; Smith, 2002; Baldassari ve ark., 2005; Xian-Hui ve Kang, 2005).
Smith (2002), Ncirophorus investigator (Coleoptera: Sliphidae)’da büyük olan larvalardan küçük larvalara göre daha büyük erginlerin meydana geldiğini, kışı larva olarak geçiren erginlerin, kışı ergin olarak geçirenlerden daha büyük olduğunu ve hayatta kalma oranlarının daha yüksek olduğunu tespit etmiştir. Aynı tür ile yapılan başka bir çalışmada beş farklı sıcaklığın (3, 7,12, 17 ve 27 ºC) bu türün üremesine ve hayatta kalmasına etkileri incelenmiştir (Baldasari ve ark., 2005). 3 ve 7 ºC’da ölüm oranının diğer sıcaklıklara göre daha fazla olduğunu, 12 ve 17 ºC’daki böceklerin metabolik hızlarının yavaşlamasından dolayı 27 ºC’a göre ölümlerinin daha yavaş ve ölüm oranının daha az olduğunu, böceklerin 7-27 ºC arasında yumurtlayabildiklerini tespit etmişlerdir. Johnson ve Valero (2003), C. maculatus yumurta, larva, pup ve
erginlerini -18 ºC’a belirli zaman aralıkları (0, 80, 100 ve 120 dakika) ile maruz bıraktıklarında düşük sıcaklığa en yüksek toleransın yumurta evresinde, en düşük toleransın ise erginlerde olduğunu bulmuşlardır.
H. transversovittatus (Coleoptera: Curculionidae)’da değişik sıcaklıkların verim ve ergin ömür uzunluğunu etkilediği, en fazla yumurta bırakmanın 20-25 ºC, yumurta açılma oranının 20-30 ºC arasındaki sıcaklıklarda olduğu ve artan sıcaklıkla beraber ömür uzunluğunun azaldığı tespit edilmiştir (McAvoy ve Kok, 1999).
Sıcaklıkta oluşabilecek büyük değişikliklerin, böceklerde yayılma özelliklerini ve göç edenlerde göç yollarını değiştirebileceği, bunun da pek çok zararlı kontrol problemini beraberinde getireceği ileri sürülmektedir (Akbulut, 2000).
2.2. Karbohidratlar ve Proteinlerin Böcekler için Önemi
Karbohidratlar, lipitler ve proteinler canlıların varlıklarını sürdürebilmelerinde değişik amaçlar için kullandıkları önemli maddelerdir (Stryer, 1975; Morgan, 2004; Nelson ve Cox, 2005; Behmer, 2006). Böcekler de diğer canlılar gibi hayatta kalma ve üreme faaliyetleri için karbohidrat, protein ve lipitlere ihtiyaç duyarlar. Böcekler bu maddelerin bir kısmını çevrelerindeki besin kaynaklarından elde ederler (Nicolson, 1998; Wackers, 2007). Böcekler çevrelerinden elde ettikleri karbohidratları ya enerji elde etmek için metabolize ederler veya trehaloz ya da glikojen olarak depo ederler (Nicolson, 1998; Morgan, 2004; Klowden, 2007; Arrese ve Soulages, 2010). Karbohidratlar hemen hemen tüm canlılar için ilk başvurulan enerji ve karbon kaynaklarıdır. Karbohidratlar kolaylıkla birbirine dönüştürülebildiklerinden hayvanlar için esansiyel değildir. Böcekler amino asitleri glikoneogenezis ile karbohidratlara çevirebilmelerine rağmen yine de besinlerinde karbohidratlara gereksinim duyarlar (Ishay ve Ikan, 1968; Behmer, 2006). Alphitobius diaperinus (Coleoptera: Tenebrionidae)’da dişilerin bıraktıkları yumurta sayısının, gelişim süresinin, verimin ve yumurta büyüklüğünün karbohidrat kaynağı olarak kullanılan besin tipinden(arpa, mısır, pirinç ve beyaz un) etkilendiği belirtilmiştir (Hosen ve ark., 2004).
Stillwell ve ark. (2007), iki farklı bölgeden topladıkları C. maculatus larvalarını üç farklı baklagil çeşidinde (Vigna radiata, V. angularis, V. unguiculata) yetiştirdiğinde kullanılan besin tipine göre gelişim süresi ve erginlerin bıraktıkları yumurta sayısının değişiklik gösterdiğini, iki farklı coğrafik bölgeden toplanan populasyonları arasında bu parametrelerde farklılıklar olduğunu tespit etmişlerdir.
Genellikle böceklerin gereksinim duydukları karbohidrat, protein, lipit veya diğer maddelerin miktarları türlere ve gelişim evrelerine bağlı olarak değişebilir (Banjo ve ark., 2006). Orthoptera, Coleoptera ve Lepidoptera ordolarına ait fitofaj türler genellikle eşit miktarda protein, karbohidrat ve aminoasit gereksinimi duyarlar. Bitki floemleri ve tohumları ile beslenen tahıl zararlıları ise daha çok karbohidrata gereksinim duyarlar (Genç, 2006). Baklagil tohumları genellikle %20-25 oranında protein, %1-2 yağ ve %37-50 karbohidrat içerir (Pekşen ve Artık, 2005). Bir böcek türünün ihtiyaç duyduğu besin maddeleri büyüme, üreme, diyapoz veya göç etme davranışına göre değişir (Andersen ve ark., 1973; Singh ve Sinha, 1977; Das ve ark., 1980; Huignard, 1983; Ekbom, 1998).
Moczek (1998), Onthophagus taurus (Coleoptera: Scarabaeidae)’da besin kalitesinin ergin vücut büyüklüğünü etkilediğini, kalitesiz besinle beslenen ergin erkeklerde boynuz büyüklüğünün değişiklik gösterdiğini veya boynuz oluşumunun gözlenmediğini tespit etmiştir. Bruchidius dorsalis (Coleoptera: Bruchidae) ile yapılan çalışmada bu türün dişilerinin, iyi beslenmiş erkeklerle çiftleştiğinde, iyi beslenmemiş erkeklerle çiftleşen dişilere göre daha büyük yumurtalar bıraktıkları tespit edilmiştir (Takakura, 2004).
A.obtectus’da dişilerin oosit gelişimi için amino asitlere ihtiyaç duydukları, dişilerin oositleri için ihtiyaç duydukları bu amino asitlerden bazılarını çiftleşme sonrası spermden sağladıkları radyoaktif etiketleme çalışmaları ile tespit edilmiştir. Yapılan etiketleme çalışmaları sonucu dişilerin oositlerdeki arjinin ve histidini spermlerden sağladıkları tespit edilmiştir (Huignard, 1983).
S. oryzae ve S. granarius ile yapılan çalışmada bu türlerin larval gelişim evreleri ile erginlerindeki protein, karbohidrat ve lipit miktarlarının gelişim aşamalarına göre değişiklik gösterdiği tespit edilmiştir. Toplam karbohidrat ve lipit miktarlarının prepupa aşamasında yükseldiği ve sonra düştüğü, S. oryzae’de toplam protein miktarının prepupal evrede yükseldiği ve pupa evresinde düştükten sonra ergin evrede tekrar yükseldiği, fakat diğer türde tüm gelişim aşamalarında lineer olarak arttığı tespit edilmiştir (Singh ve Sinha, 1977).
Böcekler büyüme ve gelişmeleri için gerekli olan aminoasitleri direkt olarak besinlerinden alabilirler. Bu aminoasitler sentezlenecek yeni proteinlerin, enzimlerin, hormon reseptörlerinin, nörotransmitterlerin sentezinde kullanıldığı gibi morfogenezis,
yumurta üretimi ve yumurtaların olgunlaşması gibi faaliyetlerde de kullanılmaktadır (Genç, 2006; Renault ve ark. 2006; Benkova ve Volf, 2007; Klowden, 2007).
Kan emerek beslenen Phlebotomus papatasi (Diptera; Psycholidae) türünde besin tipinin metabolizma ve verime etkileri incelenmiştir. Kan ile beslenen dişi bireylerin, şekerle beslenen dişi bireylere oranla daha fazla yumurta bıraktığı, kan ile beslenen böceklerin konaktan emilen kandaki proteinleri yumurtadaki yedek besin maddesi proteinlerini sentezlemek için kullandıkları ifade edilmiştir (Benkova ve Volf, 2007).
Böceklerin ihtiyaç duyduğu aminoasitlerin bazıları kendileri için esansiyel aminoasitlerdir. Esansiyel aminoasitler diğer canlılarda olduğu gibi böcek türlerine göre değişir (Genç, 2006). Böceklerin kalitatif ve kantitatif amino asit ihtiyaçları değişik hayat evrelerine ve değişik oluşumlara, böceğin cinsiyetine ve o andaki fizyolojik şartlarına göre değişebilir (Chen, 1966; Fields ve ark., 1998; Bhardwaj ve Pooja, 2005). S. oryzae ve Crystolestes ferrugineus ile yapılan çalışmada düşük sıcaklıkların (-10,-20 ºC arası) bu türlerin şeker ve amino asit içeriklerine etkileri incelenmiştir. Her iki türde trehaloz, prolin, glutamik asit ve lizin miktarının soğuğa dayananlarda daha fazla, tirozinin ise daha düşük olduğu tespit edilmiştir (Fields ve ark., 1998).
Bazı aminoasitlere morfogeneziste daha çok ihtiyaç duyulurken bazıları örneğin alanin ve glisin optimal büyüme için daha önemlidir. Örneğin tirozin, kutikul dayanıklılığı için önemliyken, triptofan pigmentlerin sentezinde önemli rol oynar (Andersen ve ark., 1973; Behmer, 2006).
C. maculatus ile yapılan çalışmada bu türün erkeklerinde 20, dişilerinde ve yumurtalarında 14, larvalarında 13 ve pupalarında 17 amino asit tespit edilmiştir. Erkeklerde alanin, prolin, metiyonin, lösin, treonin ve valin, dişilerde alanin, sistin, prolin, lösin ve valin, yumurtalarda alanin, prolin ve lösin, larvalarda prolin, lösin ve valin, pupalarda ise prolin, lösin, fenilalanin ve valin aminoasitlerinin dominant olduğu bulunmuş, hiçbir gelişim evresinde izolösin tespit edilememiştir (Bhardwaj ve Pooja, 2005).
Andersen ve ark. (1973), Tenebrio molitor ile yaptıkları çalışmada ergin kutikulasında glisinin, larva ve pupa kutikulasında ise alaninin dominant aminoasitler olduğunu, pronotum ve ventral abdomenin benzer amino asitleri içerirken, dorsal abdomenin apolar aminoasitler yönünden zengin olduğunu tespit etmişlerdir. Velten ve ark. (2007), arselin amino asiti yönünden zengin olan fasulye tohumlarının, A. obtectus
dişilerinin verimini etkilediğini, bu fasulyelerle beslenen larvaların arselin içermeyen fasulyelerle beslenenlere göre gelişim sürelerinin önemli bir şekilde uzadığını, erginlerin vücut ağırlıklarının daha az olduğunu ve daha az yumurta bıraktıklarını tespit etmişlerdir.
2.3. Lipitler
Lipitler, karbohidratlar ve proteinler gibi tüm canlılarda mevcuttur ve hayatın devamlılığı için gereklidir. Genel olarak suda çözünmeyen ancak eter, benzen, kloroform gibi organik çözücülerde çözünebilen bileşikler lipit adı altında toplanır (Holme ve Peck, 1983; Gurr ve ark., 2002). Lipitler kompozisyonları ve alkalilerle olan reaksiyonları dikkate alınarak değişik şekillerde sınıflandırılabilirler. Çok genel olarak lipitler basit lipitler, kompleks lipitler ve türev lipitler olarak sınıflandırılır. Bu sınıflara yağ asitleri, gliseritler, fosfolipitler ve steroller gibi lipitler dahildir (Gurr ve ark., 2002). Basit lipitler nötral yağlar, mumlar, sterol esterleri, vitamin A, E ve K esterleridir. Kompleks lipitler ise fosfolipidler ve glikolipidlerden oluşur (Holme ve Peck, 1983).
Nötral yağlar, yağ asitlerinin gliserol ile oluşturdukları esterlerdir. Trigiliseritler, triaçilgliseroller veya açilgliseroller olarak da adlandırılır. Trigliseritler yağların ve yağ asitlerinin ana depo şeklidir (Perez-Mendoza ve ark., 1999a; Guyton ve Hall, 2001; Morgan, 2004). Gliserolün bir yağ asidi ile esterleşmesi sonucu ‘monoaçilgliseroller’, iki yağ asidi ile esterleşmesi sonucu ‘diaçilgliseroller’, üç yağ asidi ile esterleşmesi sonucunda ‘triaçilgliseroller’ meydana gelir. Gliserolün alkol grupları ile esterleşen üç yağ asidi aynı tür ise ‘basit triaçilgliseroller’, bu üç yağ asidi farklı ise ‘karma triaçilgliseroller’ şeklinde adlandırılırlar (Guyton ve Hall, 2001; Ganong, 2002; Gunstone ve ark., 2007).
Lipitlerin canlıda önemli işlevleri vardır. Bazı lipitler hem hidrofobik hemde hidrofilik kısımlar içerdiklerinden hücre zarı ve ökaryot hücrelerdeki organel zarlarının yapısında yapı elemanı olarak iş görürler (Alberts ve ark., 2002). Lipitler, vücut ağırlığını artırmadan çok fazla miktarda enerjiyi depolayabildiklerinden, enerji elde etmede metabolik yakıtın depo ve transport formu olarak iş görürler. Lipitler, karbohidratlar ve bazı böceklerde belirli amino asitler uçma için enerji kaynağı olarak görev yaparlar. Fosfolipitler ve steroller hücre zarının yapısal komponenti olarak iş görürken, larva evresinde depolanan lipitler ve böceklerde dominant form olan triaçilgliseroller besin alınmayan dönemlerde veya açlık sırasında genellikle öncelikli
enerji kaynağı olarak görev yaparlar (Lorenz ve Anand, 2004; Chamberlain ve ark., 2005; Colinet ve ark., 2006). Göç eden böceklerde öncelikli enerji kaynağı olarak daha fazla enerji verdikleri için lipitler kullanılır (Nwanze ve ark., 1976; Perez-Mendoza ve ark., 1999a; Perez-Mendoza ve ark., 1999b).
Hayvanların yağlarındaki yağ asitleri türe, gelişim evresine, çevresel şartlara, beslenme durumuna ve aktivitesine bağlı olarak miktar ve çeşit bakımından değişebilmektedir (Nwanze ve ark., 1976; Stanley-Samuelson ve ark., 1988; Parr ve ark., 1998; Morgan, 2004). C. maculatus’un uçabilen ve uçamayan normal formlarında dominant yağ asitlerinin oleik (%56) ile palmitik (%22) asit olduğu, yağ asidi çeşitlerinin benzerlik gösterdiği ve her iki formda da dişilerin daha fazla total lipit ile trigliserit içerdikleri tespit edilmiştir. Uçabilen formun normal formdan yaklaşık iki kat fazla total lipit içerdiği, uçma sırasında lipit miktarının arttığı ve yağ asidi çeşitlerinin her iki eşeyde benzer olduğu tespit edilmiştir (Nwanze ve ark., 1976). Aynı tür ile yapılan başka bir çalışmada (Parr ve ark., 1998) dişilerin bıraktıkları yumurta sayısının bitki tohumlarına (nohut ve börülce) göre değiştiği, bu tohumların oleik asit içeren karışımla muamele edilmesi halinde, dişilerin bunlara oleik asitle muamele görmemişlere göre daha fazla yumurta bıraktıkları ve oleik asidin yumurtlama davranışını uyardığı tespit edilmiştir.
Perez-Mendoza ve ark. (1999a), Rhyzoperta dominica (Coleoptera: Bostrichidae) ile yaptıkları çalışmada belirli süre (24, 48 ve 72 saat) aç bırakılan erginlerin vücut ağırlığının, lipit miktarının ve uçma davranışının azaldığını, aç bırakılan sürelerde yağ asitlerinin de azaldığını fakat çeşitlerinin değişmediğini ifade etmişlerdir. Yapılan bu çalışmada pirinç ve unla beslenen erginlerin mısır ve süpürge darısı ile beslenenlere göre daha ağır oldukları, daha fazla lipit ve oleik asit miktarına sahip oldukları ve daha çok uçma davranışında bulundukları, erginlerde tespit edilen miristik, palmitik, palmitoleik, stearik, oleik ve linoleik asitlerinin miktarlarının, beslendikleri besin tipine ve aç bırakıldıkları sürelere göre değişiklikler gösterdikleri belirtilmiştir. Ayrıca bu türde dominant yağ asitlerinin oleik, palmitik ve linoleik asit olduğunu tespit etmişlerdir.
Yağ asitleri ayrıca yağda çözünür vitaminler olan A, D, E ve K vitaminlerinin vücuda yararlı hale gelmesinde rol oynarlar (Holme ve Peck, 1983; Canavoso ve ark., 2001). Lipitlerin karbohitrat ve proteinlerle oluşturdukları glikolipitler ve lipoproteinler hücre yüzey elemanı olarak hücrelerin birbirini tanımasında, hücresel özgüllüğün ortaya
çıkmasında ve doku bağışıklığında rol oynarlar. Lipitler canlıların birçoğunun yüzeyinde koruyucu örtü olarak (Blomquist ve ark., 1987), bazı ürünlerin kanda taşınmasında taşıyıcı olarak görev yaparlar (Stanley-Samuelson ve ark., 1988).
Lipitler canlılarda birçok formda bulunabilir. Böceklerde yağ dokusu hemolenfte bulunan proteinlerin bir çoğunun sentez bölgesi ve lipitlerin %90’ını içeren trigliseritlerin ana depolanma bölgesi olarak görev yapar (Downer ve Matthews, 1976; Lorenz ve ark., 2004; Patel ve ark., 2005; Anand ve Lorenz, 2008). Yağ doku lipitlerinin çeşitleri ve miktarları böceğin gelişim evresine, eşeysel farklılığa, besine, üremeye, ergin yaşına bağlı olarak değişebilir (Mauldin ve ark., 1971; Downer ve Matthews, 1976; Guerra ve Robacker, 1989; Howard ve Stanley-Samuelson, 1990; Howard ve Stanley-Samuelson, 1996; Perez-Mendoza ve ark., 1999a,1999b; Khani ve ark., 2007).
Dişi böceklerde embriyonun gelişimi için çok önemli olan yumurta proteinleri yağ dokuda sentezlenir, hemolenfe salınır ve lipoproteinler vasıtasıyla hedef dokulara taşınır (Downer ve Matthews, 1976; Canavoso ve ark., 2001; Giron ve Casas, 2003; Lorenz ve Anand, 2004; Ziegler ve Antwerpen, 2006; Ximenes ve ark., 2008). Böcek oositlerinin ağırlığının %30-40’ını çoğunlukla triaçilgliseroller, fosfolipidler ve kolesterol oluşturur (Ziegler ve Antwerpen, 2006). C. maculusta erginlerinde (ApoLp-I ve II) ve larvalarında (ApoLp-I, II ve III) tespit edilen lipoproteinlerin total lipit miktarının larvalarda %56, erginlerde %34 olduğu, larvada en çok bulunan lipitlerin triaçilgliseroller, yağ asitleri ve diaçilgliseroller olduğu tespit edilmiş, radyoaktif olarak işaretlenen bu lipoproteinlerdeki fosfolipit ve yağ asitleri bir gün sonra C. maculatus yumurtalıklarında belirlenmiştir (Ximenes ve ark., 2008). A. grandis erginlerinin trigliseritlerindeki yağ asidi çeşit ve miktarlarının besine bağlı olarak farklılıklar gösterdiği tespit edilmiştir. Pamuk kozası ile beslenen erginlerin, şekerle beslenenlerden daha fazla miktarda oleik ve stearik, daha az miktarda linoleik ve linolenik asit içerdiği, oleik ve palmitik asidin dominant yağ asitleri olduğu, dişilerin dominant yağ asitlerini erkeklere göre daha fazla miktarda içerdiği ortaya konmuştur. Total yağ asitleri miktarının, şekerle beslenenlerde pupal deri değiştirmeden sonraki ilk dört gün çok az olduğu ve daha sonra yaş ile birlikte arttığı, pamuk kozası ile beslenenlerde ise yaşla birlikte dalgalanmalar gösterdiği bulunmuştur (Guerra ve Robacker, 1989).
Bazı yağların fiziksel ve kimyasal özelliklerini içerdikleri yağ asitlerinin kompozisyonu belirlemektedir. Yağ asidi, yapısında karboksil grubu (-COOH) taşıyan
düz bir hidrokarbon zinciri olup yağın en önemli öğesidir (Gunstone ve ark., 2007; Klowden, 2007). Doğal yağlarda bulunan yağ asitleri genelde düz zincir türevleri olup doymuş ve doymamış yağ asitleri olmak üzere iki şekilde sınıflandırılır. Doymuş yağ asitleri hidrokarbon zincirlerinde hiç çift bağ içermezler. Doymamış yağ asitleri ise hidrokarbon zincirlerinde bir veya daha fazla sayıda çift bağ içeren yağ asitleridir. Doymamış yağ asitleri hidrokarbon zincirlerinde sadece bir tane çift bağ taşıyorsa tekli doymamış yağ asitleri, birden fazla çift bağ var ise çoklu doymamış yağ asitleri olarak adlandırılırlar (Gunstone ve ark., 2007; Klowden, 2007). Doymamış yağ asitleri zarların yapısında yer alan lipitlerde önemli rol oynar. Zardaki fosfolipitlerde yer alan yağ asitleri sıcaklığa ve nem koşullarına tepki olarak hücre zarının akışkanlığını devam ettirmek için değişebilir (Chamberlain ve ark., 2005). Doğal yağlarda bulunan yağ asitleri genellikle düz zincirli ve çift sayıda karbon atomuna sahiptir (Gunstone ve ark., 2007).
Lipit sınıfı içinde ele alınan mumlar, 14-36 C atomuna sahip doymuş ve doymamış yağ asitlerinin yüksek molekül ağırlıklı monohidroksilik alkollerle yaptığı esterlerdir. Doğada yaygın olarak bazı böceklerin salgılarında, kıl ve tüylerinde bulunan mumlar, kitin oluşumunda ve su kaybının önlenmesinde önemli rol oynarlar (Downer ve Matthews, 1976; Blomquist ve ark., 1987; Stanley-Samuelson ve ark., 1988; Gunstone ve ark., 2007). Kolesterol, birçok maddenin öncül molekülü olmasının yanı sıra plazma zarı ve lipoproteinlerin temel yapı taşlarından birisi olmasından dolayı canlılarda önemli rollere sahiptir ve böceklerde bulunan başlıca steroldür (Canavoso ve ark., 2001).
Eikosanoidler, hayvanların çeşitli dokularında hormon benzeri etkileri ile bilinen ve 20 karbonlu çoklu doymamış yağ asidi olan araşidonik asit türevi bileşiklerdir (Stanley, 2006; Stanley-Samuelson ve ark., 2009). Omurgalı ve omurgasız hayvanlarda davranış, üreme ve madde taşınmasında aracı madde olarak görev yapan eikosanoidlerin son zamanlarda yapılan çalışmalarda böceklerde bakteriyel enfeksiyonlara karşı hücresel bağışıklık yanıtının oluşmasında da katkıda bulunduğu saptanmıştır (Stanley-Samuelson ve ark., 1991; Hoback ve ark., 1999; Aliza ve ark., 2001; Stanley-(Stanley-Samuelson ve ark., 2009). Serratia marcescens türü bakterisi tarafından enfekte olan Manduca sexta ve Zophobas atratus larvalarında fosfolipaz, siklooksigenaz ve lipoksigenaz enzimlerinin inhibe olduğu, buna bağlı olarak larvaların bu bakteriye karşı direnç kazanma oranının düştüğü, larvaların eikosanoidlerin sentezi için çoklu doymamış yağ