T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
AĞUSTOS-2019 KONYA Her Hakkı Saklıdır BAZI BĠTKĠSEL ÖZELLĠKLERĠ BELĠRLENEN DOMUZ AYRIĞI (Dactylis
glomerata L.) GENOTĠPLERĠNDE FARKLI
SICAKLIKLARIN ÇĠMLENME ÜZERĠNE ETKĠLERĠ
Nurdan GÖKÇE YÜKSEK LĠSANS TEZĠ Tarla Bitkileri Anabilim Dalı
NURDAN GÖKÇE YÜKSEK LĠSANS TEZĠ
TEZ KABUL VE ONAYI
Nurdan GÖKÇE tarafından hazırlanan “Doğadan Toplanarak Bazı Bitkisel Özellikleri Belirlenen Domuz Ayrığı (Dactylis glomerata L.) genotiplerinde farklı sıcaklıkların çimlenme üzerine etkileri‟ adlı tez çalıĢması …/…/… tarihinde aĢağıdaki jüri tarafından oy birliği / oy çokluğu ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü ………... Anabilim Dalı‟nda YÜKSEK LĠSANS TEZĠ olarak kabul edilmiĢtir.
Jüri Üyeleri Ġmza
BaĢkan
Unvanı Adı SOYADI ………..
DanıĢman
Unvanı Adı SOYADI ………..
Üye
Unvanı Adı SOYADI ………..
Üye
Unvanı Adı SOYADI ………..
Üye
Unvanı Adı SOYADI ………..
Yukarıdaki sonucu onaylarım.
Prof. Dr. Mustafa YILMAZ FBE Müdürü
TEZ BĠLDĠRĠMĠ
Bu tezdeki bütün bilgilerin etik davranıĢ ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalıĢmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATIONPAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all material and results that are not original to this work.
Ġmza Nurdan GÖKÇE
ÖZET
YÜKSEK LĠSANS TEZĠ
DOĞADAN TOPLANARAK BAZI BĠTKĠSEL ÖZELLĠKLERĠ
BELĠRLENEN DOMUZ AYRIĞI (Dactylis glomerata L.) GENOTĠPLERĠNDE FARKLI SICAKLIKLARIN ÇĠMLENME ÜZERĠNE ETKĠLERĠ
Nurdan GÖKÇE
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Tarla Bitkileri Anabilim Dalı
DanıĢman: Doç. Dr. Mehmet Ali AVCI 2019, 42 Sayfa
Jüri
Doç. Dr. Mehmet Ali AVCI Prof. Dr. Ahmet TAMKOÇ Prof. Dr. Uğur BAġARAN
Doğal florada bol miktarda bulunan ve hayvanlar tarafından sevilerek tüketilen domuz ayrığı (Dactylis glomerata L.) ülkemizde önemli bir yere sahiptir. Kurağa dayanıklı ve gölgeye toleranslı, otlatma ve biçime oldukça dayanıklı olan ve ilkbaharda erken geliĢmeye baĢlayan domuz ayrığının meraların iyileĢtirilmesi ve yapay meraların oluĢturulmasında kullanılması tavsiye edilmektedir (Açıkgöz, 2001; Manga ve ark., 2002). Domuz ayrığı, üretimi geniĢ Ģekilde yapılan çok yıllık serin iklim bitkisidir (Lee ve ark., 2006). Domuz ayrığı yumak oluĢturarak gölge imkânı sağlamaktadır. Soğuğa ve kurağa karĢı dayanıklıdır, ayrıca su baskınlarına karĢı direnç göstermektedir. Bütün bu özellikler domuz ayrığını eĢsiz kılmaktadır (Serin ve Tan, 1998; Hannaway ve ark., 1999). Daha önce yapılmıĢ olan TÜBĠTAK projesinde kullanılan domuz ayrığı genotiplerinin büyük kısmında çimlenme sorunları görülmüĢtür. Bu nedenle bu çalıĢmada domuz ayrığı bitkisinin değiĢik sıcaklıklarda çimlenme özellikleri üzerine yapılmıĢtır. ÇalıĢma Selçuk Üniversitesi, Ziraat Fakültesi, Tarla Bitkileri Bölümü Sertifikasyon ve Kalite Laboratuvarı çimlendirme dolabında yürütülmüĢtür. ÇalıĢmada 113O919 no‟lu TÜBĠTAK projesi kapsamında seçilen 11 farklı Dactylis türünün 10 genotip ve 1 tescilli çeĢit, 4 farklı sıcaklık (5-15-25-35C⁰), 4 tekerrür ve 3 sayım (5-10-15 gün) Ģeklinde kurulmuĢtur. Gün uzunluğu 8 – 16 saat (aydınlık – karanlık) uygulanmıĢtır. Her çimlendirme kabı için sağlıklı 25‟er adet tohum kullanılmıĢtır. Bitkiler tarım perliti ortamında çimlendirilerek perlit oranı 10.8‟e ayarlanmıĢtır. Çimlenme denemesinde ilk ve son sayım günlerinde normal çim kökü ve çenek yaprağı teĢekkül eden tohumlar çimlenmiĢ kabul edilerek sayım iĢlemleri gerçekleĢtirilmiĢtir. Hiçbir Ģekilde çimlenme belirtisi görülmeyen ve anormal kök veya yaprak meydana getirilen tohumlar ise, çimlenmemiĢ olarak belirlenmiĢtir. Sonuç olarak 5-10-15 günlerinde yapılan gözlemler sonucu 5C⁰‟de 2, 15C⁰‟de 8, 25C⁰ „de 7 genotipte ayrıca her 3 farklı sıcaklık derecesinde de Doğu yıldızı çeĢidinde çimlenme görülmüĢtür. 35C⁰‟de ise hiçbir genotipte ve tescilli çeĢitte çimlenme gözlemlenmemiĢtir.
ABSTRACT
MS THESIS
THE EFFECTS OF DIFFERENT TEMPERATURES ON GERMINATION IN
Dactylis glomerata L. GENOTYPES DETERMINED BY SOME PLANT
CHARACTERISTICS
Nurdan GÖKÇE
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY
THE DEGREE OF MASTER OF SCIENCE FIELD CROPS IN AGRICULTURAL ENGINEERING
Advisor: Assoc. Dr. Mehmet Ali AVCI
2019, 42 Pages Jury
Assoc. Doç .Dr.Mehmet Ali AVCI Prof.Dr. Ahmet TAMKOÇ Prof.Dr. Uğur BAġARAN
Cocksfoot (Dactylis glomerata L.), which is abundant in natural flora and grained by animals, has a significant place in our country. It is recommended to be used in enhancement of pasture and forming artificial pasture as it is highly resistant to grazing, mowing, and drought, shade-tolerant, and also it develops in early spring(8 hours light- 16 hours dark). (Açıkgöz, 2001; Manga ve ark., 2002). Cocksfoot is a perennial cold climate plant which is produced in wide area (Lee ve ark., 2006). It provides shade region by forming grass ball. Besides of being resistant to cold and drought, it is resistant to flooding also. All these features make it unique (Serin ve Tan, 1998; Hannaway ve ark., 1999). Failures in germination were observed in most of the cocksfoot genotypes used in the previous scientific and technological research council of Turkey (TUBITAK) project. Therefore, in this study, germination properties of cocksfoot were determined at different temperatures. This study was carried out in Field Crops Department‟ Quality and Certification laboratory‟ climate cabinet of Agricultural Faculty of Selcuk University. This study was conducted with 11 different Dactylis species with different genotype and a proprietary cultivar which are selected within the scope of TUBITAK project no. 113O919 by experimenting 4 different temperature (5-15-25-35⁰C), 4 repetitions, 3 counting (5-10-15th day). Day
length was 8 - 16 hours (light - dark). 25 healthy seeds were used for each germination container. Plants were germinated in agricultural perlite environment and perlite ratio was adjusted to 10: 8. In the first and last counting days of germination experiment, normal grass roots and seed-leaf were considered germinated in counting processes. Seeds with no signs of germination or abnormal roots or leaves were identified as non-germinating. As a result, according to the observations made on 5th-10th-15th days, 2 genotypes at 5⁰C, 8 genotypes at 15⁰C, 7 genotypes at 25⁰C, and germination of Doğu Yıldızı were observed at all temperatures. No germination of any genotype and proprietary cultivar was observed at 35⁰C.
ÖNSÖZ
AraĢtırma konumu belirleyen ve her aĢamasında maddi manevi desteğini esirgemeyen danıĢman hocam Doç. Dr. Mehmet Ali AVCI, çalıĢmamızda laboratuvarı kullanmamız için yardımcı olan Prof. Dr. Ercan CEYHAN hocam, arkadaĢım Rabia KOYUNCU ve aileme teĢekkür ederim.
Nurdan GÖKÇE KONYA-2019
ĠÇĠNDEKĠLER
ÖZET ... iii
ABSTRACT ... iv
ÖNSÖZ ... v
ĠÇĠNDEKĠLER ... vi
ÇĠZELGE LĠSTESĠ ... vii
ġEKĠL LĠSTESĠ ... viii
1.GĠRĠġ... ... 1
2. KAYNAK ARAġTIRMASI ... 6
3. MATERYAL VE YÖNTEM ... 14
3.1. Materyal ... 14
3.2. Yöntem ... 15
4.ARAġTIRMA SONUÇLARI VE TARTIġMA ... 18
4.1. Genotiplerin, belirli tarihlere göre farklı sıcaklıklarda ki çimlenme bulguları ... 18
4.2. Genotiplerin 5C⁰‟ de çimlenme bulguları ... 20
4.3. Genotiplerin 15C⁰‟ de çimlenme bulguları ... 21
4.4. Genotiplerin 25C⁰‟ de çimlenme bulguları ... 23
4.5. Genotiplerin 35C⁰‟ de çimlenme bulguları ... 24
5. SONUÇLAR VE ÖNERĠLER ... 26
5.1 Sonuçlar ... 26
5.2 Öneriler ... 27
KAYNAKLAR ... 29
ÇĠZELGE LĠSTESĠ
Çizelge 3.1. Kullanılan bitkilerin lokasyonları, enlem, boylam ve yükseltileri…...13
Çizelge 4.1.1. 5,15,25 C⁰‟lerde gözlemlenen çimlenme belirtileri………...……...18
Çizelge 4.2.1. 5C⁰‟de çimlenen türlerin çıkıĢ sayısı ve çimlenme yüzdesi ... 20
Çizelge 4.3.1. 15 C⁰‟de çimlenen türlerin çıkıĢ sayısı ve çimlenme yüzdesi ... 22
Çizelge 4.4.1. 25 C⁰‟de çimlenen türlerin çıkıĢ sayısı ve çimlenme yüzdesi ... 23
ġEKĠL LĠSTESĠ
ġekil 3.1. Çimlendirme dolabı ... 15
ġekil 3.2. Ekim yapılmadan önce bir görüntü ... 16
ġekil 3.3. Ekim yapılırken görüntüsü ... 16
ġekil.3.4. Doğu Yıldızı 5 C⁰‟ de çimlenmiĢ görüntüsü………..……….17
ġekil 3.5. Doğu Yıldızı 15 C⁰ „de çimlenmiĢ yakın görüntüsü ... 17
ġekil 3.6. Doğu Yıldızı 15 C⁰ „de çimlenmiĢ uzak görüntüsü ... 17
ġekil 4.1.1. 11 genotip ve 1 tescilli çeĢit için 5-15-25-35℃ sıcaklıklarına göre çimlenme yüzdesi ... 19
ġekil 4.2.1. 5C⁰ de türlerin günlere göre uzama miktarı ... 20
ġekil 4.3.1. 15C⁰ de türlerin günlere göre uzama miktarı ... 21
ġekil 4.4.1. 25C⁰ de türlerin günlere göre uzama miktarı ... 23
1. GĠRĠġ
Ülkemizde hayvanların beslenmesi için gerek duyulan kaba yem üretimi tüketimi karĢılamamaktadır (Ayan ve ark., 2011). Yeryüzünün büyük bir bölümünü oluĢturan çayır ve mera alanlarının, dünya nüfusunun beslenmesinde en önemli kaynak olan hayvansal ürünlerin elde edilmesinde yeri büyüktür (SeydoĢoğlu ve Kökten, 2018). Ancak hayvanların kaba yem ihtiyacının karĢılandığı bu kaynakların verimleri istenilen düzeyde olmamaktadır ayrıca tarıma elveriĢli alanlarda da yem bitkileri üretimi beklenen düzeye ulaĢmamaktadır (Ayan ve ark., 2011).
Türkiye‟nin toprak yapısı ve iklimi çok farklılık göstermekte ve bu farklılık diğer ülkelerde tarımı gerçekleĢen birçok yem bitkisinin yetiĢtirilmesine olanak sunmaktadır. Diğer yandan ülkemiz bu bitkilerin birçoğunun gen merkezi konumundadır. Bu konumda olmasına rağmen, ülkemizde tarımı yapılan yem bitkileri sayısı oldukça yetersizdir. Çayır – mera alanlarının iyileĢtirilmesi daha kaliteli hale getirilmesi ve yüksek miktarda verimli kaba yem üretiminin olması için, değiĢik ekolojik bölgelere uyum sağlayan yem bitkisi tür ve çeĢitlerinin geliĢtirilmesine gerek duyulmaktadır. Diğer ülkelerden ülkemize getirilen, çeĢitler bölgelerimizin iklim ve toprak koĢullarına her zaman iyi adapte olamamakta, hastalık etmenlerinden ve birçok zararlıdan daha kolay ve fazla etkilenmeleri sebebiyle beklenen verim alınamamaktadır. Bu sebeple bölgelerimizin ekolojik Ģartlarına tamamıyla, hatta kalıtsal olarak adapte olan doğal vejetasyonda var olan popülasyonlardan faydalanarak yeni çeĢitlerin elde edilmesi, geliĢtirilmesi gerekmektedir (Ayan ve ark., 2011).
Ülkemiz TUIK (2018) verilerine göre; 14,6 milyon ha çayır mera alanımız, yaklaĢık 2 milyon ha yem bitkileri ekim alanımız vardır. Hayvansal üretimde en önemli girdiyi yem oluĢturmaktadır. Hayvan ve hayvansal ürünlerin üretimini artırmak ithalatı azaltmak için yem bitkileri ekim alanlarının ve üretiminin artırılması gerekmektedir.
Sürekli ve güvenilir kaba yem üretiminde yem bitkileri tarımı önemlidir (Akman ve ark., 2007). Yem bitkileri tarımı, bitkisel üretimde yem kaynağı sağlayarak hayvansal üretimin artmasını sağlamaktadır. Böylece hayvansal üretimin sigortası olmaktadır. Tarım alanlarında yetiĢtirilen yem bitkilerinden ilk olarak hayvanlar yem besini olarak faydalanmakta daha sonra insanlarda bu hayvanlardan süt, et, tereyağı vb.
ürünler elde ederek yararlanmaktadırlar (Soya ve ark., 2004). Yem bitkilerinin, düĢük maliyetli bitkisel ürün kaynağı olması hayvanların mide florası için ihtiyaç duyulan besin maddelerini bulundurması, vitaminler ve mineraller açısından zengin olması, hayvanların üreme potansiyelini fazlalaĢtırması ve kalitesi yüksek hayvansal kaynak sağlaması açısından hayvan beslemesinde önemli yere sahiptir (Serin ve Tan, 2001). Tarla tarımı içerisinde yer alan kaba yem bitkilerinin yetiĢtiriliciği ayrıca, çayır-mera alanlarında geliĢen fazla otlatma baskısını azaltacak, tahıl-nadas Ģeklinde ekim nöbeti uygulanan alanlarda ekim nöbetine girilmesi sağlanarak nadas alanlarında daralmasını sağlayacak bu durumda ülkemizdeki erozyon alanlarının daralmasına yardımcı olacaktır. Yem bitkisi yetiĢtirilen alanların artıĢıyla yok olmaya yüz tutmuĢ veya yok olan yem bitkisi alanlarının doğal vejetasyonlarının kendi kendini yenileme avantajını yakalamasına imkân tanıyacaklardır. Yem bitkileri yetiĢtiriciliğinde yer alan bitkiler ekim nöbetinde yer alarak kendisinden sonra yetiĢecek olan bitkilere önemli katkılarda bulunacaklardır (Soya ve ark., 2004).
Yem bitkileri ekimi için ekonomik, kaliteli, havyalar tarafından sevilerek tercih edilen yemlerin yetiĢtiriciliğini yapmalıyız. Bu kapsamda Dactylis glomerata L. (Domuz ayrığı) hayvanlar tarafından sevilerek tüketilen zengin içerikli bir bitkidir. Ülkemiz koĢullarında rahatlıkla üretilebilen bitkidir. Akdeniz iklimine adapte olmuĢ yem bitkisi olmakla birlikte domuz ayrığının Orta geçit bölgelerinde de üretimi yapılabilir.
Dactylis glomerata L. (Domuz ayrığı) beslenme değeri yüksek, kaba yapılı,9+ uzun boylu ve kalın yapraklı olmasından dolayı genellikle çayır mera ve yem bitkisi olarak yetiĢtirilmesinin yanı sıra azda olsa yeĢil alan bitkisi olarak değerlendirilmektedir (Açıkgöz, 1994).
Domuz ayrığı (Dactylis glomerata ssp. glomerata L.) doğal florada bol miktarda bulunan ülkemiz için önemi büyük serin iklim buğdaygil yem bitkisidir. Kurak ve gölgede bulunan alanlara toleranslı, otlatma ve biçime fazlasıyla dayanıklı olan ve yazdan önceki dönemde erken geliĢim gösteren domuz ayrığı meraların iyi duruma getirilmesi ve yapay meraların oluĢturulması için önerilmektedir (Açıkgöz, 2001; Manga ve ark., 2002). Ayrıca domuz ayrığı yol kenarlarında, kumsal alanlarda, deniz kıyısında, çayırlarda, az ağaçlıklı yerlerde ve yüksek rakıma sahip tepelerde kolaylıkla yetiĢebilmektedir. DeğiĢik yerlerde yetiĢmesi domuz ayrığının farklı iklim ve toprak Ģartlarına uyum kabiliyetinin fazla olduğunu göstermektedir. ġu ana kadar yapılan pek
çok çalıĢma gösteriyor ki, domuz ayrığının adaptasyon oldukça yüksek ve geliĢmiĢ ploidi düzeyine sahip olması nedeni ile hem kolay hem de geniĢ bir çalıĢma alanı sunmaktadır (Bushman ve ark., 2011)
Domuz ayrığı bitkisinde kültür çeĢitlerinin yanında yabani çeĢitlerinin de ıslah edilerek, bu bitkilerin kendiliğinden yetiĢtikleri bölgelerde, üreticilere verim ve uyum kabiliyeti yüksek olan yeni çeĢitlerin elde edilmesi yem bitkileri tarımı açısından faydalı olacaktır. Buna ek olarak domuz ayrığında dormansi ve çimlenmesi ile bağlantılı çalıĢmaların yapılması gerekmektedir. Bu çalıĢma ile ilgili yapılan bazı araĢtırmalarda domuz ayrığının 15-25 C⁰ arasında çimlendiği çimlenme gün sayımlarının 5-14 günde yapıldığı belirtilmektedir (Açıkgöz, 2001).
Çimlenme: Tohum integümentlerinin kırılarak yeni bitkinin ortaya çıkmasıyla
kendini gösteren embriyo büyümesinin yeniden baĢlama aĢamasıdır. Çimlenme integümentlerin veya tohum kabuğunun su alması ile baĢlar. Embriyo büyümesini tetikleyen enzimler yeni bitkinin toprakta yer edinebilmesi için aktif hale gelir. Domuz ayrığında (Dactylis glomerata L.) çimlenme ile ilgili çalıĢmalar yaparken temel koĢulların sağlanması gereklidir bunlar; su temini, oksijen, uygun sıcaklık, ıĢık faktörleridir. Yapılan çalıĢmada gerekli koĢullar sağlanmıĢtır.
Dormansi: Tohumlarda içsel faktörlerden kaynaklanan koĢullar uygun Ģartlar
sağlansa da fizyolojik olgunluğa ulaĢmıĢ tohumlarda çimlenme meydana gelmez. Ġçsel nedenlerden olarak tohumların dormant (uyku) halinde olduğu, uyuĢukluk içinde veya bir dinlenme safhasında olduğu söylenir. Dormant tohumlarda çimlenme için gereksinim duyulan Ģartlar yeterli düzeyde sağlansada çimlenme gerçekleĢmez. Dormant olmayan tohumlar ise çimlenme için gerekli koĢullar sağlanırsa çimlenirler. Bazı bitki türlerinin tohumlarının çimlenmesi için su yeterli faktördür. Diğer bazı türlerde sıcaklık, toprak Ģartları, ıĢık dalgalanmaları gibi ek koĢullarda önemlidir. Eğer bu faktörler olmazsa çimlenmenin olmasının önüne geçilir ve tohumlar dinlenmeye (yalancı dinlenme) zorunlu olarak girerler. Ġçsel Giberalik asit (GA) ve Absisik asit (ABA) dormansi durumunun oluĢması ile bağlantılıdır.
Ġçsel hormon eksikliği gösteren mutasyona uğramıĢ bitkilerde GA ve ABA uygulamalarının çimlenmede önemli bulgular gösterdiği belirlenmiĢtir. Dormansi etkisini yok etmek ve uygun olmayan Ģartlarda ekimi yapılan tohumların düzgün bir çimlenme ve çıkıĢ gerçekleĢtirmeleri için ekim öncesi ve hasat sonrası bazı uygulamalar
yapılmaktadır ve bantlama gibi priming olarak isimlendirilen uygulamalar sayılabilir (Hartmann ve ark., 1990; Hilhorst ve Karssen, 1992; Açıkgöz, 1994; Ercisli ve ark., 1997; Yamaguchi ve Kamiya, 2001; Demirkaya, 2006)
Doğadan toplanarak bazı bitkisel özellikleri belirlenen Domuz ayrığı (Dactylis glomerata L.)türünde dormansi görülmesi durumunda uygulanabilir bazı yöntemler Ģu Ģekildedir;
a) Osmotik Çözeltilerde (Osmopriming) ve Suda (Hidropriming) Bekletme b) Su ile Islatma- Kurutma Uygulamaları
c) Hormon Uygulanması Su ile Islatma- Kurutma Uygulamaları d) Bazı Kimyasal Maddeler ile Uygulamalar
e) DüĢük ve Yüksek Sıcaklık Uygulaması
f) Kombinasyon Uygulamaları ve Bazı Diğer Uygulamalar
Sıcaklık: Çimlenmenin farklı fazlardaki reaksiyonunu etkileyen önemli bir
çevresel faktördür.
Sıcaklık farklılıkları zar geçirgenliği, zar proteinlerinin aktivesi ve sitozol enzimleri gibi tohum çimlenmesini katalize eden birçok biyokimyasal olayı etkilemektedir. Sıcaklığın fazlalaĢması ile tohum çimlenmesindeki bu kimyasal reaksiyonların hızı yükselmektedir. Tohum çimlenmesi için minimum(düĢük), optimum(uygun), maksimum(yüksek) sıcaklıklar ve aralıkları vardır. Bu sıcaklıklar bitki tür ya da çeĢitleri için farklılık göstermektedir.
Domuz ayrığında çimlendirme çalıĢmaları Avrupa‟da da araĢtırılmıĢtır. IĢık ve alternatif sıcaklıklara dayalı yapılan çimlendirme çalıĢmalarında 12 Ġngiliz popülasyonunda da iki faktöre benzer cevaplar elde edilmiĢtir. Kullanılan 11 Avrupa popülâsyonunda cevap düzeyleri orijinal uyku hali durumuna ve görünüĢüne göre iklimsel kökene bağlıdır. Kuzey Avrupa popülâsyonları maksimum çimlenme için hem hafif hem de değiĢen sıcaklıklara ihtiyaç duyarken Akdeniz popülasyonları sabit bir sıcaklıkta ıĢıksız ortamda yüksek seviyelerde çimlenmiĢtir. Akdeniz popülâsyonlarında ılık beyaz flüoresan tüplerin ıĢığında günlük fotoperyot artıĢıyla çimlenme kapasitesi keskin Ģekilde azalmıĢtır. Avrupa popülasyonlarındaki farklı tepki örneklerinin gözlemlenmesi ise fitokrom içeren temel bir çimlenme kontrol mekanizmasındaki farklılıklarla ilgili olduğu tahmin edilmektedir (Probert ve ark., 1985a).
Domuz ayrığı serin bir buğdaygil yem bitkisidir. Akdeniz ikliminin sürdüğü yerlerde domuz ayrığına meraların verimliliğini sürdürmek ve meraların verimliğini artırmak için yazında hayatta kalması gerekir ancak yapılan çalıĢmalar bu duruma engel olduğu gözlenmiĢtir. Serin buğdaygil yem bitkilerinde yaz dormansisi görülmüĢtür. Yaz dormansisi görülen türlerde ilkbaharda çiçeklenmeden sonra yaprak büyümesinin yavaĢlatılması veya tamamen durdurulması, toprak üstü aksamın tamamen veya kısmen ölmesi ve bazen nemin kısıtlayıcı olmadığı koĢullarda bile meristemin içsel dehidrasyonu (su kaybetme), özellikle genç yaprakların tabanındaki meristemik doku içeren tomurcuklardan su kaybı gerçekleĢmektedir (Hatipoğlu ve Kökten, 2009; Norton ve Volaire, 2012). Ayrıca yaz dormansisi görülen türlerde yumru, soğan veya yaprak diplerinde tomurcuk oluĢmaktadır (Volaire ve Norton, 2006).
2. KAYNAK ARAġTIRMASI
Domuz ayrığının tanımlanması 1930‟lu yıllara dayanmaktadır. Domuz ayrığı Avrupa kıtası, Kanada, Ġngiltere‟de ıslah programları ile tanınmıĢtır. Domuz ayrığı ıslah çalıĢmaları 1953 yılında Fransa‟da bulunan INRA Versailles isimli araĢtırma enstitüsünde baĢlamıĢtır. 1962 yılına gelindiğinde çalıĢmalar Fransa‟nın Lusignan Ģehrinde devam etmiĢtir. Fransa‟daki çeĢitler kooperatif giriĢimleri sonucunda ortaya çıkmıĢtır. Avrupa, Kanada, Japonya, Yeni Zelanda gibi ülkelerde ise kamu kurumları özel Ģirketlerle iĢ birliği içerisinde domuz ayrığı yetiĢtirme programları oluĢturularak çeĢitler oluĢturulup, çoğaltılmıĢtır.
Domuz ayrığı (Dactylis glomerata L.) türü, buğdaygiller familyası (Poaceae)' nın yumak oymağı (Festuceae) içerisindeki en küçük cinslerden biridir ve 15 türü içerir. Cinsin baĢlıca türleri Acherson domuz ayrığı (Dactylis achersoniana graebn.), Ġspanyol domuz ayrığı (Dactylis hispanica griseb), Woronow domuz ayrığı (Dactylis woronowiiovez.) ve ekonomik öneme sahip en önemli türü ise adi domuz ayrığıdır (Dactylis glomerata L.). Ülkemizde ise domuz ayrığı cinsinin adi domuz ayrığı türünün 3 adet alt türünün doğal bulunduğu tespit edilmiĢtir.
Domuz ayrığının fenolojik, morfolojik ve tarımsal özellikleri konusunda yerli ve yabancı çok sayıda araĢtırma yapılmıĢtır. Domuz ayrığında ıslah çalıĢmaları daha çok ot kalitesinin artırılması üzerinde yoğunlaĢmıĢtır (Stratton ve Ohm, 1989). Çok yıllık, yumak oluĢturan bir serin mevsim buğdaygil yem bitkisidir. Avrupa, Kuzey Afrika ve Asya'nın ılıman bölgelerinin doğal bir türüdür (Whyte, 1975). Ülkemizin hemen hemen bütün bölgelerinde doğal olarak yetiĢir. Ġlkbaharda büyümeye erken baĢlar. Domuz ayrığı gölgeye, kurağa, sıcağa çayır salkım otu, kelp kuyruğu, çok yıllık çimden daha dayanıklıdır. Ancak kurağa, soğuğa toleransı kamıĢsı yumak ve kılçıksız bromdan daha azdır (Hatipoğlu ve Kökten, 2009). Zayıf ve verimi olmayan topraklarda kelp kuyruğu ve kılçıksız bromdan daha iyi yetiĢir (Hatipoğlu ve Kökten, 2009). Fakat zayıf drenajlı topraklara toleransı kamıĢsı yumaktan daha azdır. Dünya'nın nemli-ılıman bölgelerindeki birçok ülkede mera bitkisi, kuru ot, yeĢil ot ve silaj bitkisi olarak yetiĢtirilir. Serin mevsim buğdaygil yem bitkileri içerisinde en verimli türlerden biridir (Hatipoğlu ve Kökten, 2009). Domuz ayrığını diğer yem bitkilerinden ayıran en önemli
özelliklerinden biri de yaz aylarında diğer yem bitkilerinden daha fazla yem miktarına sahip olmasıdır.
Domuz ayrığı çok yıllık açık yumak oluĢturan 60-200 cm arasında boylanabilen bir yem bitkisidir. Saplarda yaprak az olmasına karĢın taban kısmı bol yapraklıdır. Yaprak ayasının enine kesiti V Ģeklindedir. Yaprak kını yassı ve omurgalıdır. Yakacık zarımsı ve 3-10 mm uzunluğundadır. Kulakçık bulunmaz. Yaprak ayası 2-12 mm geniĢliğinde ve 1 m‟ye kadar uzayabilir. Yaprak rengi açık yeĢilden koyu mavi-yeĢile kadar değiĢebilir. Çiçek durumu 8-15 mm uzunluğunda bir salkımdır. Salkımın alttaki dalları en üstteki dallardan daha uzundur ve daha fazla dallanır.
Salkımdaki baĢakçıların her birinde 3-7 lemmadan oluĢan 2-5 çiçek bulunur. Alt kısmında bir çift steril baĢak kabuğu bulunmaktadır. Çiçekçiler açarken baĢakçıklar beyaza yakın yeĢil renktedir, daha sonra aĢama Ģeklinde açık kremden kahverengiye dönüĢmektedir. BaĢağın kabuğu (lemma) mızraksı doğrusal Ģekilde omurgalı ve eĢit değildir. Bir çiftin kısa baĢak kabuğu 5-6 „mm uzunluğa sahiptir. Her baĢakçıkta üst lemmalar alt lemmalardan kısa boylu olma eğilimindedirler. Her çiçeğin bir çift beyaz tüylü stigma ve beyaz anterlere sahip 3 stamen vardır. Çiçeklenme dönemi ilkbahar sonundan, yaz ortasına kadar sürer. Bitkide büyük oranda yabancı tozlanma görülür.
Tosun (1996) tarafından doğal domuz ayrığı bitkilerinde bazı fenolojik özellikleri belirlemek amacıyla sekiz ekotip (Oltu, Ulubağ, Uzundere, Olur, Pasinler, Üniversite, Narman, Güzelyurt) kullanılarak tarla koĢullarında yapılan bir çalıĢmada ortalama sapa kalkma ve ilk çiçeklenme tarihleri sırasıyla 26 Mayıs ve 20 Haziran olarak belirtilmiĢtir. Söz konusu denemede Oltu ve Ulubağ ekotiplerinde sapa kalkma tarihleri sırasıyla 20 Mayıs-21 Mayıs, ilk çiçeklenme tarihleri ise yine sırasıyla ise 14 Haziran-17 Haziran olarak saptanmıĢtır.
Kavuzlu meyveler 4-7 mm uzunluğunda, 1-1,5 mm geniĢliğinde ve 0,2-0,8 kalınlığındadır (Gençkan, 1983). Bin tane ağırlığı 1 gramdır. Diğer buğdaygillerde olduğu gibi domuz ayrığında da saçak kök sistemi bulunmaktadadır. Kökler çayır salkım otu ve kelp kuyruğuna göre daha fazla derinlere iner. Buna karĢın köklerin ulaĢtığı toprak derinliği kılçıksız broma göre daha azdır. Bu sebepten ötürü domuz ayrığı; çayır salkım otu ve kelp kuyruğuna göre kurağa daha dayanıklı, kılçıksız broma göre daha az toleranslıdır. Bitki yedek karbonhidratlarını yaprak ayalarının tabanında, sapın toprak yüzeyine yakın kısımlarında ve köklerinde depolar. Fakat yedek karbonhidratların asıl depolandığı kısım sapların toprak yüzeyine yakın olduğu
kısımlardır. Yedek karbonhidratlar glikoz, früktoz, sakkaroz ve niĢasta halinde depolanır. Ancak depo karbonhidratların çoğunluğunu früktozlar oluĢturur.
Domuz ayrığı hayvan beslenmesinde birçok alanda kullanılmaktadır. Bunlar; mera yemi, kuru ot, silo yemi olarak yetiĢtirilmektedir (Gençkan, 1983). Bu da domuz ayrığının hayvanlar için mükemmel bir besin kaynağı olduğunu göstermektedir. Domuz ayrığının sindirimi kolay, fakat sindirimi loillum sp. kadar kolay gerçekleĢmemektedir. Ayrıca domuz ayrığı diğer yem bitkileri ile karıĢım halinde ekilebilmektedir. Adapte olduğu yerlerde fideleri yabancı otlarla ve birlikte ekildiği diğer bitkilerle iyi bir Ģekilde rekabet edebilir (Hatipoğlu ve Kökten, 2009). Vejetatif geliĢme döneminde otunun besleme değeri yoncaya yakındır. KarıĢım halinde yem bitkileri ile ekilebilecek yem bitkilerinin baĢında; ak üçgül, yonca, çayır üçgülü gibi bitkiler gelmektedir (Hatipoğlu ve Kökten, 2009).
Domuz ayrığı bitkisi oldukça kaliteli bir ot kaynağı olduğu için tüm hayvan cinsleri tarafından sevilerek otlanır. Özellikle ilkbaharda otlatmaya çok uygundur. Nöbetli otlatmaya da uygun bir bitkidir (Hatipoğlu ve Kökten, 2009). Domuz ayrığı saf olarak ot için yetiĢtirildiğinde yüksek verim için mutlaka iyi bir gübreleme gerekir. Ġlkbaharda ve her biçimden sonra 6-7 kg/da azot uygulaması ile yüksek ot verimi elde edilebilir. Biçim sayısı yetiĢme koĢullarına bağlı olarak 2-3 olabilir. Ġlk biçim salkım çıkarma baĢlangıcında yapıldığında, hem kaliteli bir ot elde edilir hem de ikinci biçimde de daha kaliteli ve yüksek ot verimi elde edilmesi mümkün olur.
Domuz ayrığında salkımlanma baĢlangıcından sonra olgunluğun ilerlemesiyle birlikte ot verimi artmasına karĢın kalite hızla düĢmektedir (Kunelius ve ark., 1974). Ġlkbaharda yüksek kaliteli ot hasadı için gerekli olan periyodun geniĢletilebilmesi amacıyla salkımlanma dönemi süresince ot kalitesindeki düĢüĢün yavaĢ olduğu domuz ayrığı genotiplerinin seçilmesi gerektiği ifade edilmiĢtir (Berg ve Hill, 1989).
Domuz ayrığı diğer birçok serin mevsim buğdaygil yem bitkisinden daha önce otlatma olgunluğuna ve biçime gelen bir bitkidir. Bu devrede bitkiler otlatılmalı veya biçilerek ot olarak değerlendirilmelidir. Otlatmada ve biçmede ortaya çıkacak gecikmeler bitkinin lezzetliliğinin düĢmesine neden olmaktadır (Aygün, 2007).
Domuz ayrığı erken otlandığında istekle otlanan, lezzetli besleyici yem niteliğindeyken geç otlatıldığında sertleĢmekte ve katılaĢmakta mera hayvanları tarafından istekle otlanamamaktadır (Gençkan, 1983).
Kuru ot verimi, çeĢide ve yetiĢme koĢullarına bağlı olarak 300-1000 kg/da arasında değiĢir. Erzurum koĢullarında saf yetiĢtirilen ve 15 kg/da azot ile gübrelenen domuz ayrığından (Serin, 1991) 492,8 kg/ da, (Koc ve ark., 1998) 963 kg/da kuru ot verimi elde etmiĢlerdir(Kökten ve ark., 2003). ġanlıurfa koĢullarında 10 kg/da azot uygulaması ile saf yetiĢtirilen domuz ayrığından 759,2 kg/da kuru ot verimi elde etmiĢlerdir Henning ve Risner (1993) ise Missouri koĢullarında saf yetiĢtirilen domuz ayrığından 300-500 kg/da kuru ot verimi elde edilebileceğini bilmektedirler. Erzurum koĢullarında(Altın, 1982), saf domuz ayrığı ekiminde 7 yılın ortalaması olarak 478 kg/da kuru ve iki yılın ortalaması olarak 78,8 kg/da ham protein verimi elde etmesine karĢılık, domuz ayrığı+ yonca karıĢımından 808 kg/ da kuru ot ve 157,5 kg/da ham protein verimi, domuz ayrığı+ çayır üçgülü karıĢımından ise 812 kg/da kuru ot ve 142,08 kg/da ham protein verimi elde etmiĢtir. Aynı koĢullarda (Koc ve ark., 1998), saf olarak yetiĢtirilen ve 15 kg/da azot ile gübrelenen saf domuz ayrığından %11.34 ham protein içeren 963 kg/da kuru ot, 1:1 oranı ile yetiĢtirilen domuz ayrığı+yonca karıĢımından %16,33 ham protein içeren 1468 kg/da kuru ot ve domuz ayrığı + çayır üçgülü karıĢımından %16,21 ham protein içeren 133,3 kg/da kuru ot verimi elde etmiĢlerdir. Kökten ve ark. (2003) ise, ġanlıurfa koĢullarında saf olarak yetiĢtirilen domuz ayrığından %9,2 ham protein içeren 759,2 kg/da kuru ot, domuz ayrığı + yonca karıĢımından %14,3 ham protein içeren 2632,1 kg/da kuru ot, domuz ayrığı+ çayır üçgülü karıĢımından %14,1 ham protein içeren 1102,5 kg/da kuru ot verimi elde etmiĢlerdir.
Gordon ve ark. (1962)domuz ayrığı bitkilerinde ham protein oranının %13,6 olduğunu tespit etmiĢlerdir. Diğer taraftan domuz ayrığı ve kılçıksız brom üzerinde yapılan baĢka bir denemede Bowman ve Law (1964)14 saatlik gün uzunluğunda ham protein oranının sırasıyla %10,66 ve %12,34 olduğu belirlemiĢtir. Aynı Ģekilde Blair ve ark. (1976) domuz ayrığı bitkilerinde ham protein oranının %13,2 olduğunu tespit etmiĢlerdir.
Beslenme değeri yüksek, kaba yapılı, uzun boylu ve kalın yapraklı olmasından dolayı genellikle çayır mera ve yem bitkisi olarak yetiĢtirilmesinin yanı sıra azda olsa yeĢil alan bitkisi olarak değerlendirilmektedir. Özellikle gölgeye dayanımı iyi olmasından ötürü gölge alanlar ile yol kenarları için uygun bir çim bitkisi olarak görülmektedir. Domuz ayrığı seyrek yapılı, çiğnenmeye dayanımı zayıf ve dipten biçime de hassas olmasından dolayı da yeĢil alan bitkisi olarak yetiĢtirilmesi ikinci
planda tutulan bir bitkidir (Açıkgöz, 1994). Bu nedenlerden ötürü yem bitkisi ıslahının yanı sıra yeĢil alan bitkisi olarak da yeni türlerin geliĢtirilmesi büyük önem arz etmektedir.
ġu ana kadar yapılan pek çok çalıĢma gösteriyor ki, domuz ayrığının adaptasyonu oldukça yüksek ve geliĢmiĢ ploidi düzeyine sahip olması nedeni ile hem kolay hem de geniĢ bir çalıĢma alanı sunmaktadır (Bushman ve ark., 2011). Ancak generatif özellikler konusunda daha az çalıĢma yapılmıĢ ve bu yönde bitkiler arasında geniĢ varyasyonlar gözlenmiĢtir (Hardison, 1984).
Domuz ayrığı (Dactylis glomerata L.) türü, Poaceae familyasının bir üyesidir ve bu familyanın özelliği temel kromozom sayısının x=7 olmasıdır (Clifford ve Dallwitz, 1985; Watson ve ark., 1986). Familya tek tür Dactylis glomerata L.(Sanada ve ark., 2010)ve en az 18 alt türü olan 2n = 6x = 42, 2n = 4x = 28 ve 2n = 2x = 14ploidi seviyelerinden oluĢur. Alt türlerin çoğu diploitlerdir (Lumaret ve Borrill, 1988). Ancak „tetraploidler‟, cinsin yaklaĢık %95'ini temsil eder ve büyük morfolojik değiĢkenlik gösterirler (Lindner ve ark., 2004).Tetraploidler, özellikle alttür glomerata da daha geniĢ bir coğrafi aralığı kaplar. Sensulatu‟nun kromozomlarının çiftleĢmesi doğrultusunda 2x soy‟dan ortaya çıkmıĢlardır (Lumaret ve Borrill, 1988). Hem iklim değiĢiklikleri hem de çevresel faktörler bu geliĢmeye neden olmuĢtur. ÇalıĢmalar, hem doğal (Zohary, 1959; Jones ve Borrill, 1962; Lumaret ve Hanotte, 1987; Lumaret ve Barrientos, 1990; Sahuquillo ve Lumaret, 1999) hem de yapay ortamlarda (Horn ve ark., 1988)2x ve 4x arasında melezleĢmenin gerçekleĢtiğini göstermiĢtir. Dactylis ıslah programlarında modern biyoteknoloji yöntemlerinin uygulanması için potansiyel bir bitkidir (Charmet ve ark., 1997; Forster ve ark., 2008).
Bitkisel üretimde olduğu gibi domuz ayrığında da ilk aĢama çimlenmeyle baĢlar (Karakurt ve ark., 2010). Çimlenme; tohum integümentlerinin kırılması ve yeni bitkinin ortaya çıkması ile kendini gösteren embriyo büyümesinin yeniden baĢlama safhasıdır. Çimlenme integümentlerin veya tohum kabuğunun su alması ile baĢlar. Embriyo büyümesini uyaran enzimler yeni bitkinin toprakta yerleĢebilmesi için aktif hale gelir. Domuz ayrığında (Dactylis glomerata L.)çimlenme ile ilgili çalıĢmalar yaparken temel koĢulların sağlanması gereklidir, bunlar; su temini, oksijen, uygun sıcaklık, ıĢık faktörleridir.
Su: Tohum çimlenmesini baĢlatan oluĢan bitkilerin hayatını sürdürebilmesi için
en önemli etmenlerdendir. Toprakta var olan potansiyel tuzların bulunması suya bağlıdır. Yüksek oranda tuz çimlenme ortamını olumsuz yönde etkilemektedir. Bazı tohumlarda çimlenmeye engel olan maddelerin bulunması bazı madde ile kaplı olmaları nedeni ile yıkanmaya ihtiyaç duymaktadırlar (Hartmann ve ark., 1990).
Sıcaklık: Çimlenme zamanını ayarlayan önemli faktörlerden biridir.
Dormansinin denetlenmesinde direkt olarak bağlantılıdır. DüĢük sıcaklıklarda çimlenme çoğunlukla düĢüktür. Akdeniz iklimine sahip bölgelerde bitkilerin tohumu en yüksek (optimum) 24-30 C⁰‟de çimlenirken 4,5-40C⁰ aralığında geniĢ olarak çimlenebilme kabiliyetine sahiptirler. Akdeniz iklimine sahip yerlerde ayrıca düĢük sıcaklıklarda (3-4C⁰) kromozom sayılarının iki katına çıkarılması gerekmektedir (Hartmann ve ark., 1990).
Oksijen: Çimlenme gerçekleĢirken embriyo ve çimlenme ortamı arasındaki gaz
alıĢveriĢinin hızlı ve değiĢmez olarak gerçekleĢmesi önemlidir. Çimlenen tohumlar solunumunu oksijen sayesinde gerçekleĢtirmektedirler. Bitkilerde özümleme faaliyetleri artıkça oksijen alımında da artıĢ gerçekleĢmektedir. Eğer ortamda fazla oranda su var ise oksijen birikimi engellenmektedir (Hartmann ve ark., 1990).
IĢık: Bazı bitkilerde ıĢık dormansiyi baĢlatırken, bazılarında ise dormansi
etkisini ortadan kaldırmaktadır. Tohumlarda ıĢığa verilen tepkinin temel mekanizması kimyasal olarak aktif olarak yararlanılan fitokron pigmenti ile bağlantılı olması yapılan çalıĢmalar neticesinde tespit edilmiĢtir. Kırmızı ve kızılötesi ıĢınlar bazı tohumlarda GA biyosentezi etki gösterdiği saptanmĢtır (Georghiou ve ark., 1982; Yamaguchi ve Kamiya, 2001). Bazı tohumlar kırmızı ıĢık etkisi altında bırakılarak bu tohumların çimlenmeye olan etkisi gözlemlenmiĢ bu gözlem sonucunda ise çimlenmede artıĢ olduğu görülmüĢtür. Bazı tohumlar da kızıl ötesi ıĢık verilerek gözlem yapılmıĢ ancak çimlenmenin bu ıĢıkta olumsuz olduğu görüldü. IĢığın etkisi tohum kabuğu ve embriyo olduğunda duyarlılığın olduğu bunların uzaklaĢtırıldığında ise ıĢığın etkisinin yok olduğu gözlemlenmiĢtir (Hartmann ve ark., 1990).
Sitokininler; hücrenin geniĢlemesi, bölünmesi, uyarılması, uzun süre genç kalmasını sağlama, tuber oluĢumunda görev alır. Oksinler ise hücre bölünmesi, büyümesi, kök oluĢumuna, apikal dominansiye neden olmaktadır (Ünyayar ve Topçuoğlu, 1998). Oksin ve sitokininlerin görevi içsel hormon, fizyolojik durum arasındaki bağlantı ile sınırlandırılmıĢtır. Tohum çimlenmesinde sitokinler ABA etkisini
engeller ya da ortadan kaldırarak çimlenmenin olumlu yönde geliĢmesini sağlar (Hartmann ve ark., 1990).
Etilenin çimlenmede uyarıcı bir faktör olduğu son çalıĢmalarda gözlemlenmiĢtir (Hartmann ve ark., 1990). Akdeniz iklimine sahip bölgelerde meyve türlerinde çimlenmeyi artırmak için hormon uygulaması yapılırken tohum katlaması iĢlemine yer verilmemektedir.
Gibberalinler geliĢmiĢ tohumlarda yüksek miktarda bulunmaktadır. Bu da dormansi ve tohum çimlenmesi kontrolünde önemli bir faktördür.GA3etkileri çimlenme
üzerine etkisi büyüktür, bu konuyla ilgili yapılan çalıĢmalar bunu kanıtlamaktadır (Duman, 2006).
Çimlenme oluĢurken olumsuz Ģartların yaĢanması çimlenmeyi kötü yönde etkilemektedir. Bu olumsuz Ģartlar (toprak sıcaklığının düĢük olması, toprağın kaymak tabakasına sahip olması gibi) tohumdan dolayı da tohumun sahip olduğu yapısal özelliklerinden dolayı çimlenme olumsuz gerçekleĢmektedir. Dormansi ve çimlenme arasındaki bağlantıyı saptamak için çok sayıda çalıĢma yapılmıĢtır. Bitkilerin genelinde oksijen, nem, sıcaklık, ıĢık gibi çevre faktörleri yeterli olsa da çimlenme gerçekleĢmeyebilir, bu olaya dormansi denilmektedir. Ilıman bölgelerde bulunan bitkilerin tohumlarında çoğunlukla dormansi ile karĢılaĢılmaktadır (ÇetinbaĢ ve Koyuncu, 2005; Demirkaya, 2006). Embriyoda oluĢan dormansi ile dıĢ kabuktan dolayı oluĢan dormansi aynı değildir. Embriyoda dıĢtaki katmanların uzaklaĢtırılması dormansi etkisini ortadan kaldırmaktadır. Embriyonun geliĢme gücü dıĢ kabuktaki mekanik sınırlamadan kuvvetliyse çimlenme baĢlar (Karakurt ve ark., 2010).
Dormansiden etkilenen tohumlar çimlenme için gerekli koĢullar yeterli olmadığından dolayı çimlenmez. Dormansiden etkilenmeyen tohumlar ise gerekli koĢullar sağlandığında çimlenirler. Bitki türlerinin çok azında su yeterlidir. Birçok bitki türünde ise toprak koĢulları, ıĢık, sıcaklık dalgalanmaları gibi ek faktörlerde gereklidir. Eğer bu gereklilikler olmazsa çimlenme olmaz, tohumlar dinlenme (yalancı dinlenme) durumuna geçerler. Dormansi bazı faktörlerle iliĢkilidir. Bu faktörler içsel Giberalik asit (GA) ve Absisik asit ile iliĢkilidir. Mutasyona uğramıĢ bitkilerde içsel hormon eksikliği gösterir. Bu bitkilere GA ve ABA uygulanarak çimlenmede önemli tesirler gösterdiği saptanmıĢtır. Dormansi etkisini yok etmek ve uygunsuz Ģartlarda ekilen tohumların düzgün çıkıĢ ve çimlenme sağlayabilmeleri için ekimden önce ve hasattan sonra bazı uygulamalar yapılmaktadır. Bu uygulamalardan bazıları tohumların kaplanması,
ıslatılması, büyüme düzenleyicilerin kullanılması, asitlerle aĢındırması, osmatik çözeltilerde tutulması, çimlendikten sonra jel halinde ekiminin yapılması, bantlama gibi priming olarak isimlendirilen yöntemler kullanılabilir (Hartmann ve ark., 1990; Hilhorst ve Karssen, 1992; Karakurt ve ark., 2010).
3. MATERYAL VE YÖNTEM
3.1. Materyal
Bu araĢtırmada materyaller, Doç. Dr. Mehmet Ali AVCI tarafından yürütülen TÜBĠTAK 113O919 no‟lu “Doğal Florada Bulunan Çim ve Yem Olarak Kullanılabilecek Bazı Buğdaygil Yem Bitkilerinin Toplanması ve Islah Amaçlı Kullanılması” adlı proje kapsamında, 2014 – 2015 yıllarının Nisan – Temmuz ayları arasında toplanan (Ankara, Çankırı, Çorum, Yozgat, EskiĢehir, Afyon, Konya, Aksaray, Niğde, Karaman, KırĢehir, Kayseri, Kırıkkale, Sivas, Mersin ve Antalya) 11 adet Dactylis glomerata L. türüne ait genotiplerdir. Ayrıca Erzurum- Doğu Anadolu Tarımsal AraĢtırma Enstitüsünde geliĢtirilen 1 adet tescilli domuz ayrığı çeĢidi (Doğu Yıldızı) çalıĢmada kullanılmıĢtır. ÇalıĢmada kullanılan 12 adet Dactylis glomerata L. genotiplerinin bitki numarası, lokalitesi, enlem, boylam ve yükseltisi çizelge 3.1.‟ de verilmiĢtir.
Çizelge3.1.Kullanılan bitkilerin lokasyonları, enlem, boylam ve yükseltileri
Bitki numarası (Alınan yer) Lokalite Enlem Boylam Yükselti
(m) 1. (61-8) EskiĢehir Alpullu Mihalıççık 39⁰49.776K 031⁰12.510 D 986
2.(60-15) Eber Gölü AkĢehir 38⁰36.577K 031⁰07.646 D 984
3.(2016-12-2) Çorum 40⁰34.745K 035⁰01.948 D 1028
4.(43-12) TaĢkent Alanya Sapağı 36⁰51.589K 032⁰31.228 D 1734
5.(25-2) Cuma alanlı Köyü –Denizli 37⁰48.201K 029⁰09.112 D 1460 6.(25-1) Denizli Cumali Köy1ü Kırkbudak
Geçidi 37⁰46.322K 029⁰06.318 D 1500
7.(49-5) Sivas Mescitli 39⁰39.541K 037⁰00.607 D 1296
8.(2016-12-8) Çorum Ġskilip 40⁰34.897K 034⁰46.959 D 1026
9.(2016-11-8) Akçatekir Eski Kanacık Köyü
Mezarlığı 37⁰23.776K 034⁰50.670 D 939
10.(2016-42-7) Konya Yaylaları 37⁰52.431K 032⁰18.199 D 1302
11.(2016-15-41-3) Konya Sefa köy (Eski Yol) 37⁰46.569K 032⁰13.394 D 1320
3.2. Yöntem
ÇalıĢma, Doç. Dr. Mehmet Ali AVCI tarafından yürütülen TÜBĠTAK projesinden bazı bitkisel özellikleri belirlenmiĢ bitkilerden seçilen Dactylis glomerata L. genotiplerinden elde edilmiĢ tohumlar ile çalıĢma Selçuk Üniversitesi Ziraat Fakültesi Tarla Bitkiler Bölümü Sertifikasyon ve Kalite Laboratuvarı çimlendirme dolabında yürütülmüĢtür Deneme 11genotip ve 1 tescilli çeĢit, 4 farklı sıcaklık (5-15-25-35C⁰), 4 tekerrür ve 3 sayım (5-10-15 gün) Ģeklinde kurulmuĢtur. Gün uzunluğu 8 – 16 saat (aydınlık – karanlık) uygulanmıĢtır. Her çimlendirme kabı için sağlıklı 25‟er adet tohum kullanılmıĢtır. Bitkiler tarım perliti ortamında çimlendirilerek perlit oranı 10:8‟e ayarlanmıĢtır. Çimlenme denemesinde ilk ve son sayım günlerinde normal çim kökü bulunan tohumlar çimlenmiĢ kabul edilerek sayım iĢlemleri gerçekleĢtirilmiĢtir. Hiçbir Ģekilde çimlenme belirtisi görülmeyen veya anormal kök veya yaprak meydana getirilen tohumlar ise, çimlenmemiĢ olarak belirlenmiĢtir.
ġekil 3.1. Çimlendirme dolabı
ġekil 3.2. Ekim yapılmadan önce bir görüntü
ġekil 3.4. Doğu Yıldızı 5 C⁰‟ de çimlenmiĢ görüntüsü
ġekil 3.5. Doğu Yıldızı 15 C⁰ „de çimlenmiĢ yakın görüntüsü
4.ARAġTIRMA SONUÇLARI VE TARTIġMA
4.1. Genotiplerin, belirli tarihlere göre farklı sıcaklıklarda ki çimlenme bulguları
Çizelge 4.1.1 5,15,25 C⁰‟lerde gözlemlenen çimlenme belirtileri.
Yukarıda tabloda, 5 gün arayla yapılan ölçümlere göre farklı sıcaklıklarda ki çimlenme bulguları yer almaktadır. Verilere göre, Doğu yıldızı ve 61-8 genotipleri ilk 5
BĠTKĠLER 0 5 .0 6 .2 018 5⁰C 05 .0 6 .2 0 1 0 .0 6 .2 018 5⁰C 15 .0 6 .2 018 15C ⁰ 2 6 .0 6 .2 018 15 ⁰C 0 1 .0 7 .2 0 18 25C 0 0 6 .0 7 .2 018 25C 0 1 0 .g u n 1 1 .0 7 .2 0 18 25 ⁰C Doğu Yıldızı Çimlenme var Çimlenme var Çimlenme var Çimlenme var Çimlenme var Çimlenm e var Çimlenm e var 61-8 Çimlenme var Çimlenme var Çimlenme var Çimlenme var Çimlenme var Çimlenm e var Çimlenm e var 60-15 Çimlenme var Çimlenme var Çimlenme var Çimlenme var Çimlenme yok Çimlenm e var Çimlenm e var 2016-12-2 Çimlenme yok Çimlenme yok Çimlenme yok Ekim
yapıldı Çimlenme yok
Çimlenm e yok Çimlenm e yok 43-12 Çimlenme yok Çimlenme var Çimlenme var Çimlenme geliĢimi yok Çimlenme yok Çimlenm e var Çimlenm e yok 25-1 Çimlenme yok Bazı kutularda çimlenme var Çimlenme yok Çimlenme yok Çimlenme yok Çimlenm e var Çimlenm e var 25-2 Çimlenme yok Çimlenme yok Çimlenme yok Çimlenme yok Çimlenme yok Çimlenm e yok Çimlenm e var 49-5 Çimlenme yok Bazı kutularda çimlenme var Çimlenme var DeğiĢim yok DeğiĢim yok ÇıkıĢ yok Çimlenm e yok 2016-12-8 Çimlenme yok Çimlenme var Çimlenme var Çimlenme var DeğiĢim yok Tohumda çürüme var Tohumda çürüme var 2016-11-8 Çimlenme yok Çimlenme yok Çimlenme yok DeğiĢim yok DeğiĢim yok Tohumda çürüme var Tohumda çürüme var 2016-42-7 Çimlenme yok Çimlenme yok Çimlenme yok Çimlenme yok DeğiĢim yok Çimlenm e var Çimlenm e var 2016-15-41-3 Çimlenme yok Çimlenme yok Çimlenme yok Çimlenme yok Çimlenme yok Çimlenm e yok Tohumlar cansız
gün içinde dormant halinden çıkarak çimlenme belirtileri göstermiĢler; aynı zamanda her sıcaklıkta verime geçmiĢlerdir. 2016-11-8 genotipi 4 farklı sıcaklıkta da herhangi bir çimlenme belirtisi göstermemiĢ ve tohumlar 25C⁰‟ de çürümeye baĢlamıĢtır. Geriye kalan genotipler de ise diğer sıcaklıklara oranla 25C⁰‟ de daha fazla çimlenme bulgusuna rastlanmaktadır. Genotiplerin sıcaklıklara göre çıkıĢ yüzdesi aĢağıdaki tablolarda detaylı bir Ģekilde verilmiĢtir.
ġekil 4.1.1. 11 genotip ve 1 tescilli çeĢit için 5-15-25-35℃ sıcaklıklarına göre çimlenme yüzdesi
11 genotip ve 1 adet tescilli çeĢit için 5-15-25-35 C⁰ sıcaklıklarına göre çimlenme yüzdesi grafiği yukarıdaki Ģekilde görülmektedir. Bu grafiğe göre 25⁰C birçok genotipin çimlenmesi için en uygun aralık ve 1.(61-8) genotipi için optimum sıcaklıktır. 12.(Doğu Yıldızı) için ise 15⁰C en uygun çimlenme sıcaklığı olarak gözlemlenmiĢtir. 35⁰C‟de hiçbir çimlenme bulgusuna rastlanmamıĢtır. 5⁰C‟ de ise verim oranı düĢükte olsa 3 farklı genotipte çimlenme gözlemlenmiĢtir.
0 10 20 30 40 50 60 0 5 10 15 20 25 30 Ç im len m e Yü zd esi % Sıcaklık ⁰C
Türlerin Sıcaklığa Göre Çimlenme Yüzdesi
12 2 1 3 8 11 5 6 7 4
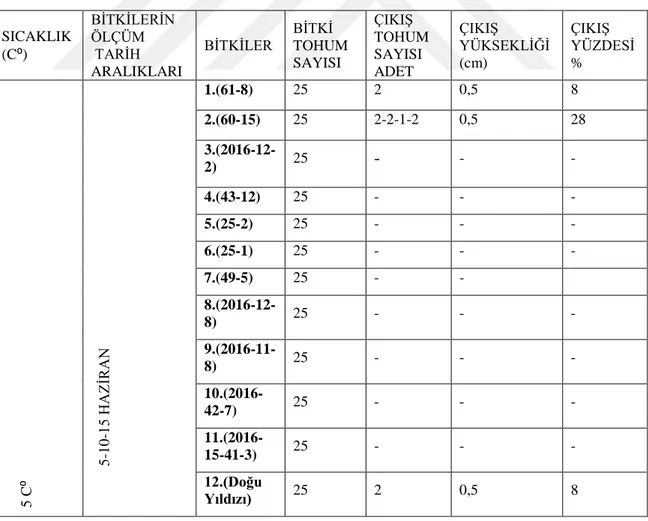
4.2. Genotiplerin 5C⁰’ de çimlenme bulguları
ġekil 4.2.1. 5C⁰ de türlerin günlere göre uzama miktarı
Çizelge 4.2.1. 5C⁰‟de çimlenen türlerin çimlenme çıkıĢ sayısı ve çimlenme yüzdesi 0 0,2 0,4 0,6 0,8 1 1,2 0 2 4 6 8 10 12 14 16 Uza m a m ik tar ı ( cm ) Zaman (gün)
5⁰C' deTürlerin Günlere Göre Uzama Miktarı
12 2 1 SICAKLIK (C⁰) BĠTKĠLERĠN ÖLÇÜM TARĠH ARALIKLARI BĠTKĠLER BĠTKĠ TOHUM SAYISI ÇIKIġ TOHUM SAYISI ADET ÇIKIġ YÜKSEKLĠĞĠ (cm) ÇIKIġ YÜZDESĠ % 5 C ⁰ 5 -10 -1 5 HAZ ĠR AN 1.(61-8) 25 2 0,5 8 2.(60-15) 25 2-2-1-2 0,5 28 3.(2016-12-2) 25 - - - 4.(43-12) 25 - - - 5.(25-2) 25 - - - 6.(25-1) 25 - - - 7.(49-5) 25 - - 8.(2016-12-8) 25 - - - 9.(2016-11-8) 25 - - - 10.(2016-42-7) 25 - - - 11.(2016-15-41-3) 25 - - - 12.(Doğu Yıldızı) 25 2 0,5 8
5C⁰‟de 8 çeĢitte çimlenme gözlemlenmemiĢtir. Diğer 2 çeĢitte 1.(61-8), 2.(60-15) ve 12.(Doğu yıldızı) tescilli çeĢidinde 5-10-15. günlerde yapılan gözlem sonucu çimlenme gözlemlenmiĢtir. Bu genotipler arasında 2.(60-15), % 28 ile en fazla çimlenme yüzdesine sahiptir. Çimlenen 2 genotip ve Doğu yıldızı çeĢidi için 10. günde çimlenmenin eĢit miktarda baĢladığı 15. günde ise Doğu yıldızının 1cm‟e ulaĢtığı yukarıdaki grafikte de olduğu gibi görülmektedir. 5C⁰‟de gerçekleĢtirilen çalıĢmanın gözlemleri sonucunda bu sıcaklığın 11 çeĢitten 2‟sinin ve tescilli çeĢidin çimlenmesi halinde bu sıcaklığın ekimi yapılan tohumlar için ideal bir sıcaklık olmadığını göstermiĢtir. 5C⁰‟nin Doğu Yıldızı, 1.(61-8), 2.(60-15) çeĢitleri için ise çimlenmeyi baĢlatmak için uygun sıcaklık olabileceği fakat fidelerin en fazla 1cm‟ e ulaĢması durumundan dolayı geliĢim göstermesi için yeterli sıcaklık olmadığı gözlemlenmiĢtir.
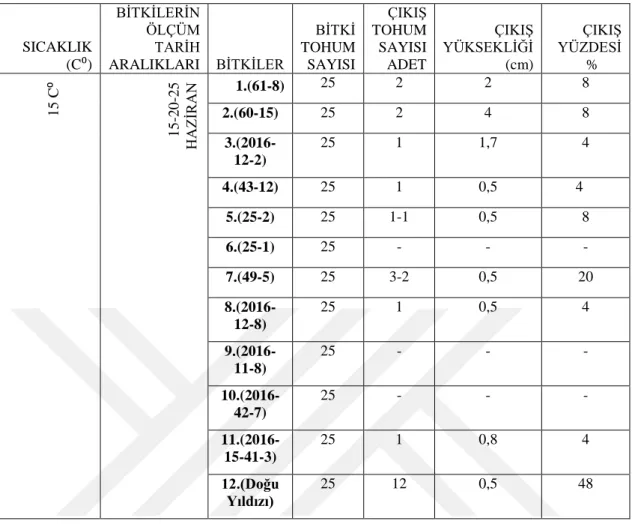
4.3. Genotiplerin 15C⁰’ de çimlenme bulguları
ġekil 4.3.1. 15C⁰ de türlerin günlere göre uzama miktarı 0 1 2 3 4 5 6 7 8 0 2 4 6 8 10 12 14 16 Uza m a m ik tar ı ( cm ) Zaman (gün)
15⁰C' deTürlerin Günlere Göre Uzama Miktarı
12 7 1 2 4 5 11 8 3
Çizelge 4.3.1. 15 C⁰‟de çimlenen türlerin çimlenme çıkıĢ sayısı ve çimlenme yüzdesi SICAKLIK (C⁰) BĠTKĠLERĠN ÖLÇÜM TARĠH ARALIKLARI BĠTKĠLER BĠTKĠ TOHUM SAYISI ÇIKIġ TOHUM SAYISI ADET ÇIKIġ YÜKSEKLĠĞĠ (cm) ÇIKIġ YÜZDESĠ % 1 5 C ⁰ 15 -20 -25 HAZ ĠR AN 1.(61-8) 25 2 2 8 2.(60-15) 25 2 4 8 3.(2016-12-2) 25 1 1,7 4 4.(43-12) 25 1 0,5 4 5.(25-2) 25 1-1 0,5 8 6.(25-1) 25 - - - 7.(49-5) 25 3-2 0,5 20 8.(2016-12-8) 25 1 0,5 4 9.(2016-11-8) 25 - - - 10.(2016-42-7) 25 - - - 11.(2016-15-41-3) 25 1 0,8 4 12.(Doğu Yıldızı) 25 12 0,5 48
15C⁰‟ de çimlenen çeĢitlerin sayısında artıĢ gözlemlenmiĢtir. 8 genotipte ve Doğu Yıldızı çeĢidinde çimlenme görülmüĢtür. 6.(25-1), 9.(2016-11-8), 10.(2016-42-7) çeĢitlerinde ise çimlenme gözlemlenmemiĢtir. Çimlenen tohumların ilk çimlenme yüksekliği genelde 0,5 cm olmuĢtur. En uzun çimlenme yüksekliğine ve en yüksek çimlenme yüzdesine ulaĢan ise Doğu Yıldızı tescilli çeĢidi olmuĢtur. En yüksek ölçüm uzunluğu 7 cm, çimlenme yüzdesi ise %48 olarak bulunmuĢtur. Doğu Yıldızından sonra en yüksek uzunluğa sahip olan 5.(25-2) çimlenme yüzdesi bakımından oldukça düĢüktür. Fakat 7.(49-5) genotipi Doğu Yıldızından sonra en yüksek çimlenme yüzdesine sahiptir ve boyu ölçümler sonucunda maksimum 3,5 cm‟e ulaĢmıĢtır.
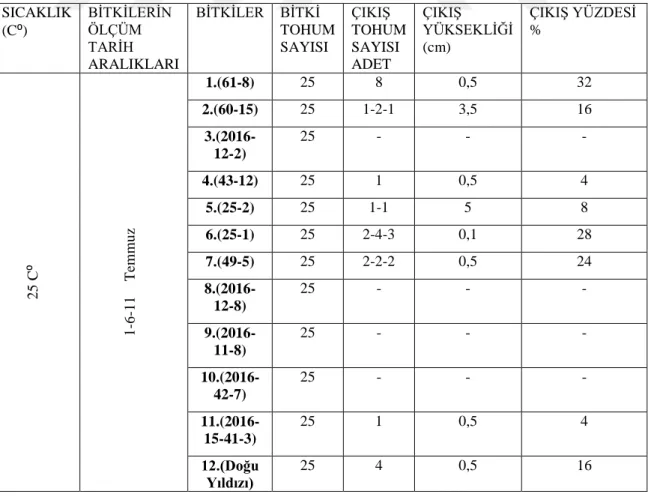
4.4. Genotiplerin 25C⁰’ de çimlenme bulguları
ġekil 4.4.1. 25C⁰ de türlerin günlere göre uzama miktarı
Çizelge 4.4.1. 25 C⁰‟de çimlenen türlerin çimlenme çıkıĢ sayısı ve çimlenme yüzdesi 0 1 2 3 4 5 6 7 8 0 2 4 6 8 10 12 14 16 Uza m a m ik tar ı ( cm ) Zaman (gün)
25⁰C' deTürlerin Günlere Göre Uzama Miktarı
12 7 1 2 4 5 6 11 SICAKLIK (C⁰) BĠTKĠLERĠN ÖLÇÜM TARĠH ARALIKLARI BĠTKĠLER BĠTKĠ TOHUM SAYISI ÇIKIġ TOHUM SAYISI ADET ÇIKIġ YÜKSEKLĠĞĠ (cm) ÇIKIġ YÜZDESĠ % 2 5 C ⁰ 1 -6 -1 1 T em m u z 1.(61-8) 25 8 0,5 32 2.(60-15) 25 1-2-1 3,5 16 3.(2016-12-2) 25 - - - 4.(43-12) 25 1 0,5 4 5.(25-2) 25 1-1 5 8 6.(25-1) 25 2-4-3 0,1 28 7.(49-5) 25 2-2-2 0,5 24 8.(2016-12-8) 25 - - - 9.(2016-11-8) 25 - - - 10.(2016-42-7) 25 - - - 11.(2016-15-41-3) 25 1 0,5 4 12.(Doğu Yıldızı) 25 4 0,5 16
25C⁰‟de 2016-11-8, 2016-12-2, 2016-12-8, 2016-42-7 genotiplerinde çimlenme gözlemlenmezken diğer 7 genotipte ve 1 adet tescilli çeĢitte çimlenme gözlemlenmiĢtir. 25-1 genotipinde daha düĢük sıcaklarda çimlenme görülmezken 25C⁰‟de çimlenme görülmüĢtür. Bu sıcaklıkta en yüksek boy uzunluğu ölçülen 7,5 cm ile Doğu Yıldızı ardından 6 cm ile 1.(61-8) genotipi olmuĢtur. En yüksek çimlenme yüzdesine sahip olan ise %32 ile 61-8 numaralı genotip olmuĢtur. 1.(61-8) genotipinin çimlenme yüzdesi 15⁰C‟ye göre 4 kat artarken Doğu Yıldızının çimlenme yüzdesi 3 kat azalmıĢ ve %48‟den %16‟lara kadar gerilemiĢtir.
4.5. Genotiplerin 35C⁰’ de çimlenme bulguları
Çizelge 4.5.1. 35 C⁰‟de çimlenen türlerin çimlenme çıkıĢ sayısı ve çimlenme yüzdesi SICAKLIK C⁰ BĠTKĠLERĠN ÖLÇÜM TARĠH ARALIKLARI BĠTKĠLER ÇIKIġ TOHUM SAYISI ADET ÇIKIġ YÜKSEKLĠĞĠ (CM) ÇIKIġ YÜZDESĠ % 3 5 C ⁰ 15 -20 -2 5 T em m u z 1.(61-8) 25 - - 2.(60-15) 25 - - 3.(2016-12-2) 25 - - 4.(43-12) 25 - - 5.(25-2) 25 - - 6.(25-1) 25 - - 7.(49-5) 25 - - 8.(2016-12-8) 25 - - 9.(2016-11-8) 25 - - 10.(2016-42-7) 25 - - 11.(2016-15-41-3) 25 - - 12.(Doğu Yıldızı) 25 - -
Yapılan çalıĢmada 35C⁰‟de çimlenme 10 genotip ve 1 tescilli çeĢitte gözlemlenmemiĢtir. Yapılan çalıĢma koĢullarına göre 35 Cº „de çimlenmenin olmaması bu sıcaklığın, genotiplerden optimum bir verim almak için uygun olmadığını göstermektedir.
5. SONUÇLAR VE ÖNERĠLER
5.1 Sonuçlar
Bu çalıĢmada 1130919 no‟lu TÜBĠTAK projesi kapsamında seçilen 12 farklı Dactylis türü 11 genotip ve 1 tescilli çeĢit olan Doğu Yıldızı kullanılmıĢtır. Bu türler laboratuvar koĢullarında 4 farklı sıcaklık altında (5-15-25-35) C⁰ çimlendirme kabininde çimlenmeleri gözlemlenmiĢtir. Gözlemlerde her sıcaklıkta tekrarlanmak üzere (5-10-15) günde bir olmak üzere çimlendirme sayımları yapılmıĢtır.
Çimlendirmede sayım iĢlemi gerçekleĢirken yapılan gözlemlerde dikkate alınan faktörler tohum çıkıĢ sayısı, çimlenmenin var ya da yok olması, tohumda renk değiĢikliği, tohumda çürüklüğü olmuĢtur. ÇalıĢma 2018 yılının mayıs ayının son haftasında baĢlayıp, temmuz ayının ikinci haftasında sona ermiĢtir. 4 farklı sıcaklıkta (5-15-25-35) C⁰ farklı gözlemler elde edilmiĢtir, bu gözlemler sırasıyla;
5 C⁰‟de gözlemlenen değiĢimler: Doğu Yıldızı tescilli çeĢit ve 61-8 genotipli dactylis türünde değiĢim gözlemlenmiĢtir, çimlenme gerçekleĢmiĢtir, diğer türlerde ise değiĢim gözlemlenmemiĢtir. Çimlenen türlerin boyu 0.5-1.0 cm arasında değiĢmiĢtir.
15 C⁰‟de gözlemlenen değiĢimler : 8 genotip ve 1 tescilli çeĢit olan Doğu Yıldızında çimlenme gözlemlenmiĢtir. Ölçümler 0.5-7.0 cm arasında değiĢmiĢtir.
25 C⁰‟ de gözlemlenen değiĢimler: 7 genotip ve 1 tescilli çeĢit olan Doğu Yıldızında çimlenme gözlemlenmiĢtir. Boy ölçümleri 0.5-6.5 cm arasında değiĢmiĢtir.
35 C⁰‟ de 11 farklı dactylis genotipi ve 1 tescilli çeĢitte değiĢim gözlenmemiĢtir. Bu durum Dactylis glomerata spp. türlerinin yüksek sıcaklıklara dayanıklı olmadığını göstermiĢtir. 5-15-25 C⁰ sıcaklıklarında çimlenen tek tür Doğu Yıldız çeĢidi olmuĢtur. Doğu yıldızı çeĢidinin farklı sıcaklılar altında kolayca yetiĢebildiği gözlemlenmiĢtir. Bu çeĢitin yüksek ve düĢük sıcaklıklara kolay adapte olmasının sebebi lokasyonun Erzum olması üzerinde düĢünülmesi gereken bir konu olmuĢtur.
5.2 Öneriler
Açıkgöz (2001)‟ de domuz ayrığı üzerinde yaptığı çalıĢmalarda kültür çeĢitlerine ek olarak yanında yabani çeĢitlerininde de ıslah edilerek, bu bitkilerin doğal olarak yetiĢtikleri bölgelerde, üreticilere verim ve uyum gücü fazla olan yeni çeĢitlerin üretilmesi yem bitkileri tarımı açısından faydalı olacağını belirtmiĢtir. Ayrıca domuz ayrığında dormansi ve çimlenmesiyle ilgili çalıĢmaların yapılması gerektiğini söylemiĢtir. Nitekim yapılan bazı araĢtırmalarda domuz ayrığının 15-25 C⁰ arasında çimlendiği çimlenme gün sayımlarının 5-14 günde yapıldığı belirtilmiĢtir.
Domuz ayrığında çimlendirme gözlemlerine dayanarak çimlenme çıkıĢının olmama sebebi; dormansi olabilir. Bu durumun önüne geçmek için dormansi olabileceği düĢünülerek tohumlara hormon uygulamak, tohumları suda bekletmek, bazı kimyasal maddeler uygulayarak tekrar çimlendirme olabilir.
Ekonomik öneme sahip dactylıs glomerata ssp. çimlenmesi ile bağlantılı birkaç çalıĢma yapılmıĢtır. ÇalıĢmalardan elde edilen sonuçlar rapor haline getirilmiĢtir. Rapordaki sonuçlara göre çimlenme sonucu olarak oluĢan tür içi ve türler arasında gözlemlenen farklılıkların iklimsel ve ekolojik Ģartlara göre olduğu belirlenmiĢtir(Probert ve ark., 1985b).
Ayrıca sıcaklığın tohumlarda fizyolojik olgunlaĢma düzeyi üzerinde olumlu etkilerde bulunduğu gözlemlenmiĢtir. Pirinçte yapılan çimlendirme çalıĢmalarında pirinç tohumlarının günlük uyuma süreci ve sıcaklık arasında pozitif olmayan doğrusal bir bağlantı olduğu gözlemlenmiĢtir. Stoyanova ve Kostov (1983)‟a göre dactylıs glomerata ssp.‟de benzer durumun görüldüğü söylenmiĢtir.
Farklı sıcaklıkların çimlenmeyi uyardığı yapılan çalıĢmalarla kanıtlanmıĢtır. ÇalıĢmalarda 15 aileden 85 tür seçilerek sıcaklıklara verilen tepki gözlemlenmiĢtir. Ayrıca değiĢen sıcaklıklarda çimlenmenin değiĢmesinde ıĢığında etkisi olduğu söylenmiĢtir. Bu gözleme göre dactylıs glomerata ssp. için sabit sıcaklıklarda değiĢimin olmadığı fakat farklı sıcaklıklarda maksimum çimlenme elde edilmiĢtir (Probert ve ark., 1986). Yapılan çalıĢmalarda yukarıdaki yapılan gözlemler kanıtlanmıĢtır. Domuz ayrığı farklı sıcaklıklara farklı tepkiler vermiĢtir.
DAVIDSON ve MILTHORPE (1965)‟ e göre sıcaklığın domuz ayrığı üzerinde etkisini incelemek için 14 C⁰, 22 C⁰, 26 C⁰‟de 6 haftalık zaman içerisinde haftalık
süreçlerde ölçüm yapılmıĢtır. Gözlemlenen sonuca göre sıcaklık artıĢı ile mutlak olarak büyüme ilk haftalarda gözlemlenmiĢtir. 22 C⁰ ve 26 C⁰‟ lerde ölçüm yapıldığında ise mutlak olarak büyümede azalma yaprak-alan oranında düĢme gözlemlenmiĢ, 26 C⁰ ve üzeri sıcaklıkların büyüme üzerine etkisinin olumsuz olarak yansıdığı düĢünülmüĢtür. Özellikle 3 C⁰ ve altındaki sıcaklıklarda hücre bölünmesinin duracağını dolayısıyla domuz ayrığında canlılığın olmayacağı belirtilmiĢtir. Domuz ayrığının optimum sıcaklık aralığı 20-25 C⁰ olduğu belirtilmiĢtir.
Bu çalıĢmada 1130919 no‟lu TÜBĠTAK projesi kapsamında seçilen 11 farklı Dactylis genotipi ve 1 tescilli çeĢit olan Doğu Yıldızı laboratuvar koĢullarında yapılan çimlendirme için dört farklı sıcaklık kullanılarak gözlem yapılmıĢtır. Bu gözlemde minimum sıcaklık değeri 5 C⁰ olmuĢtur. 5 C⁰ dactylis türlerinin Doğu Yıldızı çeĢidi, 1-8 türü hariç, diğer 9 türde çimlenme olmamıĢtır.35 C⁰ ise optimum sıcaklık değerimiz olmuĢtur, fakat 35 C⁰‟ de 11 farklı türde ve 1 tescilli çeĢitte de çimlenme gözlemlenmemiĢtir. 15 C⁰ ve 25 C⁰‟lerde ise baĢarılı sonuçlar elde edilmiĢ, çimlenme gözlemlenmiĢtir. Domuz ayrığının uygun koĢullarda çimlenmesi için gerekli sıcaklık 15 C⁰- 25 C⁰ olmuĢtur. Yukarıdaki çalıĢmada 22 C⁰- 26 C⁰ ve üzerinde çimlenme ve büyümenin olumsuz etkilenmiĢtir. Yukarıda bahsetmiĢ olduğumuz çalıĢma yaptığımız çalıĢmayla desteklenmiĢtir. Domuz ayrığının çok yüksek ve çok düĢük sıcaklıklarda çimlenmediği gözlemlenmiĢtir.
KAYNAKLAR
Uncategorized References
Açıkgöz, E., 1994, Çim Alanlar Yapım ve Bakım Tekniği, Çevre Ltd, Şti. Yayınları, 4, 1-24.
Açıkgöz, E., 2001, Yem Bitkileri (3. Baskı), Uludağ üniversitesi güçlendirme vakfı, yayın (182).
Akman, N., Aksoy, F., ġahin, O., Kaya, Ç. ve Erdoğdu, G., 2007, Cumhuriyetimizin 100. yılında Türkiye‟nin hayvansal üretimi, Türkiye damızlık sığır yetiştiriciliği birliği yayınları, 4, 116.
Altın, M., 1982, Bazı Yem Bitkileri ile bunların karıĢımlarının değiĢik ekim Ģekillerindeki kuru ot ve ham protein verimleri, Türlerin Ham Protein Oranları ve KarıĢımlarının Botanik Kompozisyonları, I, Kuru ot ve ham protein verimleri. Doğa Türk Tarım ve Ormancılık Dergisi, 6 (2), 93-108.
Ayan, Ġ., Acar, Z., Kutbay, G., AĢçı, Ö., Mut, H., BaĢaran, U. ve Töngel, M., 2011, Orta Karadeniz Bölgesi‟nde Bazı Buğdaygil Yem Bitkilerinin Toplanması Tanımlanması ve Kültüre Alınma Olanaklarının AraĢtırılması TÜBĠTAK kesin sonuç raporu, Samsun.
Aygün, C., Tahtacıoğlu,L., Çakal,ġ., 2007, Yem Bitkilerinin Kültüre alınma Olanakları I, Domuz Ayrığı (Dactylis
glomerata L )Toplama ÇalıĢmaları, Türkiye VII, Tarla Bitkileri Kongresi.
Berg, C. C. ve Hill, R., 1989, Maturity effect on yield and quality of spring harvested orchardgrass forage, Crop science, 29 (4), 944-948.
Blair, B., Rhykerd, C., Mullen, R., Jones, W. ve Vorst, J., 1976, Ecological adaptation of certain forage species on shallow muck soils, Proceedings of the Indiana Academy of Science, 217-224.
Bowman, D. E. ve Law, A. G., 1964, Effects of Temperature and Daylength on the Development of Lignin, Cellulose, and Protein in Dactylis glomerata L. and Bromus inermis Leyss 1, Agronomy Journal, 56 (2), 177-179.
Bushman, B. S., Larson, S. R., Tuna, M., West, M. S., Hernandez, A. G., Vullaganti, D., Gong, G., Robins, J. G., Jensen, K. B. ve Thimmapuram, J., 2011, Orchardgrass (Dactylis glomerata L.) EST and SSR marker development, annotation, and transferability, Theoretical and applied genetics, 123 (1), 119-129.
Charmet, G., Ravel, C. ve Balfourier, F., 1997, Phylogenetic analysis in the Festuca-Lolium complex using molecular markers and ITS rDNA, Theoretical and applied genetics, 94 (8), 1038-1046.
Clifford, H. ve Dallwitz, M., 1985, The classification of Poaceae: subfamilies and supertribes, Austral. J. Bot, 33, 433-484.
ÇetinbaĢ, M. ve Koyuncu, F., 2005, Soğukta nemli katlama ve tohum kabuğunun kuĢ kirazı (Prunus avium L.) tohumlarında dormansinin kırılması üzerine etkileri, Akdeniz Üniversitesi Ziraat Fakültesi Dergisi, 18 (3), 417-423.
DAVIDSON, J. L. ve MILTHORPE, F. L., 1965, The Effect of Temperature on the Growth of Cocksfoot (Dactylis glomerata L.), Annals of botany, 29 (3), 407-417. Demirkaya, M., 2006, POLĠETĠLENGLĠKOL ĠLE OZMOTĠK KOġULLANDIRMA VE HÜMĠDĠFĠKASYON UYGULAMALARININ BĠBER TOHUMLARININ ÇĠMLENME HIZI VE ORANI ÜZERĠNE ETKĠLERĠ, Erciyes Üniversitesi Fen Bilimleri Enstitüsü Fen Bilimleri Dergisi, 22 (1), 223-228.
Duman, I., 2006, Effects of seed priming with PEG or K3PO4 on germination and seedling growth in lettuce, Pakistan Journal of Biological Sciences, 9 (5), 923-928.
Ercisli, S., Esitken, A. ve Güleryüz, M., 1997, The effect of vitamines on the seed germination of apricots, XI International Symposium on Apricot Culture 488, 437-440.
Forster, J., Cogan, N., Dobrowolski, M., Francki, M., Spangenberg, G. ve Smith, K., 2008, Functionally-associated molecular genetic markers for temperate pasture plant improvement, Plant genotyping II: SNP technology. CABI Press, Wallingford, 154-187.
Gençkan, M., 1983, Yem Bitkileri Tarımı, Ege Üniversitesi Ziraat Fakültesi Yayınları, 342-343.
Georghiou, K., Thanos, C., Tafas, T. ve Mitrakos, K., 1982, Tomato seed germination. Osmotic pretreatment and far red inhibition, Journal of Experimental Botany, 33 (5), 1068-1075.
Gordon, C., Decker, A. ve Wiseman, H., 1962, Some Effects of Nitrogen Fertilizer, Maturity, and Light on the Composition of Orchardgrass 1, Agronomy Journal, 54 (5), 376-378.
Hannaway, D., Fransen, S., Cropper, J., Teel, M., Chaney, M., Griggs, T., Halse, R., Hart, J., Cheeke, P. ve Hansen, D., 1999, Orchardgrass(Dactylis glomerata L.) Extension & Station communications, PNW, 502, 18.
Hardison, J., 1984, Stripe rust (Puccinia striiformis) on orchardgrass in Oregon, Plant Dis, 68, 1099.
Hartmann, H., Kester, D. ve Davies, T., 1990, Principal of propagation by seed, Plant propagation, principals and practices, 104-136.
Hatipoğlu, R. ve Kökten, K., 2009, Domuz ayrığı (Dactylis sp. L.), In, Eds: TC Tarım ve KöyiĢleri Bakanlığı Tarımsal Üretim ve GeliĢtirme Genel Müdürlüğü, p. Henning, J. C. ve Risner, N., 1993, Orchardgrass, Extension publications (MU).
Hilhorst, H. ve Karssen, C., 1992, Seed dormancy and germination: the role of abscisic acid and gibberellins and the importance of hormone mutants, Plant growth regulation, 11 (3), 225-238.
Horn, M., Shillito, R., Conger, B. ve Harms, C., 1988, Transgenic plants of orchardgrass (Dactylis glomerata L.) from protoplasts, Plant cell reports, 7 (7), 469-472.
Jones, K. ve Borrill, M., 1962, Chromosomal status, gene exchange and evolution inDactylis, Genetica, 32 (1), 296-322.
Karakurt, H., AslantaĢ, R. ve EĢitken, A., 2010, Tohum çimlenmesi ve bitki büyümesi üzerinde etkili olan çevresel faktörler ve bazı ön uygulamalar, Uludağ Üniversitesi Ziraat Fakültesi Dergisi, 24 (2), 115-128.
Koc, A., Gokkus, A., Serin, Y., Tan, M. ve Comakli, B., 1998, Hay yield and quality of orchardgrass, alfalfa and red clover mixtures in relation to nitrogen application, Proceedings of the 2nd Balkan Symposium on Field Crops, 16-20.
Kökten, K., Tükel, T., Hatipoğlu, R., Polat, T., Kutlu, H. R. ve Görgülü, M., 2003, GAP ve Çukurova koĢullarında biçme ve otlatmaya elveriĢli çokyıllık buğdaygil+ baklagil karıĢımlarının saptanması üzerinde bir araĢtırma.
Kunelius, H., MacLeod, L. ve Calder, F., 1974, Effects of cutting management on yields, digestibility, crude protein, and persistence of timothy, bromegrass, and orchard grass, Canadian Journal of Plant Science, 54 (1), 55-64.