T.C
SELÇUK ÜNĐVERSĐTESĐ FEN BĐLĐMLERĐ ENSTĐTÜSÜ
DEMĐR UYGULAMASININ YABANĐ VE KÜLTÜR BUĞDAYLARININ DEMĐR, ÇĐNKO VE BAKIR ĐÇERĐKLERĐNE ETKĐSĐNĐN BELĐRLENMESĐ
Sultan YAŞAR YÜKSEK LĐSANS TEZĐ
TARLA BĐTKĐLERĐ ANABĐLĐM DALI Konya, 2008
T.C
SELÇUK ÜNĐVERSĐTESĐ
FEN BĐLĐMLERĐ ENSTĐTÜSÜ
DEMĐR UYGULAMASININ YABANĐ VE KÜLTÜR BUĞDAYLARININ DEMĐR, ÇĐNKO VE BAKIR ĐÇERĐKLERĐNE ETKĐSĐNĐN BELĐRLENMESĐ
Sultan YAŞAR
YÜKSEK LĐSANS TEZĐ
TARLA BĐTKĐLERĐ ANABĐLĐM DALI
Bu tez 30.04.2008 tarihinde aşağıdaki jüri tarafından Oy Birliği ile kabul edilmiştir.
Prof.Dr. Mehmet BABAOĞLU Prof.Dr. Sait GEZGĐN Doç.Dr. Süleyman SOYLU (Danışman) (Üye) (Üye)
ÖZET
Yüksek Lisans Tezi
DEMĐR UYGULAMASININ YABANĐ VE KÜLTÜR BUĞDAYLARININ DEMĐR, ÇĐNKO VE BAKIR ĐÇERĐKLERĐNE ETKĐSĐNĐN BELĐRLENMESĐ
Sultan YAŞAR
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Tarla Bitkileri Anabilim Dalı
Danışman : Prof. Dr. Mehmet BABAOĞLU 2008, Sayfa: 71
Jüri : Prof. Dr. Mehmet BABAOĞLU Prof. Dr. Sait GEZGĐN
Doç. Dr. Süleyman SOYLU
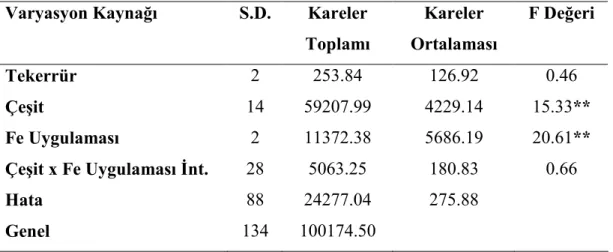
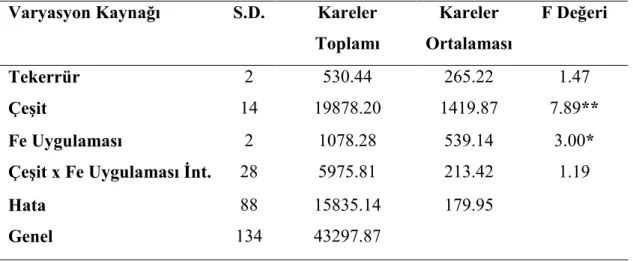
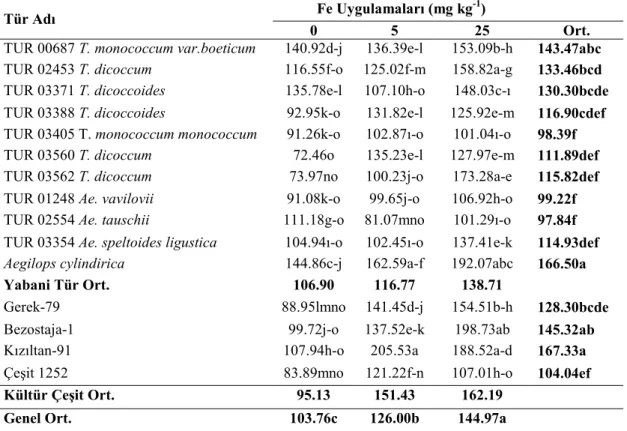
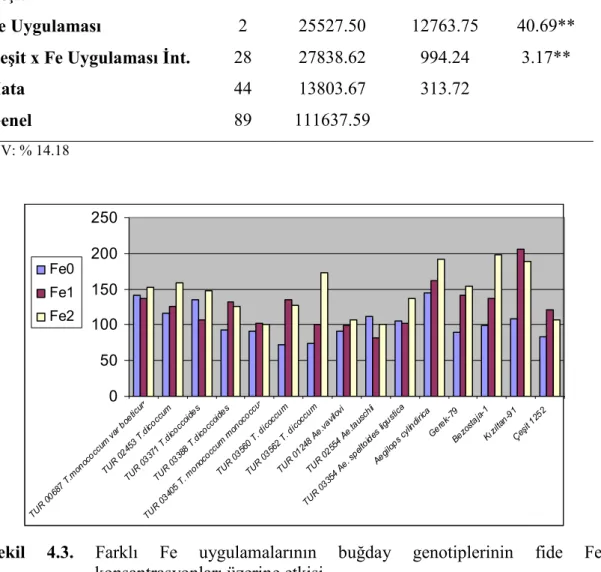
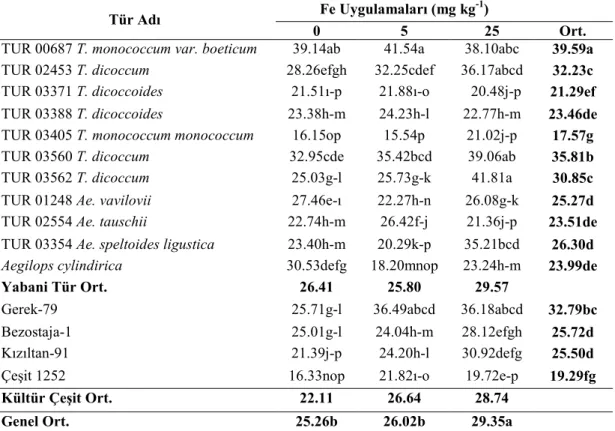
Buğday genotiplerinde demir (Fe) uygulamalarının bitkinin sürme hızı ve gücü ile fide, gövde ve tohumlarında bulunan Fe, Zn ve Cu konsantrasyonları üzerine etkisini araştırmak amacıyla yapılmış bu çalışmada materyal olarak 11 adet yabani buğday genotipi ve 2 adet makarnalık 2 adet ekmeklik buğday çeşidi kullanılmıştır. Denemede genotiplere üç Fe (0, 5, 25 mg kg-1) dozu uygulanmış, çalışma tesadüf bloklarında faktöriyel deneme desenine göre üç tekerrürlü olarak kurulmuş ve sera koşullarında yürütülmüştür.
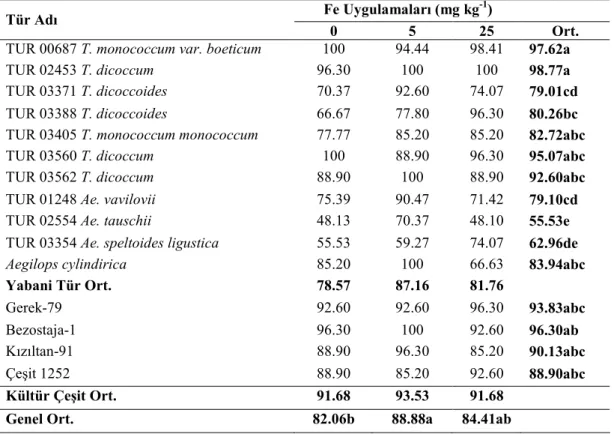
Araştırma sonuçlarına göre, fidede Zn, Cu, tohumda Fe ve Zn konsantrasyonlarının TUR 00687 T. monococcum var. boeticum genotipinde en yüksek olduğu tespit edilmiştir. Bu genotipten sonra üstün özellik gösteren genotip TUR 03354 Ae. speltoides ligustica’dır. Yabani buğday genotip fidelerinin Fe noksan şartlarda kültür çeşitlerine göre topraktan daha iyi Fe alabildikleri tespit edilmiştir. Fe dozu arttıkça fide Fe ve Cu konsantrasyonları; tohum Fe, Cu konsantrasyonları; gövde Cu konsantrasyonları doğru orantılı olarak artmıştır. En yüksek tohum Cu konsantrasyonu, TUR 03354 Ae. speltoides ligustica genotipinde görülmüştür. En yüksek gövde Zn konsantrasyonu TUR 03560 T. dicoccum genotipinde tespit edilmiştir.
Anahtar Kelimeler: Demir alımı, yabani buğday genotipleri, kültür buğdayı çeşitleri, demir (Fe), çinko Zn), bakır (Cu)
ABSTRACT MASTER THESIS
EFFECT OF IRON APPLICATION TO DETERMINE IRON, ZINC AND COPPER CONTENTS OF WILD AND CULTIVATED WHEAT
GENOTYPES Sultan YAŞAR Selcuk University
Graduate School of Natural and Applied Sciences Department of Field Crops
Supervisor : Prof. Dr. Mehmet BABAOĞLU 2008, Pages: 71
Jury: Prof. Dr. Mehmet BABAOĞLU Prof. Dr. Sait GEZGĐN
Doç. Dr. Süleyman SOYLU
11 wild, two cultivated durum and two bread wheat (Triticum sp.) genotypes were used to study on the effects of iron applications in growth rates as well as the Fe, Zn and Cu concentrations uptaken by wheat seedling shorts and grains. Three iron dosages (0, 5 and 25mg kg-1) were applied on the genotypes in green house pot experiments with 3 replications in a factorial trial design.
The results have shown that the speed of germination of wild genotypes was higher than cultivated genotypes. Zn and Cu concentrations in seedlings, Fe, Zn concentration in grains were the highest in TUR 00687 T. monococcum var. boeticum genotype followed TUR 03354 Ae. speltoides ligustica. Wild genotype seedlings takes higher iron concentrations than cultivated genotype seedlings from the soil under iron deficient conditions. The highest seed Zn concentration was found in TUR 00687 T. monococcum var. boeticum and TUR 03354 Ae. speltoides ligustica genotypes. The highest seed copper concentration was from TUR 03354 Ae. speltoides ligustica genotype.
Key Words: Iron absorption, wild wheat genotypes, wheat cultivars, iron (Fe), zinc (Zn), copper (Cu)
TEŞEKKÜR
Yüksek lisans sürecimin başlangıcından bu yana bilimsel destek ve teşviklerinden dolayı değerli danışman hocam Prof. Dr. Mehmet BABAOĞLU’na, teşekkürlerimi sunuyorum.
Tezimi hazırlarken baştan sona bana yön veren, bilgi ve desteğini esirgemeyen çok değerli hocam Doç. Dr. Süleyman SOYLU’ya sonsuz teşekkürlerimi sunuyorum.
Tez çalışmam süresince bilgi ve desteğini esirgemeyen ve laboratuvar imkanlarından faydalanmama olanak sağlayan değerli hocam Prof. Dr. Sait GEZGĐN’e sonsuz teşekkür ediyorum.
Yrd. Doç. Dr. Mustafa YORGANCILAR’a ve ihtiyacım olduğu her an desteğini hissettiğim, bilgi ve becerisiyle çalışmalarımda bana yol gösterici olan, en zor anlarda dahi pozitif düşünebilmemi sağlayan çok değerli Arş. Gör. Emine ATALAY’a sonsuz teşekkür ediyorum.
Yüksek lisans eğitimim boyunca bana daima yardımcı olan eşim, annem, babam ve kardeşlerime desteklerinden dolayı sonsuz teşekkür ediyorum.
ĐÇĐNDEKĐLER ÖZET...i ABSTRACT ...ii TEŞEKKÜR ... iii ĐÇĐNDEKĐLER...iv ÇĐZELGELER LĐSTESĐ ...v ŞEKĐLLER LĐSTESĐ...vii 1. GĐRĐŞ...1 2. KAYNAK ARAŞTIRMASI...4 3. MATERYAL VE METOT...4 3.1. Materyal...21
3.1.1. Denemede kullanılan toprak materyali...21
3.1.2. Denemede kullanılan genotipler...22
3.2. Metot ...25
3.2.1. Denemenin kurulması...25
3.2.2. Gözlem ve ölçümler ...28
3.2.3. Laboratuvar analizleri...28
3.2.4. Đstatistiki analiz ve değerlendirme...29
4. ARAŞTIRMA SONUÇLARI VE TARTIŞMA...30
4.1. Genotiplere Ait Sürme Hızı ve Gücü Yüzdeleri...30
4.1.1. Sürme hızı...30
4.1.2. Sürme gücü...32
4.2. Fide Element Konsantrasyonları ...35
4.2.1. Fide demir (Fe) konsantrasyonları ...35
4.2.2. Fide çinko (Zn) konsantrasyonları ...38
4.2.3. Fide bakır (Cu) konsantrasyonları ...40
4.3. Gövde Element Konsantrasyonları...43
4.3.1. Gövde demir (Fe) konsantrasyonları ...43
4.3.2. Gövde çinko (Zn) konsantrasyonları ...46
4.3.3. Gövde bakır (Cu) konsantrasyonları ...49
4.4. Tohum Element Konsantrasyonları ...51
4.4.1. Tohum demir (Fe) konsantrasyonları ...51
4.4.2. Tohum çinko (Zn) konsantrasyonları ...55
4.4.3. Tohum bakır (Cu) konsantrasyonları ...59
5. SONUÇ VE ÖNERĐLER ...62
ÇĐZELGELER LĐSTESĐ
Çizelge 3.1. Deneme toprağının bazı fiziksel ve kimyasal özellikler ... 21 Çizelge 3.2. Denemede kullanılan buğday çeşitleri ve yabani genotipler... 22 Çizelge 4.1. Farklı Fe uygulamalarının yabani ve kültür buğday genotiplerinde
sürme hızına etkisi (%)... 31 Çizelge 4.2. Farklı Fe uygulamalarının yabani ve kültür buğday genotiplerinde
sürme hızına etkisine ait varyans analizleri... 31 Çizelge 4.3. Farklı Fe uygulamalarının yabani ve kültür buğday genotiplerinde
sürme gücüne etkisi (%) ... 33 Çizelge 4.4. Farklı Fe uygulamalarının yabani ve kültür buğday genotiplerinde
sürme gücüne etkisine ait varyans analizleri ... 34 Çizelge 4.5. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
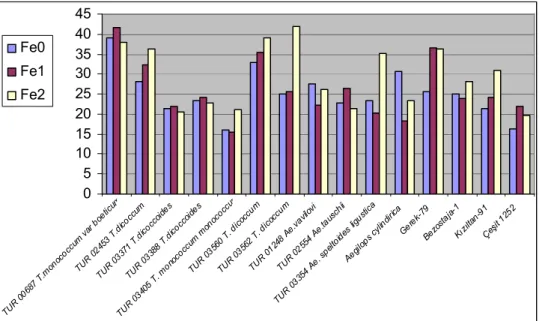
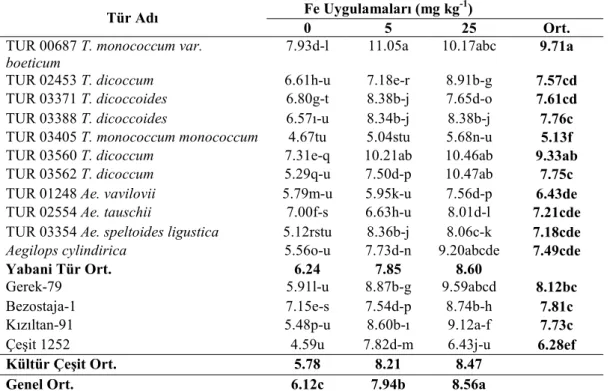
tespit edilen fide Fe konsantrasyonları (mg kg-1) ... 36 Çizelge 4.6. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen fide Fe konsantrasyonlarına ait
varyans analizleri... 37 Çizelge 4.7. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen fide Zn konsantrasyonları (mg kg-1) ... 39 Çizelge 4.8. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen fide Zn konsantrasyonlarına ait
varyans analizleri... 39 Çizelge 4.9. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen fide Cu konsantrasyonları (mg kg-1) ... 41 Çizelge 4.10. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen fide Cu konsantrasyonlarına ait
varyans analizleri... 42 Çizelge 4.11. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen gövde Fe konsantrasyonları (mg kg-1) ... 44 Çizelge 4.12. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen gövde Fe konsantrasyonlarına ait
varyans analizleri... 45 Çizelge 4.13. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen gövde Zn konsantrasyonları (mg kg-1) ... 47 Çizelge 4.14. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen gövde Zn konsantrasyonlarına ait
varyans analizleri... 48 Çizelge 4.15. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen gövde Cu konsantrasyonları (mg kg-1)... 50 Çizelge 4.16. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen gövde Cu konsantrasyonlarına ait
varyans analizleri... 50 Çizelge 4.17. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen tohum Fe konsantrasyonları (mg kg-1) ... 52 Çizelge 4.18. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen tohum Fe konsantrasyonlarına ait
varyans analizleri... 52 Çizelge 4.19. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen tohum Zn konsantrasyonları (mg kg-1) ... 57 Çizelge 4.20. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen tohum Zn konsantrasyonlarına ait
varyans analizleri... 58 Çizelge 4.21. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen tohum Cu konsantrasyonları (mg kg-1) ... 60 Çizelge 4.22. Yabani ve kültür buğdaylarında farklı Fe uygulamalarında
tespit edilen tohum Cu konsantrasyonlarına ait
ŞEKĐLLER LĐSTESĐ
Şekil 3.1. Denemeden genel bir görünüm (bitkiler 20. günde) ... 26
Şekil 3.2. Denemeden genel bir görünüm (bitkiler 65. günde) ... 26
Şekil 3.3. Bitkiler çiçeklenme döneminde... 27
Şekil 3.4. Hasat öncesi genel görünüm ... 27
Şekil 3.5. Hasat olgunluğuna ulaşmış bitkiler ... 27
Şekil 4.1. Farklı Fe uygulamalarının buğday genotiplerinin sürme hızı değerleri üzerine etkisi... 32
Şekil 4.2. Farklı Fe uygulamalarının buğday genotiplerinin sürme gücü değerleri üzerine etkisi... 34
Şekil 4.3. Farklı Fe uygulamalarının buğday genotiplerinin fide Fe konsantrasyonları üzerine etkisi ... 37
Şekil 4.4. Farklı Fe uygulamalarının buğday genotiplerinin fide Zn konsantrasyonları üzerine etkisi ... 40
Şekil 4.5. Farklı Fe uygulamalarının buğday genotiplerinin fide Cu konsantrasyonları üzerine etkisi ... 43
Şekil 4.6. Farklı Fe uygulamalarının buğday genotiplerinin gövde Fe konsantrasyonları üzerine etkisi ... 45
Şekil 4.7. Farklı Fe uygulamalarının buğday genotiplerinin gövde Zn konsantrasyonları üzerine etkisi ... 48
Şekil 4.8. Farklı Fe uygulamalarının buğday genotiplerinin gövde Cu konsantrasyonları üzerine etkisi ... 51
Şekil 4.9. Farklı Fe uygulamalarının buğday genotiplerinin tohum Fe konsantrasyonları üzerine etkisi ... 55
Şekil 4.10. Farklı Fe uygulamalarının buğday genotiplerinin tohum Zn konsantrasyonları üzerine etkisi ... 58
Şekil 4.11. Farklı Fe uygulamalarının buğday genotiplerinin tohum Cu konsantrasyonları üzerine etkisi ... 61
1. GĐRĐŞ
Hızla artan dünya nüfusunun büyük bir çoğunluğu, kısıtlı doğal kaynaklar ve bu kaynakların dengesiz dağılımı sebebiyle yetersiz beslenme ve açlık problemi ile karşı karşıya kalmaktadır (Yürür 1994). Dünyada enerji ve protein gereksinimi bakımından 800 milyon insan yetersiz beslenmekte, 2 milyara yakın insan da ‘gizli açlık’ olarak isimlendirilen mikro element (demir, bor, çinko, selenyum vb.) ve vitamin noksanlığı çekmektedir (Çakmak 2002, Welch 2002).
Temel besin kaynaklarından biri olan bitkiler, mineraller ve vitaminler yönünden oldukça zengindirler. Bununla beraber bitkiler içerdikleri mineralleri sentezleyemezler ve topraktan almak zorundadırlar (Frary ve Doğanlar 2006).
Orta Anadolu tarım topraklarının önemli bir kısmında çinko (Zn) (Çakmak ve ark. 1996) ve demir (Fe) noksanlığı (Gezgin ve ark. 2002) ile bunların hem bitkilerde hem de besin zinciri yoluyla insan ve hayvanlarda olumsuz etkileri çok yaygın olarak görülmektedir. Türkiye topraklarındaki bu yaygın mikro besin elementleriyle ilgili problemler sebebiyle bitkisel ve hayvansal besinlerde yeterince Fe bulunamamasından dolayı Türkiye’de hemen hemen her insanda Fe eksikliği ortaya çıkabilmektedir.
Demir (Fe), bitkilerde önemli fizyolojik işlevleri olan ve pek çok kimyasal tepkimeyi katalizleyen çeşitli enzimleri aktive eder. Örneğin Fe hemin enzimlerinin prostatik gruplarını oluşturur. Bu enzimler solunum zinciri içerisinde ve yükseltgenme tepkimelerinde olağanüstü önem taşıyan enerji metabolizmasında elektron taşıyıcısı olarak görev yapar (Kacar ve Katkat 1998). Fe klorofilin yapısında yer almakla beraber, bitkinin Fe beslenmesi ile klorofil içeriği arasında yakın bir ilişki bulunmaktadır (Pushnik ve Miller 1989). Yeterli miktarda yarayışlı Fe protein sentezi için gereklidir. Ortamda yeteri kadar demirin bulunmaması durumunda kloroplast hacmi ve kloroplastların protein içeriği önemli derecede azalır. Şiddetli Fe noksanlığı durumunda, hücre bölünmesi ve yaprak büyümesi olumsuz etkilenir. Ayrıca Fe, biyolojik azot bağlanması için gerekli elementlerden biridir.
Toprakta herhangi bir besin elementinin yeterli miktarda bulunması, aynı besin elementinin bitki bünyesinde de yeterli miktarda olacağını göstermez (Jones
1985, Jarrel ve Beverly 1981). Bitkinin besin element içeriği ile bitki gelişimi arasında biyolojik bir ilişki bulunmaktadır (Jones 1985). Bir bütün olarak bitkinin besin elementi içeriğinin bilinmesi kadar bitkinin farklı parçalarındaki nispi besin elementi konsantrasyonlarının bilinmesi de son derece önemlidir (Dennis 1971, Trier ve Bergman 1974). Bitki besin elementleri arasındaki etkileşim (interaksiyon) konusu çok kapsamlı ve karmaşık bir konu olup, bir besin elementinin yokluğu ya da azlığı diğer bir besin elementinin daha fazla alımına neden olabilmektedir (Güler 2002).
Đnsan beslenmesinde temel hammadde olan tahıllar, hayvan beslenmesinde ve endüstride de yaygın biçimde kullanılır. Buğday tahıllar içerisinde ilk kültüre alınan, dünyada ekim alanı, üretimi ve tüketimi en fazla olan, adaptasyon kabiliyeti yüksek bir kültür bitkisidir. Sahip olduğu büyük adaptasyon yeteneği sayesinde her türlü iklimde ve yörede yetiştirilebilme üstünlüğü buğdayı stratejik öneme sahip bir bitki haline getirmektedir. Bu anlamda artan besin maddeleri ihtiyacını karşılama yönünden tahıllar ve özelikle de buğday büyük önem taşır.
Türkiye’de buğday 21 milyon ton üretimle tarla bitkileri üretiminin yaklaşık % 35’ini oluşturmakta ve ilk sırada yer almaktadır (Anonim 2004).
Bitkisel üretimde bu denli önemli bir yere sahip tahılların Türkiye’de yaygın olarak tarımının yapıldığı alanlarda yüksek CaCO3 , yüksek pH, kil ve düşük organik
madde gibi olumsuz toprak özellikleri nedeniyle mikro element yarayışlılığı çok sınırlı düzeydedir (Altan ve ark. 1995). Bu nedenle tahıllarda verime etki eden faktörlerin araştırılması makro ve mikro besin elementlerle beslenme durumlarının belirlenmesi büyük önem arz etmektedir. Son yıllarda yapılan çalışmalarda dünya ve Türkiye topraklarında mikro besin elementleriyle ilgili beslenme problemlerinin yaygınlık gösterdiği ortaya konulmuştur (Eyüpoğlu 1999). Bu mikro besin elementleri içerisinde demir (Fe) önemli bir yere sahiptir.
Türkiye’de temel besin kaynağı olarak hemen hemen her insan buğday kaynaklı beslenmekte, un ve un mamulleriyle üretilmiş gıdalar fazlaca tüketilmektedir. Ülkemizde günlük enerji gereksinmesinin yaklaşık % 40’dan fazlası sadece buğday tarafından karşılanmaktadır. Bu oran kırsal kesimlerde olasılıkla %75’in üzerine çıkmaktadır. Diyetteki en önemli kalori ve insanların en önemli besin kaynağı olması nedeniyle Türkiye’de buğdayın statüsünün bilinmesi ve
iyileştirilmesi hem bir bilgi eksikliğinin kapatılması açısından hem de toplumsal sağlık açısından öncelikli öneme sahip bir konu durumundadır (Çakmak ve ark. 2004). Besin zincirinin en önemli halkasından birini oluşturan buğdayın ihtiva ettiği besin elementleri özellikle de Fe gibi ülkesel noksanlık gösteren mikro elementlerin oranı yeterli düzeye ulaşırsa, sadece besin kalitesi artırılmamış aynı zamanda beslenme kaynaklı birçok hastalık da önlenmiş olacaktır.
Günümüzde tüketilen bitkilerin mineral miktarlarını arttırmaya yönelik araştırmalar demir içeriğine odaklanmış durumdadır (Frary ve Doğanlar 2006). Kültür buğdaylarının mineral konsantrasyonu çok düşük düzeydedir. Modern buğdayların Fe düzeyini arttırmak için kabul gören iki temel yaklaşım gübreleme gibi agronomik düzenlemeler ve/veya genotiplerin genetiksel olarak mikro ve makro elementler bakımından iyileştirilmesidir. Gübreleme masraflı ve kalıcı olmayan bir çözümdür. Kültür çeşitlerinin ıslah programları ile iyileştirilmesi bu anlamda daha kalıcı bir çözüm olarak kabul görmektedir ve genetik tabanın oldukça daralmış olduğu tahıllarda yabani formlar bu tabanın zenginleştirilmesi için büyük önem arz eder. Yabani formlar ile ilgili yürütülecek her çalışmadan elde edilecek sonuçlar başarılı ıslah programlarının geliştirilmesinde oldukça faydalı olacaktır.
Bu tez çalışmasında materyal olarak çeşitli yabani buğday genotipleri ve kültür buğdayı çeşitleri kullanılmış, bu genotiplerin Fe ve diğer bazı element alımları (Zn ve Cu) tespit edilmiştir. Çalışma ile Fe içeriği yüksek olan genotiplerin belirlenmesi özellikle de yabani formların bu açıdan potansiyellerinin tespiti ve elde edilen verilerin buğdayın gen merkezi olan ülkemizde yürütülecek ulusal bir ıslah programında kullanılabilmesi amaçlanmıştır.
2. KAYNAK ARAŞTIRMASI
Viets (1962), farklı fraksiyonlardaki Zn’nin bitki tarafından alınabilirliliğinin farklı olduğunu, suda çözünebilir, değişebilir ve adsorbe edilmiş fraksiyonlarda bulunan Zn bitkilere kolayca yarayışlı olurken, buna karşılık birincil ve ikincil toprak mineralleriyle bulunan Zn’nin bitkilere nispeten yarayışsız olduğunu saptamıştır.
Çinkonun protein sentezinde önemli fonksiyonları bulunmakta ve noksanlığı durumunda protein sentezi hızı düşmektedir (Prask ve Plocke 1971). Çinko noksanlığında ribozomların stabilitesi bozulur, ancak Zn uygulanınca tekrar düzelir.
Bir bütün olarak bitkinin besin elementi içeriğinin bilinmesi kadar bitkinin farklı parçalarındaki nispi besin elementi konsantrasyonlarının bilinmesi de son derece önemlidir (Dennis 1971, Trier ve Bergman 1974).
Brown (1972), bazı bitkilerin köklerinden H+ iyonu ve indirgeyici bileşiklerin salgılanarak kök bölgesinde Fe+3’ün Fe+2’ye indirgeme oranının artırılmasının, kök özündeki organik asitlerin (özellikle sitratların) miktarca artmasının ve bitkilerin yetişme ortamındaki nispeten yüksek fosfora tolerans göstermesinin bitkilerin demir noksanlığına karşı uyum sağlama, demir alımı ve kullanımı ile mekanizmalarla ilgili olduğunu ifade etmiştir.
Kün’e (1979) göre buğdaygiller (Gramineae) familyasının Triticeae alt oymağında yer alan Triticum cinsi buğdayın çoğunlukla kültür türlerini, Aegilops cinsi ise yabani türlerini kapsar. Bu cinsler aralarındaki morfolojik, sitolojik ve ekolojik yakınlık nedeniyle, “Triticum Grubu” ya da “Triticum-Aegilops Grubu” adı verilen tek bir grup olarak incelenir. Yabani buğdaylar, kültür buğdaylarının genel bazı karakteristiklerini taşırlar. Aegilops cinsi, yeryüzünün 10° batı - 80° doğu boylamları ve 40°-39° kuzey enlemleri arasında, elips biçiminde kesiksiz bir dağılış alanı gösterir. Bu alan, Batı-doğu doğrultusunda Atlantik kıyısından başlayıp Akdeniz ülkeleri, G. Avrupa, Balkanlar, Türkiye, Đran ve Afganistan’dan Orta Asya’ya; güney kuzey doğrultusunda ise Sina yarımadasından Kırım yarımadası ötelerine dek uzanan ülke ve bölgeleri kapsar. Bu geniş coğrafi alan içinde, değişik Aegilops türleri değişik ekolojilere uymuşlardır. Genellikle kışları kısa ve yağışlı, yazları uzun ve kurak bölgelerin bitkisi olan Aegilops, doğada 1800 m’nin üstündeki
yükseltilerde görülmez. Cinsin yayılış alanları bir yandan yağışlı orman öte yandan çöl iklimi gibi iki ekstremle sınırlanmıştır (Zhukovsky 1928, Eig 1929).
Triticum L. ve Aegilops L. cinsleri arasında genetik yakınlığın bulunduğunu; Triticum türlerinin oluşumunda Aegilops türlerinin katkıda bulunduğunu belirten görüşler 20. yüzyıl başlarında yoğunluk kazanmıştır (Zhukovsky 1928, Eig 1929). Bu görüşler günümüzde, özellikle sitogenetik alanındaki gelişmelerinde katkısıyla güçlenme olanağına kavuşmuştur (Lilienfeld ve Kihara 1951, Kihara 1954).
Kün’e (1979) göre günümüzde bilinen 21 Aegilops türünden 12’si diploid (2x=14), 8’i tetraploid (4x=28) ve 1’i hekzaploid (6x=42) türleridir. Ayrıca iki türün (Ae. triaristata ve Ae. crassa) tetraploid ve hekzaploid formları bulunmaktadır. Türkiye, birçok buğday türlerinin ve onlara genom veren yabani türlerin ana yurdudur. Yurdumuzun geniş buğday üretim bölgelerinde, kilometrelerce uzayan alanlarda zengin Aegilops populasyonlarına rastlanır. Aegilops L. cinsinin coğrafi dağılış alanının ortasında bulunan Türkiye, diploid Aegilops’lar ve diploid buğdaylar bakımından da zengin olup; poliploid Aegilops ve Triticum türlerini ortaya çıkaran evrim olaylarının yoğun bir biçimde yer aldığı bir ülkedir. Anadolu’da tetraploid ve hekzaploid buğday kültürünün 8-10 bin yıl gibi eski bir geçmişi oluşu da buğdayın evrimine ilişkin soruların yanıtlanmasında yurdumuz materyaline gereksinme olduğunu göstermektedir.
Yüce ve Turgut’a (1980) göre yabani buğdayların çiçeklerinin küçük ve çiçek kavuzlarının sıkı olması, T. monococcum’da olduğu gibi, flamentlerin sağlam oluşu kastrasyonu büyük ölçüde güçleştirmektedir. Çiçeklenme zamanlarının kültür buğdaylarınınkinden farklılıklar göstermesi uygun bir çiçeklenme zamanının tespitini zorlaştırmakta bu nedenden dolayı melezleme materyali olarak kullanılacak yabani buğdayların sıcaklık ve üşüme isteğinin çok iyi incelenmesi gerekmektedir. Yabani buğday türleri gerek dayanıklılık genleri gerekse kalite genleri bakımından kombinasyon ıslahında yararlanabilecek önemli bir kaynaktır. Bu türlerin ana vatanı içinde Türkiye’nin de bulunuşu, adaptasyon özeliklerinin de kullanılmasına imkan vermesi açısından ıslah çalışmalarında büyük bir anlam taşımaktadır.
Toprakta herhangi bir besin elementinin yeterli miktarda bulunması, aynı besin elementinin bitki bünyesinde de yeterli miktarda olacağını göstermez (Jones 1985, Jarrel ve Beverly 1981).
Welch ve ark. (1982), Zn noksanlığı altındaki bitkilerin kök hücre membranlarının yapısal bütünlüğünü kaybettiğini ve bu nedenle iyon alımının (örneğin P) arttığını belirlemiştir.
Dünya'daki tarım topraklarının yaklaşık %30'nun (Sillanpaa 1982), Türkiye'deki toprakların ise yaklaşık %50'sinin (Eyüpoğlu ve ark. 1995) Zn bakımından fakir oldukları saptanmıştır. Türkiye'de özellikle Konya Ovası’nda söz konusu oranın % 90'a ulaştığı bildirilmiştir.
Mangan ve demir oksitlerce, aynı şekilde organik maddelerce adsorbe edilmiş şekilde bulunan bakırın büyük bir kısmı çok kuvvetli bağlı ve zor değişebilir bir şekilde bulunmaktadır (McLaren ve ark. 1983).
Aktaş ve Hatipoğlu (1984), pH’ları ve CaCO3 kapsamları farklı topraklarda
serada farklı seviyelerde Fe uygulayarak mısır yetiştirmişler, demir uygulaması yapılan topraklarda bitkinin Zn kapsamının Fe uygulamasına bağlı olarak azaldığını belirlemişlerdir.
Bakır, bütün canlıların beslenmesi için gerekli olan elementlerden birisidir. Ancak fazlalığında bitkiler ve hayvanlarda (daha çok koyunlarda) bakır toksisitesi ortaya çıkabilmektedir. Buna karşılık insanlarda kronik bakır zehirlenmesi ile pek karşılaşılmaktadır (Marian 1984).
Bitkinin besin element içeriği ile bitki gelişimi arasında biyolojik bir ilişki bulunmaktadır (Jones 1985).
Taban ve Turan (1987), toprağa artan miktarlarda verilen Fe ve Zn’nin sera şartlarında mısır bitkisinin kuru madde miktarı ile Fe, Zn, Mn, Cu, N, P ve K içerikleri üzerinde etkilerinin istatistiki bakımdan önemli olduğunu bulmuşlardır. Zn uygulamasının bitkinin Zn içeriğinde artış meydana getirdiği, bitkinin Mn içeriğini azalttığı, demir uygulamasının ise Zn içeriğinde azalmaya sebep olduğunu tespit etmişlerdir.
Turgut ve ark. (1987), doğal yabani buğday populasyonlarında morfolojik ve fizyolojik özelliklerin varyabilitesini belirlemek amacıyla yapmış oldukları araştırmada önemli sonuçlara varmışlardır. Triticum boeticum ve Triticum dicoccoides populasyonlarından tek bitki örnekleri toplanmış ve bunların döllerinde çiçeklenme zamanı, başak sayısı, sap uzunluğu, enzim desenleri ve flavonit desenleri saptanmıştır. Araştırılan tüm özellikler için populasyonlar içinde ve populasyonlar
arasında beklenmedik ölçüde yüksek bir genetik varyans bulunmuştur. Saptanan bu yüksek genetik varyabiliteden dolayı, yabani buğday populasyonlarından değerli agronomik özellikler (temel amino asitler, protein içeriği, dayanıklılık vb.) taşıyan genotiplerin seçilmesi şansının olduğu belirtilmiştir.
Demir klorofilin yapısında yer almakla beraber, bitkinin demir beslenmesi ile klorofil içeriği arasında yakın bir ilişki bulunmaktadır (Pushnik ve Miller 1989).
Demir noksanlığı altındaki çift çenekli bitkilerdeki Zn absorpsiyonunun artması strateji I bitkilerinin sahip olduğu bir özellik olan rizosfer bölgesinin asitleştirilmesine bağlı olabilir (Marschner ve ark. 1989). Zhang (1991), Zn mobilize edici salgıların salgılanmasının bitki türüne bağlı olduğunu göstermiştir. Aynı araştırmacı çift çenekli türlerin (örneğin, fasulye ve domates) kök salgılarının Zn mobilizasyonunu artırmadığını, buna karşılık buğdaygil türlerinin (örneğin, arpa ve buğday) Zn mobilizasyonunu arttırdığını bulmuştur. Ayrıca bitkilerde Zn eksikliğinin polen üretmek için başakçıkların kapasitesini azaltarak polen verimliliği ve yaşama yeteneğini olumsuz etkilediği belirlenmiştir (Sharma ve ark. 1990).
Çinko 300’e yakın enzimde rol alarak bitkide pek çok temel metabolik reaksiyonu etkilemektedir. Çinko bu enzimlerde strüktürel bileşim maddesi olarak veya bir katalizör olarak gereklidir. Bitkilerde Zn’nin temel fonksiyonları protein, karbonhidrat metabolizmasıyla ve oksin, RNA ve ribozom formasyonları ile de ilişkilidir. Çinko noksanlığında bitkide protein sentezinin gerilemesi ribozomların yapısal dayanıklılığının azalması, Zn içeren RNA polimeraz’ın aktivitesinin düşmesi ya da RNA’nın parçalanması sonucu ortaya çıkmaktadır (Obata ve Umebayashi 1988, Marschner 1995). Price (1962), çinko noksanlığında RNA sentezinin ve buna bağlı olarak da protein üretiminin durduğu saptanmıştır.
Bayraklı ve Gezgin (1991), Konya-Ereğli ve Çumra Ovası topraklarının DPTA’da çözülebilen Fe miktarının 1.80-8.60 mg kg-1 arasında değiştiğini bulmuşlar, elverişli Fe miktarı ile ele aldıkları toprak özelliklerinden sadece kil miktarı arasında istatistiki açıdan önemli (p< 0.01) bir ilişki tespit etmişlerdir.
Gezgin (1991), Büyük Konya Havzası topraklarında elverişli Fe miktarının 2.45-18.7 arasında değiştiğini bulmuştur.
Mortvedt (1991), tahıllarda demir noksanlığına bağlı olarak ortaya çıkan klorozun organik maddesi düşük kireçli topraklarda karşılaşılan yaygın bir problem olduğunu ifade etmiştir.
Yalçın ve Usta (1992), sera şartlarında yürüttükleri çalışmada Büyük Konya Havzası’na ait değişik tekstürlü 5 toprak kullanmışlar ve Zn uygulamasının mısırda gelişime etkisi ile bitkinin Zn, Fe, Mn ve Cu içeriklerini incelemişlerdir. Artan miktarda Zn uygulamasıyla bitkinin Fe, Mn ve Cu içeriklerinde azalma meydana geldiğini, Zn uygulaması ile mısır bitkisinin Fe, Mn ve Cu içeriklerinde de sırasıyla %28, %23 ve %25’lik azalmanın görüldüğünü ifade etmişlerdir.
Türkiye'deki çalışmaların yanı sıra dünyada yapılan birçok çalışmada da Zn noksanlığının bitkisel üretimde önemli verim azalmalarına yol açtığı bildirilmiştir. Örneğin Hindistan (Takkar ve ark. 1997) ve Avustralya’da (Graham ve ark. 1992) yapılan araştırmalar sonucunda topraktaki Zn eksikliğinin buğday üretim alanlarında önemli verim azalmalarına neden olduğu görülmüştür. Literatür çalışmaları, toprakta ve bitkide Zn noksanlık şiddetinin artışıyla bitkide önemli verim kayıplarının olduğunu göstermektedir.
Çinko ayrıca membran proteinlerinde (örneğin, iyon taşıyıcı proteinler) muhtemelen sülfihidril gruplarının korunmasında, antioksidantlar gibi rol almaktadır (Welch ve Norvell 1993).
Graham ve Rengel (1993), yüksek tohum Zn içeriğinin, yetersiz Zn’ye sahip toprakta gelişen buğday genotipleri için oldukça önemli olduğunu tohumu yüksek Zn içeren genotiplerin büyümenin ilk aşamalarında daha çok kök ve yeşil aksam yaptıklarını, daha fazla miktarda kök kütlesinin oluşumunun büyümenin sonraki devrelerinde bitkinin çevreden daha çok Zn almasına katkıda bulunabileceğini tespit etmişlerdir. Ancak bunun her zaman Zn noksanlığına karşı genotiplerin dayanıklılığını belirleyen bir mekanizma olmadığını belirtmişlerdir.
Anaç ve Saatçi (1993), çinko, demir ve fosfor bakımından fakir kireçli topraklarda bunların herhangi birinin ilavesi ile diğer besin elementlerinin eksikliğinin ortaya çıktığını, fosforun ve çinkonun neden olduğu Fe eksikliği ile Fe’nin neden olduğu Zn eksikliği ise bu interaksiyonlara örnek gösterilebileceğini ifade etmiştir. Aynı şekilde bitkilere bakırlı preparatların fazla uygulanması ile bitkilerde Fe eksikliği görülmektedir.
Özbek ve ark. ’na (1994) göre kirlenmemiş topraklarda bakır içeriği 2-40 ppm arasında değişirken, kirlenmiş topraklarda 1000 ppm’e kadar çıkabilmektedir. Cu; toprakta organik maddelerce, mangan ve demir oksitlerce adsorbe edilmiş bir şekilde bulunmaktadır. Bunların dışında silikatlara bağlı olarak, az miktarda da değişebilir ve çözünebilir formda bulunmaktadır.
Lu ve ark. (1995), Arabidopsis thaliana’da klorofil eksik ch5 mutantının thylakoid membranında demir (Fe) ve magnezyum (Mg) eksikliğinin etkisini irdeledikleri çalışmalarında Fe ve Mg noksanlığının A. thaliana’nın hem ch5 mutantında hem de mutant olmayan tiplerinde klorofil ve protein miktarını azalttığı, karotenoit miktarını ise önemli oranda arttırdığını tespit etmişlerdir.
Jolley ve ark. (1996), çeltik, mısır ve sorgumda, Berg ve ark. (1993), buğdayda demir noksanlığına bağlı klorozun ortaya çıktığını rapor etmişlerdir.
Fe noksanlığı durumunda toprağın iyileştirilmesi ve demir içeren yaprak gübreleri sıklıkla kullanılan yöntemlerdir. Bununla beraber bu metotlar pahalıdır. Topraktan daha fazla demir alan yüksek verimli bitki genotiplerinin yetiştirilmesi daha gerçekçi bir yaklaşımdır. Ancak kloroz sadece demir noksanlığında değil özellikle toprak yapısının homojen olmadığı alanlarda diğer çevre koşullarından da fazlaca etkilenerek ortaya çıkmaktadır. Bu sebeple demir noksanlığından daha az etkilenen genotiplerin belirlenmesi zordur. Klorozisi etkileyen diğer faktörlerin kontrol edilebildiği büyütme kabinlerinde yada laboratuvarda demir noksanlığı durumunda ortaya çıkan klorozla ilgili çalışmaların yürütülmesi ve dayanıklı genotiplerin belirlenerek geliştirilmesi için ön koşul olarak gereklidir (Jolley ve ark. 1996).
Buğdaygil bitkilerinde demir alınımının düzenlenmesi üzerine yaptıkları araştırmada bazı buğdaygil monokotiledon bitkilerin (strateji II), dikotiledon ve buğdaygil olmayan monokotiledon bitkilerden (strateji I) farklı demir kazanım stratejilerine sahip olduğu belirlemişler ve strateji II bitkilerince demir kazanım prosesinin biyosentez, salınım, çözünür hale gelme ve alınım olarak 4 ana adımda gerçekleştiğini ifade etmişlerdir. Bu proses içinde mugineik asidin önemini vurgulamışlardır. Demir tutulum prosesi, mugineik asit-Fe+3 kompleksinin oluşturulması ve spesifik alım sistemlerince düzenlenmektedir (Ma ve Nomoto 1996).
Yehuda ve ark. (1996), buğdaygil bitkilerinde demir alımında ligand değişimin rolünü irdeledikleri çalışmalarında rizofer bölgesinin arpa (Hordeum vulgare L.) ve mısır (Zea mays L.) için yararlı Fe kaynağı olduğunu, demir komplekslerinin alınım ve taşınım oranının fitositerofor salınım miktarıyla paralel olduğunu ifade etmişlerdir.
Çakmak ve ark. (1996), Dünya’da ve Türkiye'de yapılan çalışmalarda tarım topraklarında oldukça yaygın bir Zn noksanlığının olduğu bildirmiştir.
Alpaslan ve Taban (1996) çeltikte çinko-demir ilişkisini belirlemek amacıyla, bitkiye elverişli Zn kapsamı 0.4 ppm Zn olan toprak ile sera koşullarında deneme yürütmüşlerdir. Toprağa Zn, ZnCl2.7H2O şeklinde 0, 2.5, 5.0 ve 10.0 ppm Zn
düzeylerinde, demir, FeSO4 şeklinde 0, 5.0, 10.0 ve 15.0 ppm Fe düzeylerinde
uygulanmıştır. Çeltik bitkisinin kuru madde miktarları artan Zn uygulamasıyla % 21, 34 ve 44 artan demir uygulaması ile de %7, 11 ve 6 oranında artmıştır. Deneme bitkisinin Zn kapsamı uygulanan çinkolu gübrelemeye bağlı olarak %49.6, 89.5 ve 126.0 artarken, demir kapsamı ise %16.1, 27.2 ve 36.2 azalmıştır. Demirli gübreleme ile çeltik bitkisinin demir kapsamı %26.1, 66.0 ve 105.9 artmış, buna karşılık Zn kapsamı % 15.7, 28.6 ve 42.6 azalmıştır.
Kenbaev ve Sade (1997), Konya kıraç koşullarında arpa çeşitlerinin (Hordeum vulgare L.) çinko dozlarına tepkilerinin belirlenmesi amacıyla yaptıkları çalışmada çinko dozlarının tane verimi üzerine etkisini istatistiki olarak önemli bulmuşlardır. Çinko uygulamasıyla tane verimi artmakla beraber, kontrol dışındaki tüm çinko dozlarının aynı verim grubunda yer aldığını tespit etmişlerdir.
Yetiştirilen bitkilerin ürün miktarları ve kaliteleri, toprakların besin maddesi içeriklerinin bitkilerin ihtiyaçlarını iyi bir şekilde karşılamasıyla yakından ilgilidir. Bitkilerin ihtiyaç duydukları besin elementlerinin toprakta yeter ölçüde ve uygun oranda bulunmadığı ya da bulunsa bile bitkilerin çeşitli faktörler sebebiyle bunlardan yararlanamadığı hallerde bitkiler normal gelişememekte, ürün miktarı düşmekte ve kalite bozulmaktadır (Arcak ve ark. 1997).
Türkiye’deki tarım topraklarının %23.08’inin az kireçli, geri kalan %76.92’lik bölümünün ise kireçli ve çok fazla kireçli özellikte olduğu (Eyüpoğlu 1999), tarım topraklarının %26.87’sinde başka bir anlatımla 7.5 milyon ha’lık kısmında yarayışlı Fe içeriğinin 4.5 mg kg-1 ın altında ve yetersiz olduğu, bitkiler için
yarayışlı Fe içeriği ile toprak pH’sı ve kireç içerikleri arasında negatif korelasyonların bulunduğu belirlenmiştir (Eyüpoğlu ve ark. 1998).
Toprakta demir Fe+3 ve Fe+2 formlarında bulunmaktadır. Bitkiler metabolizmalarında demiri Fe+2 iyonu şeklinde kullandıkları için kökleriyle Fe+2 iyonunu ya da bu şekle indirgenmiş demiri alırlar. Fe elementinin alımı sırasında gösterdikleri farklılıklar sebebiyle bitkiler Fe etkin (strateji I bitkileri) ve Fe etkin olmayan bitkiler (Strateji II bitkileri) olarak iki grupta toplanmıştır. Fe etkin bitkiler olan çift çenekli bitkiler yeteri kadar Fe alamadıklarında çeşitli yollarla gidermeye çalışmışlardır. Örneğin anılan bitkiler köklerinde daha fazla kök tüyü oluştururlar ve rizosfer bölgesindeki pH’yı salgıladıkları H+ iyonları ile düşürüp ortamdaki Fe+3’ün indirgenmesini sağlayarak yada toprak kolloidlerince fiske edilmiş Fe’in çözünürlülüğünü artırarak Fe ihtiyaçlarını karşılarlar. Demirden yararlanma yönünden anılan mekanizmaya sahip olmadıkları için Fe içerikleri düşük ve pH’sı yüksek topraklarda yeterli Fe alamayan ve Fe noksanlığı gösteren bitkiler “Fe etkin olmayan” bitkilerdir (Strateji II). Buğdaygil bitkileri Strateji II mekanizmasıyla topraktan Fe alırlar. Buğdaygil bitkileri kökleriyle salgıladıkları fitesiderofor adı verilen maddelerle rizosferde yarayışlı şekle dönüştürerek demiri alırlar. Fitesiderofor adı verilen maddeler mugineik ve avenik asitler gibi protein olmayan aminoasitlerdir (Kacar ve Katkat 1998).
Bitki besin elementi içeriği ile toprak özellikleri arasında çok önemli bir bağlantı mevcuttur. Türkiye topraklarının besin elementlerinin alımını etkileyen yüksek pH ve kireç, düşük organik madde ve dengesiz gübreleme gibi nedenlerle çoğu kez farklılıklar arz etmesi dünyada farklı şartlarda yapılan çalışmaların ülkemiz koşullarında kullanılabilirliğini sınırlamakta ve bu bakımdan detaylı çalışma yapılması gerekliliğini ortaya koymaktadır.
Türkiye’deki tarım topraklarının %23.08’inin az kireçli, geri kalan %76.92’lik bölümünün ise kireçli ve çok fazla kireçli özellikte olduğu (Eyüpoğlu 1999), tarım topraklarının %26.87’sinde başka bir anlatımla 7.5 milyon ha’lık kısmında yarayışlı Fe içeriğinin 4.5 mg kg-1 ın altında ve yetersiz olduğu, bitkiler için yarayışlı Fe içeriği ile toprak pH’sı ve kireç içerikleri arasında negatif korelasyonların bulunduğu belirlenmiştir (Eyüpoğlu ve ark. 1998).
Demir, bitkilerde önemli fizyolojik işlevleri olan ve pek çok kimyasal tepkimeyi katalizleyen çeşitli enzimleri aktive eder. Örneğin demir hemin enzimlerinin prostatik gruplarını oluşturur. Bu enzimler solunum zinciri içerisinde ve yükseltgenme tepkimelerinde olağanüstü önem taşıyan enerji metabolizmasında elektron taşıyıcısı olarak görev yapar (Kacar ve Katkat 1998).
Çakmak ve ark. (1998), fitositerofor salınım oranı üzerine farklı ışık yoğunluklarının (50 – 600 mol.m2 s-1) etkisini, ekmeklik ve makarnalık buğday çeşitlerinde Zn noksanlığında ve arpada Fe noksanlığında araştırdıkları çalışmalarında kontrollü çevre şartlarında besin solüsyonlarında buğday bitkisi 15 gün, arpayı 11 gün kültüre almışlar, hem çinko hem de demir eksikliği altında köklerden fitositerofor salınımında ışık yoğunluğunun rolünü dikkate değer bulmuşlar, tahıllarda tür içi ve türler arasında Zn ve Fe verimliliğinin ve fitosterfor salınımının belirlenmesi ile ilgili çalışmalarda ışık durumlarına özel önem verilmesi gerektiğini belirtmişlerdir.
Robinson ve ark. (1999), topraktan demir alımıyla ilgili olan ferric-chelate reductase’ı inceledikleri çalışmada FRO2 genini izole etmişler ve bu genin membranlara boydan boya elektron taşıyıcı aracı flavositokromların üst familyasına ait olduğunu ifade etmişlerdir. FRO2’nin izolasyonu; demir noksanlığı olan topraklarda bitki büyümesi ve besin kalitesinin artırılması açısından anlamlı olduğu vurgulanmıştır.
Köhl ve Römheld (1999), kök apoplastlarının bitkilerdeki demir birikiminde rolünü belirlemek için yürüttükleri çalışmalarında apoplastik demir konsantrasyonunun hem strateji I hem de strateji II bitkilerinde düşük olması sebebiyle demir birikiminde daha önce düşünüldüğünden daha az etkili olduklarını belirlemişlerdir.
Zhang ve ark. (1999), kireçli topraklarda büyüyen buğdaygillerin (buğday) ve dikotiledon bitkilerin (soya ve hıyar) kök apoplazmında Fe birikimini inceledikleri çalışmalarında su kültürü kullanmışlar, sınırlı demir alınımı durumlarında bu bitkilerin köklerinde biriken apoplazmik Fe’nin, mobilize edilebildiğini ve bitkinin Fe içeriğinin iyileştirilmeye çalışıldığını belirlemişler, buğdaygillerde fitositerofor salgısının artışı ile köke taşınabilen ve kök apoplastlarındaki Fe birikiminin arttığını ifade etmişlerdir.
Strasser ve ark. (1999), toprakta yetiştirdikleri Strateji I ve Strateji II bitkilerinde demir apoplast konsantrasyonu kullanılan farklı topraklar tarafından az miktarda da etkilendiğini belirlemişlerdir.
Chang ve ark. (1999), buğdayda (Triticum aestivum L.) alüminyumun (Al) fitositerofor üzerindeki etkisi ve Fe-fitositerofor kompleksini irdeledikleri çalışmalarında ortama Al eklenmesinin 2'-deoksimugineik asit’e olan Fe çözünülebilirliğini etkilemediğini, bununla beraber bakır (Cu) eklenmesinin önemli ölçüde çözünülebilirlik kapasitesini inhibe ettiğini tespit etmişlerdir. Alüminyumun fazlaca bulunduğu ortamda fitositerofor Fe+3 kompleksinin alımındaki azalmanın alümniyumun köklerde yaptığı zarar sebebiyle olduğu ve direkt olarak fitositerofor Fe+3 kompleksininin bloke edilmediği belirlenmiştir.
Rengel ve Römheld (2000a), 8 ekmeklik ve 2 makarnalık buğday genotipinde kök salgıları, Fe alınımı ve taşımını ele aldıkları çalışmalarında aynı zamanda Zn noksanlığına toleransını da incelemişler, ekmeklik buğday genotiplerinden Aroona, Exalibur ve Stilleta’nın Zn ve Fe noksanlığına tolerans gösterdiklerini, makarnalık buğday genotiplerinin ise toleransının düşük olduğunu belirlemişlerdir. Buğday genotiplerinin Zn noksanlığına gösterdikleri tolerans ile salgıladıkları fitositerofor miktarı arasında olumlu bir ilişki olduğu, fitosterofor miktarının artması ile toleransın da arttığı belirlenmiştir.
Rengel ve Römheld (2000b), Zn ve Fe içermeyen besin çözeltilerinde 8 adet T. aestivum L. ve 2 adet T. turigidum L. conv. durum çeşidini kültüre almışlar ve bu koşullarda Durati ve Yallaroi makarnalık buğday çeşitlerinin Zn noksanlığına duyarlı, Fe eksikliğine ise kısmi toleranslı olduklarını tespit etmişlerdir. Çalışmada Fe noksanlığında bütün buğday genotiplerinin ciddi şekilde sarardıkları, bununla birlikte sürgün üretiminin ve kök kuru maddesinin bu durumdan çok etkilenmediği, yapraklardaki klorofilin oransal olarak azlığı ile oransal sürgün büyümesi arasında bir bağlantı oluştuğu belirlenmiştir.
Dünya topraklarının yaklaşık %39’u bitkilerin mineral alımını ciddi boyutta engelleyen kireçli topraklardan oluşmaktadır. Dünyanın kurak ve yarı kurak iklim özelliklerine sahip bölgelerindeki kireçli ve alkali reaksiyonlu topraklarda yetiştirilen pek çok bitkide de bu nedenle önemli görevleri olan demir (Fe) elementinin eksikliği
görülmektedir ve bu durum dünya populasyonunun %30’unu etkileyen önemli bir mineral noksanlık sorununa neden olmaktadır (Başar ve Taban 2001).
Başar ve Taban (2001), değişik demir bileşikleri ve uygulama yöntemlerinin, serada yetiştirilen soya fasulyesinin toplam ve aktif demir içeriği ile bazı verimlilik özellikleri üzerine etkisini araştırmışlardır. Araştırma sonuçlarına göre; yapraktan FeSO4 verilmesi soya fasulyesinin Fe içeriğini en fazla arttıran uygulama olurken,
FeEDDHA’nın da yaprakların Fe içeriğini belirgin şekilde arttırdığı, incelenen bütün özellikler üzerine en etkili bileşiğin FeEDDHA olduğu görülmüştür. Diğer uygulamaların etkisinin bu iki uygulama kadar belirgin olmadığı ifade edilmiştir.
Katkat ve Çilözgüven (2001), toprağa artan miktarlarda verilen çinkonun mısır bitkisinin çinko, demir, mangan ve bakır içerikleri üzerine etkisini incelemişler; uygulanan çinkonun mısır bitkisinin çinko, demir, mangan ve bakır içeriğine etkisinin %1 düzeyinde önemli olduğunu, artan miktarlarda uygulanan çinkoya bağlı olarak mısır bitkisinin çinko içeriği artarken, demir, mangan ve bakır içeriklerinin ise azaldığını ifade etmişlerdir.
Metabolizma faaliyetlerinin sağlıklı bir şekilde işleyebilmesi için besin maddelerinin yeterli ve dengeli alınması son derece önemlidir. Gelişmekte olan ülkelerde bitki kaynaklı besinler, günlük beslenme diyetinde önemli bir yer tutar. Ancak bitkisel üretimde yaşanan bazı problemler sebebiyle bu ülkelerde Fe eksikliğine bağlı kansızlık, iyot eksikliği ile ortaya çıkan guatr gibi mikro besin elementlerindeki noksanlıklara bağlı problemler yaşanmaktadır. Mikro besin eksikliği sonuçları insan varlığı için alarm verici ve derindir. Gelişmekte olan ülkelerde bu problemler 3 milyardan fazla insanı etkilemektedir. Bu probleme desteklenebilir çözümler bulmak için zirai yaklaşımlar acilen gereklidir (Welch 2002).
Çakmak’a (2002) göre, 21. yüzyılda hızla artan dünya nüfusu ile birlikte, tarıma uygun alanların ve ihtiyaç duyulan suyun sınırlı oluşu, ürün verimliliğinde ciddi azalmaların yanı sıra besin güvenliğini de risk altına alan küresel bir tehdit oluşturmaktadır. Yetersiz gübreleme, toprakta mineral besinlerin yetersiz bulunuşu, bitki besin elementlerinin noksanlığına veya toksisitesine toleranslı bitki genotiplerinin eksikliği, besin kaynaklarının güvensizliği, kötü beslenme ve ekosistem bozulmaları bitkisel üretimi sınırlayıcı faktörler olarak ifade edilmiştir. Bu
durumda, bitki besleme ile ilgili araştırmaların bitki genetiği ve moleküler biyolojisi ile entegrasyonu, besin miktarı eksik ve toksik toprak durumlarına adapte olmada, yenilebilir bitkisel ürünlere (örneğin tahıl ürünleri) daha çok mikro besinlerin temin edilmesinde, yüksek genetik kabiliyetli bitki genotiplerini geliştirmek vazgeçilmez olmaktadır.
Orta Anadolu tarım topraklarının önemli bir kısmında çinko (Zn) (Çakmak ve ark. 1996) ve demirin (Fe) noksanlığı (Gezgin ve ark. 2002) ile bunların hem bitkilerde hem de besin zinciri yoluyla insan ve hayvanlarda olumsuz etkileri çok yaygın olarak görülmektedir. Türkiye topraklarındaki bu yaygın mikro besin elementleriyle ilgili problemler sebebiyle bitkisel ve hayvansal besinlerde yeterince Fe bulunamamasından dolayı Türkiye’de hemen hemen her insanda Fe eksikliği ortaya çıkabilmektedir.
Yetersiz Fe içeriğine sahip ürünlerle beslenen insanlarda demir noksanlığı ve kan değerlerinde düşüşler ile çeşitli sağlık problemleri yaygın olarak görülmektedir (Çakmak 2002, Welch 2002).
Bu nedenledir ki buğdayın anavatanı olarak gösterilen ülkemizde yabani ve ilkel buğday gen kaynaklarının besin elementi açısından taranması ve varyasyonun hangi düzeyde olduğunun bilinmesinde yarar vardır.
Bitkilerin mikroelement çeşitliliği için yetiştirilmesi, mikroelement açısından kötü beslenmeye karşı savaşta kullanılabilecek güçlü bir araçtır (Welch 2002).
Bitki besin elementleri arasındaki etkileşim (interaksiyon) konusu çok kapsamlı ve karmaşık bir konu olup, bir besin elementinin yokluğu ya da azlığı diğer bir besin elementinin daha fazla alımına neden olabilmektedir (Güler 2002).
Fe eksikliği, tahıllarda yaygın bir beslenme problemidir. Fe eksikliği, özellikle kireçli, kurak ve yarı kurak bölgelerdeki topraklarda, genellikle ürün kaybıyla sonuçlanmaktadır (Zhang ve ark. 2002).
Son yıllarda mısır, çeltik, buğday ve fasulyede yüksek biyoelverişliliğe sahip Fe, Zn ve vitamin A içeren genotiplerin geliştirilmesi amacıyla yoğun şekilde çalışılmaktadır (Welch 2002).
Vert ve ark. (2002), demir eksikliğine yanıt olarak, Arabidopsis köklerinin IRT1’in ekpresyonunu arttırdığını ve divalant katyon taşıyıcı sentezlediğini belirtmişler, Arabidopsis’ te IRT1 mutant haldeyken klorosisin görüldüğünü, bitkinin
bir süre sonra öldüğünü ancak dışardan Fe uygulamasıyla bitkilerin yaşayabildiklerini ifade etmişlerdir. Demirin alınamadığı ya da demir noksanlığı durumlarında bitki köklerinin dış hücre katmanlarında IRT1’in ifade edilmesinde artış tespit etmişler, mutasyon durumunda diğer divalant katyonların alınımında da sıkıntı olduğunu, bu anlamda IRT1’nin demir başta olmak üzere eksiklik durumunda metal alımından sorumlu en önemli taşıyıcı olduğunu göstermişlerdir.
Welch ve Graham (2002), fasulye ile ilgili yaptıkları çalışmalarda genotipler arası farklılıkları tanede demir ve çinko içeriklerine göre ortaya çıkarmaya çalışmışlar, bu amaçla taradıkları 1000 genotipte tane demir içeriklerinin 34 - 89 µg Fe .g-1, çinko içeriklerinin de 21 - 54 µg Zn .g-1 arasında değiştiğini bulmuşlardır. Bu sonuçlar ilgili elementler bakımından fasulyede geniş bir genetik varyabilite olduğunu, tane demir içeriğinin %80, Zn içeriğini en az %50 oranında artırılabileceğini göstermiştir.
Erdal ve Kızılgöz (2002), tarla koşullarında yapmış oldukları araştırmada, farklı buğday genotiplerinde (Triticum spp. 073/44, Dağdaş, Fandango, Warigal-5RL, Gun-91, SBVD-2-23, Bülbül, Kıraç, P. Niska, Zitavka, ES-14, BDME-19, Kate-A, Kırkpınar, BDME-9, ES-90-3, ES-90-8-1, ES-90-14) ekimde kullanılan tohum ile hasatta elde edilen tanenin çinko, mangan, bakır ve demir içerikleri arasındaki ilişkileri incelemişlerdir. Elde edilen sonuçlara göre, tohumun mikro element konsantrasyonunun, bu tohumlardan elde edilen tanenin mikro element konsantrasyonu üzerine olumlu etkisinin olduğu belirlenmiştir. Yüksek mikro element içerikli tohumlardan elde edilen tanenin mikro element konsantrasyonu daha yüksek bulunmuştur.
Doğan ve ark. (2002), çinko içerikli bir bileşik olan Teprosyn F-2498’in ekmeklik buğday çeşitlerinin tane verimi ve verim komponentlerine olan etkisini belirlemek amacıyla yürüttükleri çalışmada, tohumlara ekim öncesi 4, 8 ve 12 kg Zn. ton-1 dozlarında bileşik uygulamışlardır. Araştırma sonucunda çinko bileşiği uygulamasının tane verimi ve verim komponentlerinde istatistiki açıdan önemli bir farka neden olmadığı belirlenmiştir. Ancak istatistiki olarak önemli olmamakla birlikte çinkonun bazı dozları verimde rakamsal olarak kısmi artışlara neden olmuştur.
Başar ve ark. (2002), değişik Zn bileşiklerinin farklı sürelerle tohumla temas ettirilmesinin, mısırın Zn, kuru madde miktarı ve bazı besin elementleri içerikleri üzerine etkisinin belirlenmesi amacıyla araştırma yapmışlardır. Araştırmada, Zn bileşikleri ve konsantrasyonları olarak, %30 ZnO, %60 ZnO ve Teprosyn F-2498 kullanılmıştır. Bu bileşikler, 0 (çok kısa), 1, 12 ve 24 saat sürelerle tohumla temas ettirilmiştir. Araştırmadan elde edilen sonuçlara göre, Teprosyn F-2498'in 0 (çok kısa) ve 1 saat sürelerle tohumla temas ettirilmesiyle, mısırın Zn içeriği ve kuru madde miktarlarında en yüksek artışlar belirlenmiştir. ZnO' nun % 30 ve %60' lık çözeltilerinin ise 0 ve 1 saatlik sürelerde Teprosyn kadar etkili olamadığı belirlenmiştir.
Gezgin ve ark. (2002), Orta ve Đç Anadolu tarım alanlarından topladıkları 667 adet toprak örneğinin analiz sonuçlarına göre, bitkiye elverişli demir kapsamlarının 0.20-98.38 ppm (ort. 4.31 ppm) arasında değiştiği tespit etmişlerdir. Araştırıcılar bu çalışmada toprakların elverişli demir kapsamının %42.7’sinde noksan (<2.5 ppm), %27.5’inde orta (2.5-4.5 ppm), %20.1’inde yeterli (4.5-9 ppm) ve %9.7’sinde yüksek (>9 ppm) düzeyde bulunduğunu bildirmişlerdir.
Zengin ve Şeker (2003), buğday bitkisinin besin elementi kapsamına toprak özelliklerinin etkilerini belirlemek amacıyla yapmış oldukları çalışmada; toprak özellikleri ile bitkinin besin elementi kapsamı arasında önemli regresyon ilişkileri saptamışlardır.
Connolly ve ark. (2003), Arabidopsis bitkisinde demir noksanlığı durumunda gen ekspresyonlarının nasıl düzenlendiğini araştırdıkları çalışmalarında Arabidopsis FRO2 geninin, kök yüzeyinde Fe indirgenmesinden sorumlu ferric chelate reduktase’ı kodladığını belirlemişlerdir. Demir alımından sorumlu FRO2 ve IRT1’in, demir noksanlığında birlikte aktifleştiklerini, demirin tekrar sağlanmasıyla baskılandıklarını, topraktan demir alımında IRT1 ve FRO2’ye düşen roller birbiriyle tutarlı olduğunu belirlemişlerdir. Demir noksanlığında 35s-FRO2 transgenik arabidopsis bitkilerinde, transgenik olmayanlara göre ferric chalate aktivitesinin daha yüksek olduğunu ve bitkilerin diğerlerine göre daha iyi büyüdüklerini belirlemişler, demir alınımında hızı sınırlayan basamağın ferric demirin (Fe+3) ferro demire (Fe+2) indirgenmesi olduğunu belirtmişlerdir. Aynı zamanda bu genlerin bakır ve kadmiyum alınımında da aktif görev aldıklarını ifade etmişlerdir.
Zhang ve ark. (2003), kireçli topraklarda yetiştirilen bitkilerde özellikle tahıllarda demir noksanlığının önemli bir problem olduğunu ifade etmişler, fitosiderofor salınımı ile buğdaygillerde demir noksanlığına dayanma kabiliyeti arasındaki pozitif bir ilişkinin varlığı üzerinde durmuşlardır. Kontrollü çevre koşulları altında noksan ve yeterli demir konsantrasyonlarına sahip besin ortamlarında yetiştirdikleri makarnalık buğdaylar, Aegilops ve bunların melezlerinden oluşan 20 genotipte fitosterofor salınımını analiz etmişler, kültür buğdayının köklerinden fitositeroforun salınım oranının yabani genotipler kullanılarak değiştirilebileceğini, yabani genotiplerin ortaya koyduğu etkilerin ebeveynlerin farklı kullanıldığı melezlemelerde farklı olabileceğini, Ae. speltoides tauschii ve Ae. cylindrica Host’un kültür buğdaylarında fitositerofor salınım oranının gelişmesinde potansiyel ve ümit verici olabileceğini ifade etmişlerdir. Đhtiyaç duyulan kültür çeşitlerini geliştirmek ve yeniden yapmak için tür içi ve türler arası melezlemeye öncelik verilmesi gerektiği vurgulanmıştır.
Vasconcelos ve ark. (2003), transgenik çeltikte ferritin geniyle artırılan demir ve çinko tutulumunu inceledikleri çalışmada soya ferritin geni ekspresyonu transgenik çeltikte demir ve çinko seviyelerinin yükselme eğiliminde olduğunu tespit etmişlerdir. Söz konusu çalışmada çeltikteki soya ferritinin ekspresyonunun, tahıl besin seviyelerini geliştirmede işlenmiş pirinçte dahil etkili olduğu kanıtlanmıştır. Transgenik çeltikte ayrıca çinko konsantrasyonu da incelenmiş ve çinko konsantrasyonunun kontrol durumuna göre transgenik tohumlarda daha yüksek olduğu tespit edilmiştir.
Birleşmiş Milletler Gıda ve Tarım Organizasyonu’na (FAO 2004) göre, 2000-2002 yılları arasında yaklaşık 852 milyon insanın yeterli beslenemediği belirtilmektedir.
Chen ve ark. (2004), Commelina communis’te demir eksikliğinin bakır (Cu) tutulması ve birikmesine sebep olduğuna, Cu tutulumu ve birikmesi muhtemelen kök ferric-chelate reductase aktivitesine bağlı olduğuna ilişkin güçlü verilere ulaşmışlardır.
Sharma ve ark. (2004), farklı demir dozlarında arpada kadmiyum (Cd) toksitesini belirlemeye yönelik araştırmalarında, oksidatif stres ve katalaz aktivitesinin hem Cd hem de Fe tarafından uyarıldığını tespit etmişler, Fe ve Cd
arasındaki antogonistik etkinin varlığının gen ekspresyonu seviyesinde doğrulandığını vurgulamışlardır.
Kocakaya ve Erdal (2005), yaptıkları çalışmada Van yöresinde yetiştirilen 6 farklı buğday çeşidi (Kırgız 95, Karacabey 97, Palandöken 97, Doğukent 1, Kutluk 94, Çukurova 86) ile 4 farklı buğday hattının (Tir 2, Tir 6, Tir 7, Tir 9) Zn uygulamasına (2 kg Zn da-1) göstermiş olduğu tepkiyi incelemişlerdir. Çinkonun etkisini değerlendirmek için, bitkinin yeşil aksam ve tane Zn içeriği ile verim sonuçlarını incelemişlerdir. Araştırmacıların elde ettikleri bulgulara göre, Zn uygulamasına bağlı olarak bütün çeşit ve hatların yeşil aksam ve tane Zn içerikleri ile verim miktarları artmış fakat elde edilen artışlar çeşid ve hatlara göre farklılık göstermiştir.
Hamurcu ve ark. (2006a), kontrollü sera koşullarında farklı seviyelerde demir uygulamalarının makarnalık buğdayın kuru madde miktarı, demir konsantrasyonu ve içeriği, Zn, Cu, Mn konsantrasyonları üzerine etkisini belirlemek amacıyla yaptıkları araştırma sonuçlarına göre, bitkide uygulanan demir miktarı arttıkça demir konsantrasyonunun belli bir noktaya kadar artış gösterdiği, belli bir seviyeden sonra düştüğü belirlenmiştir.
Hamurcu ve ark. (2006b), kontrollü sera koşullarında farklı seviyelerde uygulanan demir (Fe) dozlarının bezelye hatlarında kuru madde miktarı, bitki boyu, Fe konsantrasyonu ve miktarı üzerine etkisini belirlemek amacıyla yaptıkları çalışmada Fe (0, 6, 60 mg kg-1) üç farklı seviyede uygulamıştır. Bezelye hatlarının kuru madde miktarı, bitki boyu, Fe konsantrasyonu ve miktarı üzerine Fe uygulamalarının ve interaksiyonlarının etkisi istatistiki olarak önemli (p<0.01) bulunmuştur. Bitkiye uygulanan Fe miktarı arttıkça Fe konsantrasyonunun artış gösterdiğini belirlemişlerdir.
Karagöz ve ark. (2006), Türkiye’nin değişik bölgelerinden topladıkları yabani buğday (Aegilops L. ve Triticum L.) populasyonlarını populasyonlar arası farklılıkları ortaya koymak amacıyla agro-morfolojik yönden karakterize etmişlerdir. En yüksek değişim katsayısı sap sayısında, en düşüğü ise gelişme formunda bulunmuştur. Kümeleme analizi türlerin kendi özel gruplarında yer aldığını göstermiş, birbirine çok yakın alanlardan toplanan örnekler arasında bile farklılıklar olduğu saptanmıştır. Bu durumda ex-situ koruma programlarının mümkün olan en
geniş varyasyonu yakalayabilmeleri için kısa mesafelerde örnekleme yapmalarının gerekli olduğu sonucuna varılmıştır.
3. MATERYAL VE METOT
3.1. Materyal
3.1.1. Denemede kullanılan toprak materyali
Sera saksı denemesinde kullanılan toprak örneği seçilirken demir miktarının düşük ve CaCO3 miktarının yüksek olmasına dikkat edilmiştir. Denemede, mikro
besin elementleri yönünden noksan ve yüksek kireç içeren, Konya Merkeze bağlı Sağlık Kasabası’ndan temin edilen toprak kullanılmıştır.
Denemede kullanılan toprağın bazı fiziksel ve kimyasal özellikleri Çizelge 3.1’de sunulmuştur.
Çizelge 3.1. Deneme toprağının bazı fiziksel ve kimyasal özellikleri
Özellik Miktar Kil (%) 18.3 Silt (%) 14.3 Kum (%) 67.4 Tekstür sınıfı Kumlu tın pH (1:2.5) 8.1 EC (1:5) (µS cm-1) 125 CaCO3 (%) 31.3 Organik madde (%) 4.9
Ekstrakte edilebilir Ca (me/100 g) 10.85
Ekstrakte edilebilir K (me/100 g) 0.21
Ekstrakte edilebilir Mg (me/100 g) 0.70
Ekstrakte edilebilir Na (me/100 g) 0.082
Elverişli Cu (ppm) 0.2 Elverişli Fe (ppm) 0.9 Elverişli Mn (ppm) 2.4 Elverişli Zn (ppm) 0.1 B (ppm) 0.2 P (ppm) 17.7 Tarla kapasitesi (%) 22.5 Toplam N (%) 0.2
3.1.2. Denemede kullanılan genotipler
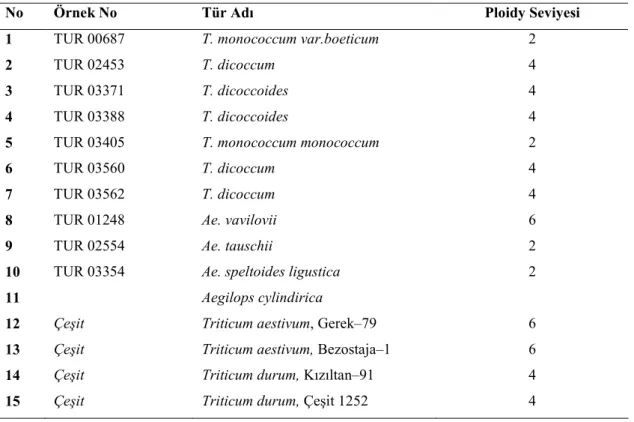
Bu çalışmada 11 adet yabani buğday genotipi ve 2 makarnalık, 2 ekmeklik olmak üzere 4 adet kültür buğdayı çeşidi kullanılmıştır. Denemede kullanılmış olan yabani genotipler Ankara Tarla Bitkileri Merkez Araştırma Enstitüsü’nden, kültür çeşitleri ise Konya Bahri Dağdaş Uluslararası Kışlık Hububat Araştırma Enstitüsü’nden temin edilmiştir. Denemede kullanılan buğday genotipleri Çizelge 3.2.’de sunulmuştur.
Çizelge 3.2. Denemede kullanılan buğday çeşitleri ve yabani genotipler
No Örnek No Tür Adı Ploidy Seviyesi
1 TUR 00687 T. monococcum var.boeticum 2
2 TUR 02453 T. dicoccum 4
3 TUR 03371 T. dicoccoides 4
4 TUR 03388 T. dicoccoides 4
5 TUR 03405 T. monococcum monococcum 2
6 TUR 03560 T. dicoccum 4
7 TUR 03562 T. dicoccum 4
8 TUR 01248 Ae. vavilovii 6
9 TUR 02554 Ae. tauschii 2
10 TUR 03354 Ae. speltoides ligustica 2
11 Aegilops cylindirica
12 Çeşit Triticum aestivum, Gerek–79 6
13 Çeşit Triticum aestivum, Bezostaja–1 6
14 Çeşit Triticum durum, Kızıltan–91 4
15 Çeşit Triticum durum, Çeşit 1252 4
Bilindiği gibi buğdaylarda üç değişik ploidi grubu yer almaktadır. Diploid (2x=14), tetraploid (4x=28) ve hekzaploid grup (6x=42). Ayrıca her ploidy grubuna üç farklı form ayırt edilmektedir. Bunlar; yabani, kavuzlu kültür ve çıplak kültür formlarıdır. Yabani formlar doğada kendiliğinden yetişen ve birçok özellikleri bakımından kültür buğdaylarından farklılık gösteren türleri içermektedir. Yabani buğdayları kültür buğdaylarından ayıran özelliklerin başında; başak ekseninin aşırı
kırılıcılığı, bindane ağırlığının düşüklüğü ve normal harmanlamayla tanelerinin çiçek kavuzlarından ayrılmayışı sayılabilir. Yabani formlar sadece diploid ve tetraploid grupta yer almaktadır. Bilinen yabani buğday türleri T. boeticum, T. urartu (2x=14); T. dicoccoides, T. araraticum ve, T. timopheevi (4x=28) dir. Hekzaploid gruptan yabani formlar bilinmemektedir. T. timopheevi bazı araştırıcılar tarafından kavuzlu kültür formu olarak bildirilmektedir.
Yeryüzünde diploid yabani buğdayların geniş bir yayılma alanı göstermelerine karşın, tetraploid yabani buğdayların yayılışı oldukça sınırlıdır.
Anadolu yabani buğdaylar açısından çok büyük bir potansiyel taşımaktadır. Bugün yetiştirilmekte olan makarnalık ve ekmeklik buğdayların gerçekte oldukça sınırlı bir genetik temele dayandığı düşünülebilir. Çünkü amphidiploid olan kültür buğdaylarının ataları, türler arası melezlerin büyük ölçüde kısır olmaları nedeniyle, çok az sayıda melezlerden kaynaklanmaktadır.
Bugün kültürü yapılan buğdaylar, onların genom vericisi olan yabani populasyonlardan yeni gen aktarımı olmamış ve bu yolla genetik varyebilitede herhangi bir artış sağlanmamıştır. Bunun da ötesinde, yeni varyebilite kaynakları girmediği gibi; sürekli seleksiyon sonucu başlangıçtaki varyebilitenin de oldukça daraldığı düşünülebilir. Kuşkusuz aynı durum hekzaploid buğdaylar için de geçerlidir. Bununla birlikte, buğdaylarda farklı ploidi basamaklarının bulunması nedeniyle buğday ıslahçıları şimdiye dek kültür buğdaylarının dışında bir genetik varyabilite kaynağı arama gereğini büyük ölçüde duymamışlardır. Ancak son zamanlarda bazı araştırıcılar, ıslah açısından önem taşıyan bazı özelliklerin yabani buğday populasyonlarındaki durumunu ve dayandığı potansiyeli incelemeye yönelmişlerdir. Şimdilik çeşitli hastalıklara dayanıklılık ve kalite ıslahı yönünden yabani buğdaylar üzerinde durulmaktadır (Yüce ve Turgut 1980).
Denemede kullanılan yabani buğday genotipleri; TUR 00687 T. monococcum var. boeticum, TUR 2453 T. dicoccum, TUR 03371 T. dicoccoides, TUR 03388 T. dicoccoides, TUR 03405 T. monococcum monococcum, TUR 03560 T. dicoccum, TUR 03562 T. dicoccum, TUR 01248 Ae. vavilovii, TUR 02554 Ae. tauschii, TUR 3354 Ae. speltoides ligustica, Aegilops cylindirica’dır. Kültür çeşitleri ise aşağıda verilmiştir.
Kızıltan–91, Tarla Bitkileri Merkez Araştırma Enstitüsünce 1991 yılında tescil ettirilen alternatif bir çeşittir. Orta Anadolu ve geçit bölgelerinde kurağa dayanıklılığı oldukça yüksek olan çeşidin, kışa ve soğuğa dayanıklılığı da iyidir. Gübreye reaksiyonu iyi olan çeşitte, gübre yeterli düzeyde yapıldığında dönme görülmez ve camsılık oranı artar. Tane dökmeyen ve harman olma yeteneği yüksek olan çeşidin bin dane ağırlığı 37-42 g dolayında olup, bulgurluk kalitesi oldukça yüksektir (Yürür 1994).
Çeşit 1252, Tarla Bitkileri Merkez Araştırma Enstitüsü tarafından 1991 yılında üretim izni alınıp 1999 yılında tescil ettirilen alternatif bir çeşittir. Yabancı otlarla rekabeti iyi olan çeşidin Orta Anadolu ve geçit bölgelerinde su stresinin olmadığı yörelerde verim potansiyeli çok yüksektir. Soğuğa ve kışa dayanıklılığı iyi olan çeşidin gübreye reaksiyonu yüksektir. Tane dökmeyen ve makarnalık kalitesi iyi olan çeşidin bin dane ağırlığı 38-42 g dolayındadır (Yürür 1994).
Gerek–79, Eskişehir Zirai Araştırma Enstitüsü tarafından 1964 yılında melezleme yöntemi ile elde edilmiştir. Kışlık gelişme tabiatlı, orta uzun boylu (90-100 cm), yeşil tüysüz yapraklı, orta uzunlukta ve seyrek başaklı, dış kavuz rengi kahverengi, kılçıklı, buğday sarısı renkte, orta uzunlukta ve genişlikte taneli, 1000 dane ağırlığı 36 g olup unsu yapı gösteren bir çeşittir (Yürür 1994).
Bezostaja-1, Rusya’da Tukyanen tarafından melezleme yoluyla elde edilmiş, ülkemize de introdüksiyon çeşidi olarak gelmiştir. Kışlık gelişme tabiatlı, yerli buğday çeşitlerimize göre daha kısa boylu, başağı orta uzunlukta ve sıklıkta, dış kavuzları tüysüz ve beyaz renkte, kılçıksız, başaktaki tane sayısı yüksek, kırmızı sert tane yapılı, bin dane ağırlığı 44.5 g olup kışa dayanma kabiliyeti yüksek, kurağa karşı dayanıklı değildir. Orta erkenci, yüksek verimli, sapı sağlam, kardeşlenmesi yerli çeşitlere göre azdır (Yürür 1994).
3.2. Metot
3.2.1. Denemenin kurulması
3.2.1.1. Saksıların hazırlanması
Deneme toprağı 4 mm çapındaki elekten geçirilip, kuru ağırlığa ulaşması sağlandıktan sonra 3 kg tartılıp plastik saksılara konulmuştur. Deneme tesadüf bloklarında faktöriyel deneme deseninde 3 tekerrürlü olarak kurulmuştur.
Denemede Fe sequestrin formunda ve 3 farklı dozda (Fe0 = Kontrol, Fe1 = 5
mg kg-1, Fe2= 25 mg kg-1) uygulanmıştır.
Bitkide normal bir gelişme sağlamak amacıyla bütün saksılara ekimden önce bitkinin ihtiyaç duyduğu makro ve mikro elementler de uygun seviyelerde 0.75 mg kg-1 B (H3BO3 halinde), 4 mg kg-1 Zn (ZnSO4.7H2O halinde), 10 mg kg-1 Mn
(MnSO4.H2O halinde), 1 mg kg-1 Cu (CuSO4.5H2O halinde), 100 mg kg-1 Mg
(Kiserit halinde), 80 mg kg-1 P ve 80 mg kg-1 K (15.15.15 gübresinden) çözelti şeklinde denemede bulunan tüm saksılara verilmiştir. Ekimden üç ay sonra üst gübrelemesi olarak 25 mg kg-1 N (%46 N içeren üre gübresinden) uygulanmıştır.
3.2.1.2. Tohumların saksıya ekilmesi
Denemede 15 buğday genotipi; noksan, yeterli, fazla olmak üzere 3 farklı Fe dozunda (0 mg kg-1 - 5 mg kg-1 - 25 mg kg-1), 3 tekerrürlü olarak yetiştirilmiştir (15 buğday çeşidi x 3 Fe dozu x 3 tekerrür = 135 saksı). 11.01.2005 tarihinde her saksıya, önceden tespit edilmiş olan çimlenme yüzdelerine göre uygun miktarda tohum, yüzeyden 2 cm derinlikte ekilmiş, çıkış tamamlandıktan sonra saksılarda bitki sayısı 5 olacak şekilde seyreltme yapılmıştır. 09.05.2005 tarihinde saksılarda bulunan 5 bitki fidesinden 2 tanesi yapılacak olan analizlerde kullanılmak üzere kardeşlenme
başlangıcında, ekimden 120 gün sonra başaklanma öncesi, fide numunesi olarak alınmıştır. Saksılar ihtiyaç durumuna göre haftada 2-3 kez tarla kapasitesinde saf su ile sulanmıştır ve hasada kadar rutin bakım işlemleri devam ettirilmiştir.
Deneme serada kurulmuştur ancak olumsuz sera şartlarından dolayı 17.03-27.05.2005 tarihleri arasında sera dışına çıkarılmıştır. Yabani buğdaylar birbirleriyle aynı anda olgunlaşma göstermediğinden dolayı hasat 14.06-17.07.2005 tarihleri süresince yapılmıştır.
Şekil 3.1. Denemeden genel bir görünüm (bitkiler 20. günde)
Şekil 3.3. Bitkiler başaklanma döneminde
Şekil 3.4. Hasat öncesi genel görünüm