Signal detection theory analysis of category-based visual search in natural movies
Tam metin
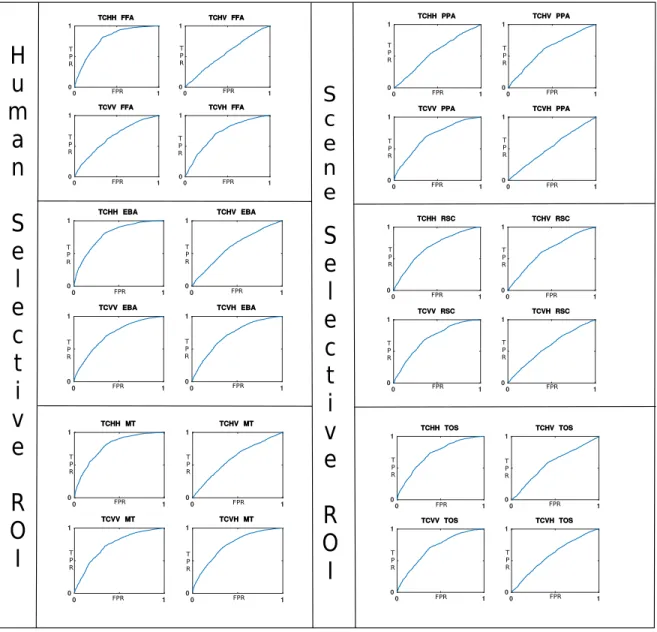
Şekil
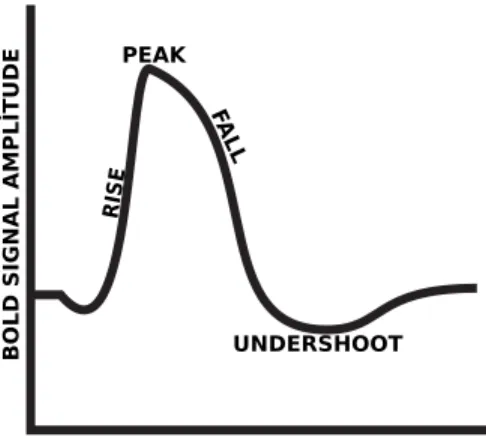
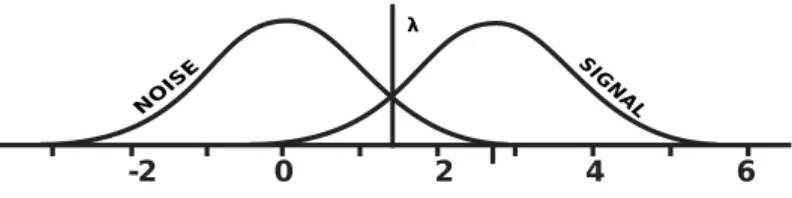
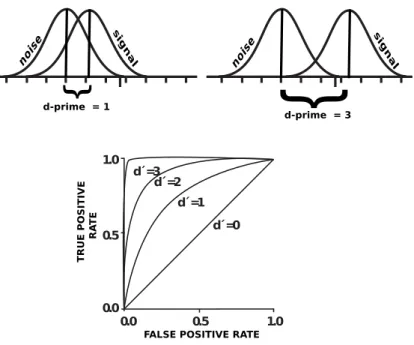
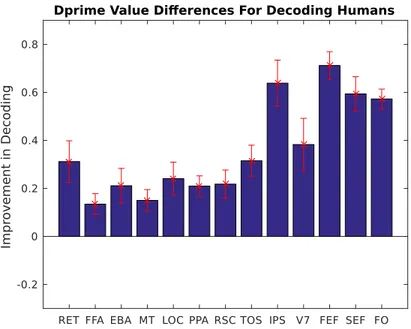
![Figure 4.3: Sigmoid function. Our modeling function is changed to estimate the binary classification [4].](https://thumb-eu.123doks.com/thumbv2/9libnet/6016537.126922/45.918.290.674.188.495/figure-sigmoid-function-modeling-function-changed-estimate-classification.webp)
Benzer Belgeler
development of the PLC program for a continuous process on the system; creation of a dataset based on the events generated; determination and reduction of the attributes; and
Detection of Toxoplasma gondii in a Eurasian Badger (Meles meles) Living in Wildlife Areas of Izmir, Turkey.. İzmir Doğal Hayatında Yaşayan bir Avrasya Porsuğunda (Meles
Temel kriz göstergeleri; cari işlemler, dış ticaret dengesi, merkez bankası rezervleri, para ikamesi, döviz kurları ve kısa vadeli sermaye hareketleri olarak
[r]
Kahramanmaraş ili merkez Dumlupınar İlkokulunda kullanılan öğretmen görüşlerine dayalı olarak seçilen, Talim ve Terbiye Kurulunun tavsiye ettiği Time For English ve Spring
In this study, the pre-test questionnaire was used to evaluate participants’ previous utilization and involvement details in Turkish e-government web pages. According to the
Next, we consider several case studies with a focus on recent experiments: the Brownian motion of a microscopic particle in thermal equilibrium with a heat bath in the presence of
Bu projede, hedeflenen tiyofen katılma ürünleri (4a-i), sentezlenen kalkon (3a-i) türevlerine, potasyum tersiyer butoksit katalizliğinde tiyofenol katılarak sentezlendi.. Bu
