Tatlı su ve deniz balıkları insanların beslenmesinde büyük önem taşımaktadır (Weatherley ve Gill 1989). Ülkemiz coğrafi yapısı ve iklim koşulları dikkate alındığında, deniz ve iç sularımızda çeşitli su ürünlerinin yetiştirilmesine ve geliştirilmesine imkân verecek kaynaklara sahiptir (Dönmez ve Tatar 2001). Balık etinin kalitesini belirleyen esas bileşenler proteinler ve lipidlerdir. Balığın canlı ağırlığının %70-80’ini su, %20-30’unu protein ve %2-12’sini lipidler oluşturmaktadır (Love 1970). Fakat balıklarda bu değerler türler arasında, aynı türün bireyleri arasında hatta aynı bireyin organları arasında faklılıklar gösterebilmektedir (Forss 1967).
Balıklar, lipidleri yağ dokusunda depo eden memelilerin aksine, daha çok iskelet kası ve karaciğer dokusunda depo ederler (Neuhaus ve Halver 1969). Balıklarda depolanan bu lipidlerin bir kısmı hayvanın fizyolojik durumu ve çevre koşullarında meydana gelen değişmelere göre gereksinim duyulan kısımlara nakledilir (Halver 1972, Johansson ve ark. 2000). Lipidlerin balıklar tarafından tamamıyla sindirildiği ve enerji kaynağı olarak karbohidratlara tercih edildiği ifade edilmiştir (Cho ve ark. 1985). Bu yağ asitlerinin başlıca kaynakları sudaki besin zincirinin ilk halkasını oluşturan planktonik organizmalardır.
Balık yağlarının komposizyonunu oluşturan üç yağ asidi tipi vardır. Doymuş yağ asitleri, doymamış yağ asitleri (MUFA) ve aşırı doymamış yağ asitleridir (PUFA) (Halver 1989). Balık yağları %20-30 oranında doymuş yağ asitlerini, %70- 80 oranında doymamış yağ asitlerini içerir. Balık yağlarındaki aşırı doymamış yağ asitlerinin miktarı %25-30 civarındadır. Su ürünlerinin yağlarındaki aşırı doymamış yağ asitleri genellikle omega-3 (ω-3) şeklindedir (Ackman 1988, Weatherley ve Gill 1989, Skorski 1990).
Balık yağlarındaki aşırı doymamış yağ asitlerinin zincir uzunlukları genelde 18 karbonun üzerindedir. Bitkisel ve hayvansal yağlarda zincir uzunluğu 18 karbonu geçen yağ asitleri miktarı %1-5 arasında iken balık yağında bu oran %33’tür hatta %50’ye kadar varabilir. Balık yağları 4, 5, 6 çift bağ içeren aşırı doymamış yağ
asitlerini de içerir. Özellikle ω -3 serisinden eikosapentaenoik asit EPA (C20:5) ve dokosaheksaenoik asit DHA (C22:6) balık yağına özgüdür. Balık yağlarındaki aşırı doymamış yağ asitleri ω-6’dan ziyade ω-3 şeklindedir (Göğüş 1988).
Balık yağının bileşimi içinde yer alan karbon sayısı ve doymamışlığı yüksek olan aşırı doymamış yağ asitleri hücre zarlarının yapısında yer alarak hücre zarlarının akışkanlığı ve geçirgenliğinde rol oynarlar. Eikosanoidler, EPA gibi yirmi karbonlu aşırı doymamış yağ asitlerinden türerler. Eikosanoidler, eikosa-(20 C) polienoik yağ asitlerinin türevi olan bileşiklerdir ve yerel hormon olarak görev yaparlar. Ayrıca bu yağ asitleri derideki geçirgenlik bariyerinin devamında, kolesterol metabolizmasında ve taşınmasında önemli görevleri vardır (Steffens 1997). Yine balık yağlarının bileşiminde bulunan (C22:6) DHA, hücre membranının fonksiyonel bütünlüğü ve temel yapısal özelliklerin devamı için gereklidir (Gunasekara ve ark. 1999).
Balıklar lipidleri depo etme durumuna göre; yağlı balıklar, yarı yağlı balıklar ve yağsız balıklar olarak sınıflandırılır. Yağlı balıklar lipidleri kas dokularında depo ederken, yağsız balıklar lipidlerin çoğunluğunu karaciğer veya karın bölgesinde depo ederler, kas dokusu yağsızdır (Huss 1988).
Balıklarda lipid ve yağ asidi bileşimi esas olarak türlere, mevsim ve aylara özellikle üreme mevsimine, beslenme ortamına ve besin yapısına, suyun sıcaklığına ve kirliliğine, türün kültür ya da yabani form olup olmadığına ve vücut kısımlarına göre değişiklik göstermektedir (Ackman 1967, Deng ve ark. 1976, Agren ve ark. 1987, Cai ve Curtis 1989).
Kalorik fonksiyonlarının yanında yağlar, vücut yapısının gelişmesi için gerekli ve dışarıdan alınması zorunlu olan yağ asitlerini de içerirler. Vücudun üretemediği ve mutlaka besinler yoluyla dışarıdan alınması gereken yağ asitlerine esansiyel yağ asitleri adı verilir. Yağsız dietle beslenen fareler üzerinde yapılan araştırmada; büyümenin gecikmesi, böbrek fonksiyon bozuklukları, cilt sorunları, üreme fonksiyon bozuklukları gibi rahatsızlıklara rastlanılmıştır. Fakat bu araştırma, sorunun yağ asidi eksikliğinden değil, linoleik asit (ω-6) adlı yağ asidi eksikliğinden kaynaklandığını göstermiştir. Araştırmalar devam ettikçe, linolenik asit (ω-3) ve arakidonik asidin (ω-6) de esansiyel olduğu saptanmıştır. Yapılan birçok araştırma,
ise bu oran 1.2/1’dir. Girit adasında yaşayanların dietindeki yüksek miktardaki ω-3 yağ asitleri; sardalya, ringa gibi balıkları ve yumurtaları yemelerine bağlıdır (ω-3 ve ω-6 yağ asitlerinin dengeli alınmasının sayısız faydalar getirdiğini göstermektedir (Burr ve Fehily 1990). Hücre membranının fleksibilitesi, akışkanlığı esansiyel yağ asitlerinin membrandaki miktarına bağlıdır. Ayrıca esansiyel yağ asitleri enerji sağlar, vücut ısısının korunmasına yardımcı olur (Hawkins 1997).
Balık yağları başta kalp-damar hastalıkları olmak üzere ölüme sebebiyet verebilen birçok hastalığın iyileştirilmesinde rol oynamaktadır (Steffens 1997). Fazla miktarda balık tüketen toplumlarda kardiovasküler hastalıklar ve depresyon daha az görülmektedir (Leaf ve Weber 1988). Dünyanın çeşitli yerlerinde diet ve kardiovasküler hastalıkları hakkında yapılan çalışmalar Japonya ve Girit adasında yaşayan toplumların kardiovasküler sistem hastalıkları açısından daha düşük ölüm riskine sahip oldukları gösterilmiştir. Girit adasında yaşayan toplumların dieti yağlardan zengin, ancak sature yağlardan fakirdir. Japon toplumunun dieti ise hem total yağlardan hem de sature yağlardan fakirdir. Japonların dietinde ω-6/ω-3 oranı 2-4/1 iken, Giritlilerin dietinde Tornaritis 1993). Girit adasındaki tavukların yumurtalarında ω-6 /ω-3 oranı 1.3/1 iken Amerika’da süpermarketlerde satılan tavuk yumurtalarında bu oran 19.4/1’dir (Charalambakis ve Divanach 1993).
Balık yağında bulunan ω-3 grubu yağ asitlerinden eikosapentaenoik ve dokosaheksaenoik asitler platelet yapışkanlığını azaltıp birikimine engel olarak, kan viskozitesinin, trigliserit ve kolesterol seviyesini azaltarak damar lümeninin zarar görmesini ve tıkanmasını önlemektedir (Li ve Steiner 1990, Weber ve Leaf 1990). Kalp-damar hastalığı olan 852 orta yaşlı erkek yirmi yıl süre ile takip edilmiş ve haftalık yenen balık miktarı ile bu hastalıktan ölüm oranı arasında ters bir ilişki olduğu saptanmıştır. Her gün 30 g balık tüketiminin kalp-damar hastalıklarından ölümü %50 azalttığı belirtilmiştir (Leaf ve Weber 1988). Balık ve balık yağı tüketiminin kardiovasküler sistem bozukluklarından kaynaklanan ölümleri azalttığı ortaya konmuştur (Jonkers ve ark. 2000).
Balık yağlarının sedef ve ekzema gibi çeşitli cilt hastalıklarını (Ziboh 1990); meme, pankreas, barsak ve prostatik tümörlerin gelişmesini önlemesi (Kanders ve Kowalchuk 1990) gibi birçok klinik etkilerinin olduğu belirtilmiştir. Balık yağının bu
etkilerinin prostaglandinler veya benzeri bileşiklerin üretimindeki değişimlerin bir sonucu olarak meydana geldiği ileri sürülmüştür (Galli ve Butrum 1990).
ω-3 yağ asitlerinin sentezi insanlarda oldukça yavaştır. İnsan yaşlanınca ve belli hastalıklarda desatüre aktivitesi kısmen kaybolur. Bundan dolayı ihtiyaç olan bu önemli yağ asitlerinin dışarıdan besinle alınması zorunludur. Balık yağı bu gerekli yağ asitlerini içermektedir. Balık yağı en zengin ω-3 yağ asitleri kaynağı olduğu bildirilmiştir.
İnsan beslenmesinde önemli rol oynayan balıklardan yeterince yararlanmak için bunların besinsel değerlerinin araştırılması önem taşımaktadır. Ülkemiz coğrafi yapısı ve iklim koşulları bakımından, deniz ve tatlı sularımızda çeşitli su ürünlerinin yetiştirilmesine ve geliştirilmesine imkân verecek kaynaklara sahiptir. Bu çalışma da; ülkemizde bol olarak bulunan, üretim kültürüne uygun olan çipura (Sparus aurata L.)’nın total lipid ve total yağ asidi bileşiminin mevsimsel değişimi araştırılmıştır.
2. KAYNAK ARAŞTIRMASI
2.1. Çipura (Sparus aurata)’nın Biyolojisi
Ülkemizde Ege ve Akdeniz’de bol bulunan bu balık türü Marmara’da seyrek bulunmaktadır. Yaz aylarında sığlarda, kış aylarında ise 35-40 m derinliklerde yaşarlar. 2 yaşın üzerindekiler daha da derinlere inmektedirler. Boyları 25-35 cm kadar olabilmektedir. Ağırlıkları ise 0.5-3 kg kadardır. En çok 60 cm ve 6 kg kadar olabilmektedirler. Karnivordurlar, küçük kabukluları, balıkları ve diğer hayvansal canlıları yerler. Üremeleri Ekim –Aralık aylarında olur. 100000- 150000 yumurta bırakırlar. Etleri çok lezzetlidir. Ayrıca üretim kültürüne uygunluğu nedeniyle ekonomik değerleri çok yüksektir (Akşiray 1954).
2.2. Balıklarda Yağ Asidi Bileşimi
Yağ asitleri, içerdikleri karbon atomlarının sayısı, zincir uzunlukları, karbon atomları arasındaki çift bağların sayıları ve doymamışlık derecelerine göre sınıflandırılırlar. Bunlar doymuş, doymamış ve aşırı doymamış yağ asitleridir. Balık yağları %20-30 oranında doymuş yağ asitlerini, %70-80 oranında doymamış yağ asitlerini içerir. Balık yağlarındaki aşırı doymamış yağ asitlerinin oranı ise %25-30’dur. Su ürünlerinin yağlarındaki aşırı doymamış yağ asitleri genellikle ω-3 şeklindedir. ω-6 yağ asitleri ise toplam yağ asitleri oranının %1-3’ünü oluşturmaktadır (Ackman 1988, Weatherley ve Gill 1989, Skorski 1990).
Su ürünlerindeki yağlar, bitki ve hayvan yağlarına göre daha kompleks yapıdadırlar. Karbon zinciri uzunluğu C 14 ve C 24 arasındadır, hatta C 12 ile C 26 bile bulunabilmektedir. C 14 ve C 16 tekli doymamış bağ içerirken C 20 ve C 22 yağ asitleri; 4, 5 ve hatta 6 çift bağ içerirler (Keskin 1981). Her ne kadar balığın yağ asidi bileşimi çok sayıda yağ asidinden oluşuyorsa da bunlardan sadece birkaç tanesi, total yağ asidi bileşiminin genellikle %70-80 gibi önemli bir yüzdesini oluşturmaktadır. Doymuş yağ asitlerinden en yüksek yüzdelerde bulunanları C 16:0 palmitik asit,
C14:0 miristik asit ve C 18:0 stearik asitlerdir. Tekli doymamış yağ asitlerinden C16:1 palmitoleik asit ve C 18:1 oleik asidin yüzdeleri oldukça yüksektir. Aşırı doymamış yağ asitlerinden ise özellikle C 20:5 eikosapentaenoik asit (EPA) ve C22:6 dokosaheksaenoik asit (DHA) oldukça yüksek yüzdelerdedir (Gunstone 1986). Balık etindeki yağların kaynağı vücuda besinlerle alınan yağlar, karbohidratlar ve proteinlerdir. Vücuda alınan bu besin maddelerinin ihtiyaç fazlası organ ve dokularda yağ şeklinde depo edilmekte, bu durum ise balık etinin yağ asidi düzeylerini etkilemektedir (Boggio ve ark. 1985, Csengeri ve ark. 1986, Henderson ve ark. 1998, Domaizon ve ark. 2000, Bell ve ark. 2001, Kiessling ve ark. 2001). Balıklardaki doymuş yağ asitleri yem kökenli olabilir, bağırsaktaki bakteriler tarafından oluşturulur veya mevcut bakterilerden absorbe edilir. Başlıca doymuş yağ asitleri miristik (C 14:0), palmitik (C 16:0) ve stearik (C 18:0) asitlerdir. Tekli doymamış yağ asitlerini ise besinlerden biyosentez yoluyla sağlarlar ve C 18:1’den C22:1’e kadar tekli doymamış yağ asitlerini uzatma yeteneğine sahiptirler. Aşırı doymamış yağ asitleri içerisinde yaygın olarak bilinen en yüksek doymamışlık derecesi C 22:6 dokosahekzaenoik asittir (Halver 1989).
Huss (1988), balıkları yağlı, yarı yağlı ve yağsız olmak üzere sınıflandırmıştır. Yağlı balıklara ringa, uskumru, tuna, çaça gibi balıklar, yarı yağlı balıklara lüfer, tekir gibi balıklar, yağsız balıklara ise pisi, morina gibi balıklar örnek olarak verilebilir. Yağlı balıklar lipidleri kas dokuda depo ederken, yağsız balıklar ise lipidlerin çoğunluğunu karaciğer veya karın bölgelerinde depo ederler. Yani kas dokuları yağsızdır (Huss 1988). Su ürünlerindeki yağların yağ asitleri kompozisyonlarının farklı olması bazı faktörlere bağlıdır. Bu faktörler beslenme şekli, coğrafik şartlar, çevre sıcaklığı, mevsimlere göre avlanma, vücut uzunluğu, cinsiyet, tür ve yağ içerikleri olarak sayılabilir.
Uysal (2000), Eğridir Gölü’nde yaşayan sudak (Stizostedion lucioperca) balıklarının kas dokularında yağ oranının yıl içinde hiçbir zaman %1’i geçmediği ve yağsız balıklar sınıfına girdiğini tespit etmiştir. Karaciğer yağ içeriğinin kas dokusu yağ içeriğinin fazla olduğunu saptamıştır.
Farklı araştırıcılar balık dokusunda en yüksek oranda bulunan yağ asidi çeşidinin palmitik asit olduğunu tespit etmişlerdir. Brezilya’nın güney bölgesinde on yedi balık türünün yenilebilir kısımlarının yağ asidi bileşimi araştırılmış ve bütün türlerde palmitik asidin en çok bulunan doymuş yağ asidi olduğu bildirilmiştir. Bu araştırmada total doymuş yağ asitlerinin %50-70’ini palmitik asidin oluşturduğu saptanmıştır (Andrade 1995).
Wang ve ark. (1990), Superior Gölü balıklarının yağ asidi bileşimlerini inceledikleri araştırmada, kas dokusunda en çok bulunan yağ asidinin palmitik asit olduğunu belirlemişlerdir. Doymuş yağ asitlerinin %68-79’unu palmitik asidin oluşturduğunu saptamışlardır.
Sinclair (1982), Avusturalya’nın kuzeybatı sahillerindeki on balık türünün yağ asidi bileşimini incelemiş ve araşidonik asidin en çok bulunan ω-6 yağ asidi olduğunu saptamıştır. Bu verilere dayanarak Güney Yarım Küre’deki denizsel besin zincirinin ω-6 yağ asitlerince zengin olduğunu ileri sürmüştür.
Gibson (1983), 24 balık türünün yağ asidi bileşimini araştırmış genel olarak ω-3 yağ asitlerinin ω-6 yağ asitlerinden daha çok bulunduğunu bildirmiştir.
Gibson ve ark. (1984), Malezya balıklarından 22 türün total lipidlerini analiz etmişlerdir. Bu balıklarının yağ asidi bileşimlerinin genellikle yüksek seviyelerde doymuş yağ asitlerini ve doymamış yağ asitlerini değişik oranlarda içerdiğini tespit etmişlerdir. Doymuş yağ asitleri içinde palmitik asit ve stearik asit fazla miktarda bulunmuştur. Kuzey yarım kürenin soğuk sularında yakalanan balıklara benzeyen Malezya balıklarının, araşidonik asidi %2-12; eikosapentaenoik asidi %1-13 ve dokosaheksaenoik asidi %6.6-40.4 arasında ihtiva ettiklerini belirtmişlerdir.
Ünlüsayın (1999), gökkuşağı alabalığı, yılan balığı ve sudak balıklarının yağ asidi bileşimini araştırmıştır. Yılan balıkları yağlarının %16.09 doymuş, %67.30 doymamış ve %16.61 aşırı doymamış yağ asitleri olduğu tespit edilmiştir. Gökkuşağı alabalıklarının yağ asidi kompozisyonu, %19.79 doymuş, %57.77 doymamış, %4.14 aşırı doymamış yağ asitlerinin meydana getirdiği saptanmıştır. Sudak balıklarının
yağ asidi analizinde doymuş yağ asitlerinin oranı %40.14, doymamış yağ asitleri oranı %41.31 ve aşırı doymamış yağ asitleri oranı ise %17.97 olarak tespit edilmiştir.
Sağlık (1994), denizlerimizde yaşayan ve besin olarak tüketilen on üç balık türünün total lipid ve yağ asidi bileşimlerini araştırmış ve türler arasında oldukça fazla farklılıklar olduğunu tespit etmiştir. Araştırma sonucunda total lipid içeriği en fazla sardalya balığında (%16.96), en az kılıç balığında (%0.54) bulunmuştur. Balık dokusundaki hiçbir kimyasal madde türler arasında bu kadar değişim göstermemektedir. Ayrıca bu araştırma sonucuna göre kılıç balığı en çok doymuş yağ asidi içeren tür, aşırı doymamış yağ asitlerini en fazla içeren tür ise hamsi olarak tespit etmiştir (%36.4).
Kıraç (2004) Konya’da tüketilen 15 tür balığın yağ asidi bileşimlerini incelediği araştırmasında, bu balıkların; %37.93 doymuş yağ asidi, %33.07 doymamış yağ asidi, %29.00 aşırı doymamış yağ asidi içerdiklerini göstermiştir. Çalışmasında en yüksek doymuş yağ asidi yüzdesi %44.16 ile çinakop balığında, en yüksek doymamış yağ asitleri yüzdesi %38.87 ile barbun balığında, en yüksek aşırı doymamış yağ asitleri yüzdesi ise %38.64 ile sardalya balığında olduğunu tespit etmiştir.
Balığın yağ asidi bileşimi mevsimlere bağlı olarak değişim göstermektedir. Kandemir (1999), yapmış olduğu çalışmada gökkuşağı alabalığının (Oncorhynchus mykiss) total lipid ve yağ asidi miktarlarının sabit kalmayarak mevsimlere, mevsimin aylarına ve organlara göre değiştiğini ortaya koymuştur.
Deniz ve tatlı su balıklarının yağ asidi bileşimi farklılıklar göstermektedir. Tatlı su balıklarında 16–18 karbonlu, deniz balıklarında ise 20–22 karbonlu yağ asitleri daha fazladır. Tatlı su balıkları ω-6, deniz balıkları ω-3 yağ asitleri bakımından zengindir (Ackman 1967). Balıkların yağ asidi bileşimine etki eden en önemli faktör dietinde bulunan besinlerin ihtiva ettiği yağ asidi bileşimleridir (Brenner 1989, Crawford ve Marsh 1989).
Deniz balıklarının yağındaki yağ asidi bileşimini deniz planktonlarının yağ asidi bileşimi oluşturur (Kelly ve ark. 1959, Kayama ve ark. 1963, Sargent ve
Henderson 1980). Deniz balıklarının yiyeceklerindeki planktonlar düşük seviyede ω-6 aşırı doymamış yağ asitleri içerirken yüksek oranda EPA ve DHA gibi ω-3 aşırı doymamış yağ asitlerini içerirler (Sargent ve ark. 1989, 1995). Tatlı su balıkların yağ asidi bileşiminde linoleik ve arakidonik gibi ω-6 yağ asitlerinin oranı yüksektir. Bunun sonucunda ω-3/ω-6 oranı tatlı su balıklarında deniz balıklarınkinden daha düşüktür (Henderson ve Tocher 1987, Steffens 1997).
Tatlı su balıklarında besinle alınan linoleik ve linolenik asitler zincir uzaması mekanizması ve çift bağ ilavesiyle doymamışlık dereceleri arttırılır (Boggio ve ark. 1985, Halver 1989). Bu zincir uzaması ve çift bağ ilavesi genellikle hücre membranındaki yağ asitleri için meydana gelir (Hagen ve ark. 1995). Balıklardaki bu özelliğin amacı, besin değeri az olan yiyecekleri besin değeri daha yüksek olan yiyeceklere çevirmektir (Ahlgren ve ark. 1999).
Balıkların beslenme şekli yani karnivor, herbivor veya omnivor oluşu da balıklardaki yağ asidi bileşimine etki etmektedir. Balıkların yağ asidi bileşiminde karakteristik olarak bulunan EPA ve DHA gibi aşırı doymamış yağ asitlerinin yüzdeleri özellikle karnivor beslenen balık türleri ile alg ve yüksek bitkilerle beslenen herbivor balık türleri için daha yüksek iken omnivor türlerde nispeten daha ılımlı seviyelerdedir (Roy ve ark. 1999). Tatlı sularda et yiyen balık türleri fazla yaygın olmadığı için tatlı sularda yaşayan balıkların yağ asidi bileşimi nispeten birbirlerine benzerlik gösterir (Ahlgren ve ark. 1994).
Balıkların yağ asidi bileşimi, balığın doğal ortamlarda yetişmiş ya da kültüre edilmiş olmasına göre de değişiklik gösterir. Doğal yiyeceklerde DHA seviyesi düşük olmasına rağmen doğal ortamda yetişmiş tatlı su balıklarının etinde DHA oranı nispeten daha fazladır. Perca fluviatilis, Coregonus albula, gibi iki göl formu ve Salmo gairdneri gibi kültür formu olan üç tatlı su balığının kas lipidlerinin yağ asidi seviyeleri araştırılmış ω-3 aşırı doymamış yağ asitlerinin nisbi miktarı, Coregonus albula ve Perca fluviatilis’de kültür formu olan Salmo gairdneri’den daha yüksek olarak bulunmuştur (Agren ve ark. 1987).
Balıkların yağ asidi içeriğine etki eden bir diğer faktör ise üreme durumları ve cinsiyet özellikleridir (Fremont ve Marion 1982, Johnson ve Casillas 1991,
MacFarlane ve ark. 1993). Balıklar yeterli besin bulduklarında üremelerini ve yağ depo etme periyodlarını kontrol edebilmektedirler. Yağ depo etmeleri ortamdaki besin miktarına bağlıdır. Besinin az bulunduğu yerlerde yağ asidi değişimi yıl boyunca az, besinin bol olarak bulunduğu yerlerde ise yıllık değişim daha belirgindir. Gonadların gelişim evresinde fazla miktarda enerjiye gereksinim duyulduğundan bu dönemde bol besin bulunması gerekmektedir. Balıklar gonadların gelişim evresinde ve üreme periyodunda gereksinin duydukları enerjiyi depo ettikleri yağlardan sağlamaktadırlar (Hayashi ve Takagi 1977).
Tatlı su balıklarından olan Aplodinotus grunnies, Coregonus artedii, Lota lota ve Alosa pseudoharengus türleri üzerinde yapılan çalışmada, üreme periyodu öncesinde lipit miktarının arttığı ve üreme periyodu sonunda da azaldığı rapor edilmiştir (Ackman 1967).
Balıklar memeli hayvanların aksine yağlarını yağ dokuda değil kas ve karaciğer dokularında depo ederler. Bu organların önemi balık türlerine göre farklılıklar göstermektedir. Hareketli balıklarda, lipidler daha çok kas dokuda; hareketsiz ve suyun dibinde yaşayan balıklarda, karaciğer dokusunda depo edildiği belirtilmiştir (Neuhaus ve Halver 1969).
2.3. Balık Yağ Asidinin Mevsimsel Değişimi
Balıklar poikloterm canlılardır. Yapılan çalışmalar, çevre sıcaklığının balıkların yağ asidi metabolizması üzerine doğrudan etkili olduğunu göstermiştir. Balık membranlarının biyokimyasında çevre sıcaklığı ve dietindeki yağ asidi bileşiminin etkisi çok önemlidir (Roy ve ark. 1999). Suyun sıcaklığının düşmesi, yapısal lipidlerindeki yağ asitlerinin karbon sayılarının yükselmesine ve doymamışlığının artmasına neden olmaktadır (Willams ve Hazel 1992). Balıklar için çevre sıcaklığını etkileyen en önemli etken mevsimsel değişimlerdir. Buna bağlı olarak balığın yağ asidi bileşimi mevsimlere bağlı olarak değişim göstermektedir.
Balıklar, yağlarını kas ve karaciğer dokularında depo ederler. Depo edilen bu lipidlerin bu organlardan gonadlar gibi vücudun diğer ihtiyaç olan yerlerine taşındığı
belirtilmiştir (Jangaard ve ark. 1967). Bütün balıklar üreme periyodundan sonra, üreme faaliyetlerinde kaybettikleri kondisyonlarını tamamlamak için yoğun bir beslenme periyoduna girerler. Kışa ve üremeye hazırlık nedeniyle, yaz ve sonbahar aylarında metabolizma olayları daha hızlıdır. Yazın lipidlerin depo edilmeleri kış mevsimine hazırlık ve üreme faaliyetleri için gereklidir. Gonadların gelişmeye başlamasıyla birlikte kas, karaciğer ve diğer dokulardaki depo lipidler gonadlara mobilize olmakta, gonadların lipit miktarı artarken kas ve karaciğer lipid içeriği de azalmaktadır. Ayrıca balıklar poikloterm canlılar olmaları nedeniyle beslenme olaylarında çevre sıcaklığı önemlidir (Agren ve ark. 1987).
Uysal (2000), Eğirdir Gölü’nde yaşayan sudak balıklarının kas, karaciğer ve gonadlarının total lipid, total yağ asidi içeriğinin mevsimsel değişimini araştırmıştır. Çalışmasında kullanılan sudakların, üreme periyodundan sonra ağırlıklarının, total lipid ve total yağ asidi oranlarının artmasında, su sıcaklığının artmasıyla birlikte besin maddelerinin çoğalmasının ve balığın biyolojik ihtiyaçlarından dolayı yoğun bir beslenme periyoduna girmesinin etkili olduğunu belirtmiştir. Sudakların yağ asidi metabolizmasının çevre sıcaklığından ziyade beslenme, gonad gelişimi ve üremeye bağlı olarak değiştiğini belirtmiştir.
Kandemir (1999), Derbent Baraj gölünde kültürü yapılan gökkuşağı alabalığının kas dokusu ve karaciğerinde total lipid total yağ asidi miktarı ile yağ asidi bileşimlerini aylar ve mevsimlere göre incelemiştir. Çalışma sonucunda, total lipid miktarının, kas dokusunda sonbahar, karaciğerde ise yaz aylarında maksimum seviyede olduğunu saptamıştır. Total yağ asidi miktarının ise kas dokusunda yaz, karaciğerde ise kış aylarında diğer aylardan daha yüksek düzeyde olduğunu tespit etmiştir. Doymamış yağ asitleri analizinde aşırı doymamış yağ asitleri, doymamış yağ asitlerinden yüksek bulunmuştur. Ayrıca ω-3 yağ asitlerinin ω-6 yağ asitlerinden fazla olduğunu tespit etmiştir.
Öztürk (2003), Beyşehir Gölü’nde yaşayan kadife balığının yağ asitlerinin mevsimsel değişimini incelediği çalışmasında, doymuş yağ asitleri toplamı %48,32 ile yaz mevsiminde en fazla olarak tespit edilmiştir. Doymamış yağ asitleri ise ilkbahar mevsiminde %32,02 olarak bulunmuştur. Aşırı doymamış yağ asitleri toplamı ise %39,44 ile kış mevsiminde en yüksek olarak ölçülmüştür. Sağlık
açısından öneli olan yağ asitlerinden EPA ve DHA’ da en yüksek kış mevsiminde ölçülmüştür.
Gallagher ve ark. (1991), Micropogonias andulatus, Mugil cephalus ve Paralicthys dentatus’un kaslarındaki yağ asidi bileşimini bir yıl süre ile incelemiş ve M. andulatus, P. dentatus ‘un kas dokularındaki aşırı doymamış yağ asidi içeriğinin ağustos ayından ocak ayına doğru düzenli olarak arttığı halde M. cephalus’un kas dokusunda önemli bir değişme olmadığını saptamıştır.
Mogan Gölü’ndeki Cyprinus carpio’nun kas dokusu yağ asitlerinin değişimi eşeye ve mevsime bağlı olarak araştırılmış ve her iki eşeyin kas dokusu yağ asidi bileşiminin kantitatif yönden faklı olmadığı belirlenmiştir. Buna rağmen uzun zincirli aşırı doymamış yağ asitlerinin miktarında önemli düzeyde değişmeler gözlenmiş, bu değişimlerin nedenlerinin gonad gelişimi ve üreme periyodu ile ilgili olabileceği ifade edilmiştir (Akpınar 1987).
Yılmaz ve ark. (1996), Capoeta capoeta umbla’nın dişi bireylerinde, total lipit miktarının ekim ayında ve erkek bireylerde de kasım ayında diğer aylara göre belirgin düzeyde yüksek olduğunu ifade etmişlerdir. Bu farklılığın nedeninin beslenme ve üreme gibi bazı fizyolojik faaliyetlerden kaynaklandığını ileri sürmüşlerdir. Nevsome ve Leduc (1975), üreme dönemi boyunca balıkların gerekli olan enerji ihtiyaçlarını kas dokusundaki lipidlerden sağladıklarını ifade etmişlerdir.
Yapılan çalışmalara göre; balıklardaki yağ asidi miktarlarının ve yağ asidlerinin değişimlerini çevre sıcaklığından ziyade beslenme, üreme periyodu ve gonad gelişimi gibi fizyolojik faaliyetler etkilemektedir.
3. MATERYAL VE METOT
3.1. Balıkların Temin Edilmesi
Araştırmada kullanılan çipura (Sparus aurata) numuneleri Mersin yöresinden elde edilmiştir. Balığın yağ asidi bileşimine yaşadığı ortamın yani su sıcaklığının etkisinin olup olmadığını araştırmak için numuneler dört mevsim ayrı ayrı toplanmıştır. Numunelerin alınması her mevsimin ortasına denk gelen aylarda; kış için ocak ayında, bahar için nisan ayında, yaz için temmuz ayında, sonbahar için ekim ayında gerçekleştirilmiştir. Her mevsimde üçer balık alınmıştır. Eşeysel olgunluğunu tamamlamış, ortalama kabul edilen ağırlıkta bireyler toplanmıştır. Numuneler alınırken cinsiyet ayrımı yapılmamıştır.
3.2. Total Lipid Miktarlarının ve Yağ Asidi Bileşimlerinin Tespit Edilmesi
Balık numunelerinin her birinden sol pektoral yüzgeci ile dorsal yüzgeci arasındaki bölgeden alınan kas dokusu tartılmış ve analiz için Süleyman Demirel Üniversitesi Deneysel ve Gözlemsel Öğrenci Araştırma Merkezine gönderilmiştir. Dondurulan numuneler, buz kalıpları arasında, çözünmeden teslim edilmiştir. Total lipid miktarlarının tespiti ve yağ asidi bileşimlerinin analizlerinin tamamı bu araştırma merkezi tarafından yapılmıştır. Bu merkezde, total lipid miktarlarının tespit edilmesi için 5 g balık eti alınmış ve üzerine 30 ml. kloroform-metanol (2:1) karışımı eklenerek homojenize edilmiş ve süzülmüştür. Çözücünün uçurulması ile sabit tartıma gelmesi beklenen numunenin total lipid miktarı bulunmuştur. Elde edilen yağlar %0.5’lik sodyum metilat içeren türevlendirici karışım içinde bir gece bekletilmiş ve türevlendirilmiştir. Türevlenen kısım hekzan içinde sisteme verilmiştir. Yağ asidi metil esterlerinin analizi için QP 5050 GC/MS (GC: Shimadzu 17 A, MS: Quadrapole Dedektör) kullanılmıştır. Çalışma şartları ise aşağıdaki gibi ayarlanmıştır;
Enjeksiyon bloğu : 250 ºC Dedektör : 250 ºC Akış Hızı (psi) : 10 Dedektör : 70 V İyonlaştırma Türü : EI Kullanılan Gaz : Helyum
Kullanılan Kolon : Cp WAX 52 CB 50 m 0.32 mm 1.2 µm
Sıcaklık Programı : 60 ºC’ den 220 ºC’ e dakikada 2 ºC’ lik artışla ulaşıyor. 220 ºC’ de 20 dk. bekliyor.
Kütüphaneler :Wiley. Nist. Tutor
3.3. Verilerin Değerlendirilmesi
Her mevsim için alınan üç ayrı tekrara ait verilerin istatiksel analizi SPSS 10.0 programı kullanılarak One Way ANOVA ve Tukey Testi ile yapılmıştır. Üçten az veriye sahip yağ asitlerinin karşılaştırılmasında ise T Testi kullanılmıştır (SPSS 10.0).
4. SONUÇLAR
S. aurata’nın total lipid miktarları mevsimlere göre değişiklik göstermektedir (Tablo 1). Sonbahar mevsiminde %3.71 ile en yüksek değerde iken, kış mevsiminde %2.05 ile en düşük seviyede bulunmuştur.
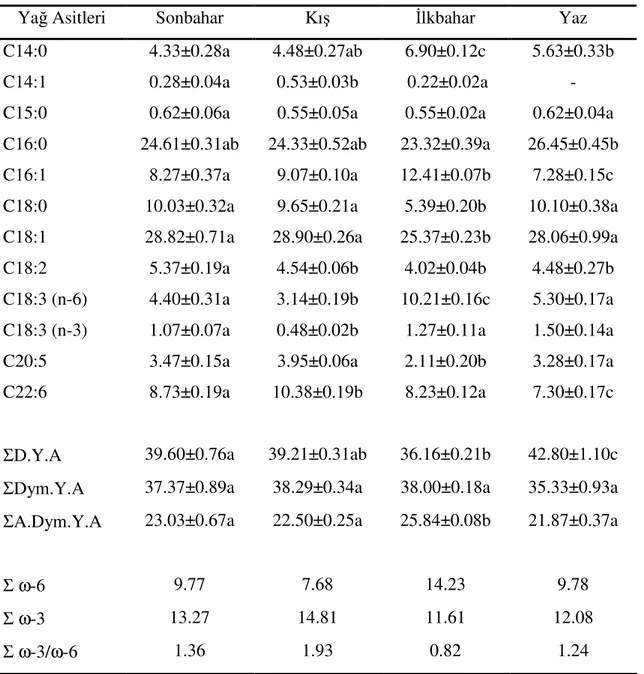
Mersin yöresinde yaşayan çipura balığı, Sparus aurata L.’nin total yağ asidi bileşiminde mevcut olan yağ asitlerinin karbon sayılarının 14-22 arasında değiştiği görülmüştür (Tablo 2). Toplam 12 değişik yağ asidi balığın total yağ asidi bileşimini oluşturmuştur.
Genel olarak C 18:1 oleik asit, C 16:0 palmitik asit, C 18:0 stearik asit, C 16:1 palmitoleik asit ve C 22:5 dokosahekzaenoik asit yüzdelerinin balıkların total yağ asidi bileşiminde yüksek yüzdelerde bulunduğu görülmüştür. Bu yağ asitlerinin toplam yüzdeleri mevsimlere göre %74.72-82.33 arasında değişiklik göstermiştir.
Balıkların total yağ asidi bileşimlerindeki doymuş, doymamış ve aşırı doymamış yağ asitlerinin yüzdeleri toplamlarının mevsimlere bağlı olarak değişiklikler gösterdiği belirlenmiştir. Mevsimlere göre doymuş yağ asitlerinin yüzdeleri toplamının %36.16-42.80, doymamış yağ asidi yüzdeleri toplamın %35.33-38.29, aşırı doymamış yağ asidi yüzdeleri toplamının %21.87-25.84 arasında olduğu görülmüştür.
4.1. Sparus aurata’ nın Mevsimlere Göre Total Lipid Miktarlarının Değişimi
S. aurata’nın mevsimsel total lipid miktarındaki değişiklikler Tablo 1’ de verilmiştir. S. aurata’nın total lipid miktarı sonbahar mevsiminde %3.71, kış mevsiminde %2.05, ilkbahar mevsiminde %3.15 ve yaz mevsiminde %3.49 olarak bulunmuştur. Buna sonuca göre sonbahar mevsiminde total lipid miktarı en yüksek, kış mevsiminde ise en düşük seviyededir.
Tablo 1. S. aurata’ nın mevsimlere göre total lipid miktarları Mevsim Ort ± SH * Sonbahar 3.71±0.21a Kış 2.05±0.06b İlkbahar 3.15±0.07c Yaz 3.49±0.07ac
*Aynı sütunda aynı harfle gösterilen değerler birbirinden farklı değildir (p>0.05).
4.2. Sonbahar Mevsiminde Sparus aurata’nın Total Yağ Asidi Bileşimi
S. aurata’nın sonbahar mevsimi total yağ asidi bileşimi Tablo 2’de verilmiştir. Bu sonuçlara göre S. aurata’nın total yağ asidi bileşiminde en yüksek yüzdeye sahip olan yağ asidinin %28.82 gibi büyük bir yüzdeyle oleik asit olduğu görülmüştür. Oleik asidin yanı sıra palmitik asidin ve stearik asidin de büyük yüzdelerde olduğu tespit edilmiştir. Bu üç yağ asidine ilaveten C 22:6 dokosahekzaenoik asit, palmitoleik asit, C 18:2 linoleik asit, C 18:3 (ω-6) γ-linolenik asit ve C 14:0 miristik asit de yüksek oranlarda bulunan yağ asitleridir. Yüksek oranlarda bulunan bu yağ asitlerinin toplam yüzdeleri yağ asidi bileşiminin %94.56’sını oluşturmuştur.
Bu sekiz yağ asidinin dışında kalan eikosapentaenoik asit (EPA), C 18:3 (ω-3) linolenik asit, C 15:0 pentadesilik asit ve C 14:1 miristoleik asitlerinin yüzdeleri toplamı bileşimin ancak %5.44’ünü oluşturmuşlardır.
Sağlık açısından varlığı önemli olan yağ asitlerinden eikosapentaenoik asit (EPA) %3.47, dokosahekzaenoik asit (DHA) ise %8.27 olarak bulunmuştur.
Sonbahar mevsiminde S. aurata’nın total yağ asidi bileşiminde doymuş yağ asitlerinin toplamı %39.60 olarak bulunmuştur. Bu yüzdenin oluşmasında palmitik asidin yüksek yüzdesinin önemli bir rolü vardır. Doymamış yağ asitlerinin toplamı
oleik ve palmitoleik asitlerinin yüksek yüzdeleri sayesinde %37.37’dir. Aşırı doymamış yağ asitlerinin toplamının ise %23.03 olarak bulunmuştur. Görüldüğü gibi total yağ asidi bileşiminde yer alan yağ asitlerinin içerisinde en çok bulunan yağ asidi grubu, doymuş yağ asitleridir.
Sonbahar mevsiminde ω-6 yağ asitlerinin yüzdeleri toplamı %9.77 iken ω-3 yağ asitlerinin yüzdeleri toplamı %13.27 olarak tespit edilmiştir. Sonbahar mevsiminde ω-3/ω-6 (veya n-3/n-6) oranı 1.36 olarak bulunmuştur.
4.3. Kış Mevsiminde Sparus aurata’nın Total Yağ Asidi Bileşimi
S. aurata’nın kış mevsimi total yağ asidi bileşimi Tablo 2’de verilmiştir. Kış mevsiminde S. aurata’nın total yağ asidi bileşiminde en yüksek yüzdeye sonbahar mevsiminde olduğu gibi oleik asit sahiptir (%28.90). Oleik asidin yanı sıra palmitik asidin %24.33 ve dokosahekzaenoik asidin de %10.38 gibi büyük yüzdelere sahip olduğu tespit edilmiştir. Bu yağ asitlerine ilaveten stearik asit, palmitoleik asit, linoleik asit ve miristik asit de yüksek oranlarda bulunan yağ asitleridir. Yüksek oranlarda bulunan bu yağ asitlerinin toplam yüzdeleri yağ asidi bileşiminin %91.35’ ini oluşturmuşlardır.
Oleik asit, palmitik asit, dokosahekzaenoik asit, stearik asit, palmitoleik asit, linoleik asit, miristik asit dışında kalan eikosapentaenoik asit, γ-linolenik asit, pentadesilik asit, miristoleik asit ve linolenik asit toplam bileşimin %8.65’ini oluşturmuşlardır.
Sağlık açısından varlığı önemli olan yağ asitlerinden eikosapentaenoik asit (EPA) %3.95, dokosahekzaenoik asit (DHA) ise %10.38 olarak bulunmuştur.
Kış mevsiminde S. aurata’nın total yağ asidi bileşiminde doymuş yağ asitlerinin toplamının %39.21 olduğu görülmüştür. Doymamış yağ asitlerinin toplamı oleik ve palmitoleik asitlerinin yüksek yüzdeleri nedeniyle %38.29 olarak tespit edilmiştir. Aşırı doymamış yağ asitlerinin toplamı ise %22.50 olarak bulunmuştur. Aşırı doymamış yağ asitlerinin önemli bir bölümünü dokosahekzaenoik asit
oluşturmuştur. Total yağ asidi bileşiminde yer alan yağ asitlerinin içerisinde en çok bulunan yağ asidi grubu, doymuş yağ asitleridir.
Kış mevsiminde ω-6 yağ asitlerinin yüzdeleri toplamı %7.68 iken ω-3 yağ asitlerinin yüzdeleri toplamı %14.81 olarak tespit edilmiştir. Kış mevsiminde ω-3/ω-6 (veya n-3/n-ω-3/ω-6) oranı 1.93 olarak bulunmuştur.
Tablo 2. S. aurata’nın mevsimlere göre yağ asidi bileşimlerinin değişimi*
Yağ Asitleri Sonbahar Kış İlkbahar Yaz
C14:0 4.33±0.28a 4.48±0.27ab 6.90±0.12c 5.63±0.33b
C14:1 0.28±0.04a 0.53±0.03b 0.22±0.02a -
C15:0 0.62±0.06a 0.55±0.05a 0.55±0.02a 0.62±0.04a C16:0 24.61±0.31ab 24.33±0.52ab 23.32±0.39a 26.45±0.45b C16:1 8.27±0.37a 9.07±0.10a 12.41±0.07b 7.28±0.15c C18:0 10.03±0.32a 9.65±0.21a 5.39±0.20b 10.10±0.38a C18:1 28.82±0.71a 28.90±0.26a 25.37±0.23b 28.06±0.99a C18:2 5.37±0.19a 4.54±0.06b 4.02±0.04b 4.48±0.27b C18:3 (n-6) 4.40±0.31a 3.14±0.19b 10.21±0.16c 5.30±0.17a C18:3 (n-3) 1.07±0.07a 0.48±0.02b 1.27±0.11a 1.50±0.14a C20:5 3.47±0.15a 3.95±0.06a 2.11±0.20b 3.28±0.17a C22:6 8.73±0.19a 10.38±0.19b 8.23±0.12a 7.30±0.17c
ΣD.Y.A 39.60±0.76a 39.21±0.31ab 36.16±0.21b 42.80±1.10c ΣDym.Y.A 37.37±0.89a 38.29±0.34a 38.00±0.18a 35.33±0.93a ΣA.Dym.Y.A 23.03±0.67a 22.50±0.25a 25.84±0.08b 21.87±0.37a
Σ ω-6 9.77 7.68 14.23 9.78
Σ ω-3 13.27 14.81 11.61 12.08
Σ ω-3/ω-6 1.36 1.93 0.82 1.24
*Aynı satırda aynı harfle gösterilen değerler birbirinden farklı değildir (p> 0.05). Σ D.Y.A., Doymuş yağ asitlerinin yüzdeleri toplamı.
Σ Dym.Y.A, Doymamış yağ asitlerinin yüzdeleri toplamı. Σ A.Dym.Y.A, Aşırı doymamış yağ asitlerinin yüzdeleri toplamı.
4.4. İlkbahar Mevsiminde Sparus aurata’nın Total Yağ Asidi Bileşimi
S. aurata’nın ilkbahar mevsimi total yağ asidi bileşimi Tablo 2’de verilmiştir. İlkbahar mevsiminde de S. aurata’nın total yağ asidi bileşiminde en yüksek yüzdeye sahip olan yağ asidinin %25.37 gibi bir yüzdeyle oleik asit olarak tespit edilmiştir. Oleik asidin yanı sıra palmitik asit ile palmitoleik asit de büyük yüzdelerde tespit edilmiştir. Bu yağ asitlerine ilaveten γ-linolenik asit, dokosahekzaenoik asit, miristik asit, stearik asit ve linoleik asit de yüksek oranda bulunan yağ asitleridir. Yüksek oranlarda bulunan bu yağ asitlerinin toplam yüzdeleri yağ asidi bileşiminin %95.85’ini oluşturmuştur.
Yukardaki sekiz yağ asidi dışında kalan eikosapentaenoik asit, linolenik asit pentadesilik asit ve miristoleik asit yüzdeleri toplam bileşimin ancak %4.15’ ini oluşturmuşlardır.
Sağlık açısından varlığı en önemli yağ asitlerinden olan eikosapentaenoik asit (EPA) %2.11, dokosahekzaenoik asit (DHA) ise %8.23 olarak bulunmuştur.
İlkbahar mevsiminde S. aurata’nın total yağ asidi bileşiminde doymuş yağ asitlerinin toplamı %36.16 olarak bulunmuştur. Bu yüzdenin büyük bir kısmını palmitik asit oluşturmuştur. Doymamış yağ asitlerinin toplamı ise %38.00 olarak tespit edilmiştir. Bu yüzdenin oluşmasında oleik asit ve palmitoleik asidin yüksek oranda bulunmasının önemli bir rolü vardır. Aşırı doymamış yağ asitlerinin toplamının ise γ-linolenik asit ve dokosahekzaenoik asit sayesinde %25.84 olarak bulunmuştur. İlkbahar mevsiminde diğer üç mevsimden farklı olarak total yağ asidi bileşiminde yer alan yağ asitlerinin içerisinde en çok bulunan yağ asidi grubu, doymamış yağ asitleridir.
İlkbahar mevsiminde ω-6 yağ asitlerinin yüzdeleri toplamı %14.23 iken ω-3 yağ asitlerinin yüzdeleri toplamı %11.61 olarak tespit edilmiştir. İlkbahar mevsiminde ω-3/ω-6 (veya n-3/n-6) oranının 0.82 olduğu görülmüştür. Diğer mevsimlerde bu oran 1’den yüksek iken bu mevsimde 1’in altında bulunmuştur.
4.5. Yaz Mevsiminde Sparus aurata’nın Total Yağ Asidi Bileşimi
S. aurata’nın yaz mevsimi total yağ asidi bileşimi Tablo 2’de verilmiştir. Bu mevsimde 11 yağ asidi çeşidi tespit edilmiştir. Yaz mevsiminde S. aurata’nın total yağ asidi bileşiminde en yüksek orana %28.06’lık bir yüzdeyle oleik asit sahiptir. Oleik asidin yanı sıra palmitik asit ve stearik asit de büyük yüzdelerde tespit edilmiştir. Bu yağ asitlerine ilaveten dokosahekzaenoik asit, palmitoleik asit, miristik asit, γ-linolenik asit, linoleik asit de yüksek oranda bulunan yağ asitleridir. Yüksek oranlarda bulunan bu yağ asitlerinin toplam yüzdeleri yağ asidi bileşiminin %94.60’ını oluşturmaktadır.
Oleik asit, palmitik asit, stearik asit, dokosahekzaenoik asit, palmitoleik asit, miristik asit, γ-linolenik asit, linoleik asit dışında kalan eikosapentaenoik asit, linolenik asit ve pentadesilik asit yüzdeleri toplam bileşimin ancak %5.40’ını oluşturmuşlardır. Yaz mevsiminde diğer mevsimlerden farklı olarak miristoleik aside rastlanmamıştır.
Sağlık açısından varlığı önemli olan yağ asitlerinden eikosapentaenoik asit (EPA) %3.28, dokosahekzaenoik asit (DHA) ise %7.30 olarak bulunmuştur.
Yaz mevsiminde S. aurata’nın total yağ asidi bileşiminde doymuş yağ asitlerinin toplamının %42.80 olarak bulunmuştur. Bu yüzdenin oluşmasında palmitik asidin %26.45 gibi yüksek bir yüzdeye sahip olmasının önemli bir rolü vardır. Doymamış yağ asitlerinin toplamı oleik ve palmitoleik asitlerinin yüksek yüzdeleri sayesinde %35.33’dür. Aşırı doymamış yağ asitlerinin toplamının ise %21.87 olarak bulunmuştur. Bu mevsimde total yağ asidi bileşiminde yer alan yağ asitlerinin içerisinde en çok bulunan yağ asidi grubu, %42.80’lik bir yüzdeye sahip olan doymuş yağ asitleridir.
Yaz mevsiminde ω-6 yağ asitlerinin yüzdeleri toplamı %9.78 iken ω-3 yağ asitlerinin yüzdeleri toplamı %12.08 olarak tespit edilmiştir. Yaz mevsiminde ω-3/ ω-6 (veya n-3/n-6) oranı 1.24 olarak bulunmuştur.
5. TARTIŞMA
Balık yağlarının kompozisyonunu oluşturan üç temel yağ asidi tipi vardır.
Bunlar doymuş, doymamış ve aşırı doymamış yağ asitleridir (Halver 1989). S. aurata’nın doymuş yağ asitleri %36.16 oranı ile ilkbahar mevsiminde en düşük
seviyede, %42.80 oranı ile yaz mevsiminde en yüksek seviyededir. Aşırı doymamış yağ asitlerinde ise %21.87’lik bir yüzdeyle yaz mevsiminde en düşük, %25.84’lik yüzdeyle ilkbahar mevsiminde en yüksek oranda bulunmaktadır. Doymuş yağ asitleri yüzdeleri ise mevsimler arasında farklı bulunmamıştır (p>0.05). Doymamış ve aşırı doymamış yağ asitleri toplamı her mevsim için doymuş yağ asitlerinden yüksek olarak tespit edilmiştir.
Balıklardaki doymuş yağ asitleri yem kökenli olabilir, bağırsaktaki bakteriler tarafından oluşturulur veya mevcut bakterilerden absorbe edilir. Başlıca doymuş yağ asitleri C 14:0 miristik, C 16:0 palmitik ve C 18:0 stearik asitlerdir. Başlıca doymamış yağ asitleri ise C 14:1 miristoleik, C 16:1 palmitoleik ve C 18:1 oleik asitlerdir. Bu doymamış yağ asitlerini besinlerden biyosentez yolu ile sağlarlar. Aşırı doymamış yağ asitleri içerisinde ise yaygın olarak bilinen C 20:5 eikosapentaenoik asit (EPA) ve C 22:6 dokosahekzaenoik asit (DHA)’tir (Halver 1989). Yaptığımız çalışmada da doymuş yağ asitleri palmitik asit, stearik asit ve miristik asit olarak bulunmuştur Doymamış yağ asitlerinden oleik asit ve palmitoleik aside yüksek oranlarda rastlanırken, miristoleik aside oldukça düşük oranda rastlanmıştır. Aşırı doymamış yağ asitlerinden dokosahekzaenoik asit yüksek yüzdelerde iken, eikosapentaenoik asit düşük sayılabilecek yüzdelerde tespit edilmiştir.
Balıkların yağ asidi bileşiminde diğer hayvanlarınkine nazaran daha fazla sayıda yağ asidi çeşidi yer alır. Her ne kadar balıkların yağ asidi bileşimi çok sayıda yağ asidinden oluşuyorsa da bunlardan sadece birkaç tanesi total yağ asidi bileşiminin yüksek bir yüzdesini oluşturmaktadır (Gunstone 1986). S. aurata’nın yağ asidi bileşiminde karbon sayısı ve doymuşlukları farklı olan 12 yağ asidi gözlenmiştir. Bunlardan oleik asit, palmitik asit, palmitoleik asit, stearik asit ve dokosahekzaenoik asit yüzdeleri total yağ asidi yüzdesinin %74.72-82.33’lik kısmını
oluşturmaktadır. Yağ asidi bileşimlerindeki yağ asitlerinin yüzdeleri mevsimlere göre değişiklik göstermelerine rağmen bütün mevsimlerde en yüksek yüzdeye sahip yağ asidi çeşidi olarak oleik asit bulunmuştur (%25.37-28.90).
Farklı araştırıcılar balık dokusunda en çok bulunan yağ asidinin palmitik asit olduğunu tespit etmişlerdir. Yapmış olduğumuz çalışmada dört mevsimde de oleik asitten sonra %23.32-26.45 gibi yüksek bir yüzde ile palmitik asidin geldiği gözlenmiştir. Balık yağ asidi bileşimi içinde palmitik asidin yüksek olmasının nedeni; yağ asidi metabolizmasında rol oynadığından kaynaklanabileceği ileri sürülmüştür (Wang ve ark. 1990, Andrade 1995).
Kıraç (2004), Konya’da tüketilen bazı balıkların yağ asidi bileşimi üzerine yapmış olduğu çalışmasında, çipura (S. aurata) türünde en yüksek yağ asidi yüzdesinin %21.23 ile oleik aside ait olduğunu göstermiştir. Oleik asidi %15.76 ile palmitik asit, %13.20 ile linoleik asit ve %12.07 ile dokosahekzaenoik asit izlemiştir. Bu balığın yağ asidi bileşiminde en düşük yüzdeye %0.19 ile miristoleik asitte rastlanmıştır. Yapmış olduğumuz çalışmada da her mevsimde en yüksek yüzdeye sahip yağ asidi çeşidi oleik asit olarak tespit edilmiştir (%25.37-28.90). Her mevsimde oleik asitten sonra en yüksek yüzdeye palmitik asit sahiptir (%23.32- 26.45). Çalışmamızda üçüncü sıradaki yağ asidi, sonbahar ve yaz mevsiminde stearik asit, kış mevsiminde dokosahekzaenoik asit, ilkbahar mevsiminde palmitoleik asit olarak bulunmuştur. Kıraç’ın yapmış olduğu çalışmada linoleik asit %13.20 gibi yüksek bir yüzdeye sahipken; yaptığımız çalışmada ise her mevsimde düşük seviyelerde tespit edilmiştir (%4.02- 5.37). Yine Kıraç’ın çalışmasında %4.00 gibi önemli bir yüzde ile C 20:4 arakidonik asit mevcutken, balıklar için önemli yağ asitlerinden olan eikosapentaenoik aside rastlanmamıştır. Çalışmamızda hiçbir mevsimde arakidonik aside rastlanmazken, eikosapentaenoik asit ise %2.11-3.95 gibi bir yüzdeye sahiptir. Yapmış olduğumuz çalışmada da miristoleik asit genel olarak en düşük yüzdeye sahip yağ asididir. Bu farklılıklara, balığın yaşadığı ortama bağlı olarak oluşan diet farklılığı ve su sıcaklığı gibi durumlar, cinsiyet ve üreme durumları neden olabilir.
Öztürk (2003), Beyşehir gölünde yaşayan Tinca tinca’ yağ asidi bileşiminin mevsimsel değişimini incelediği çalışmasında en yüksek yüzdeye kış ve ilkbahar
mevsiminde oleik asidin, sonbahar ve yaz mevsiminde ise palmitik asidin sahip olduğunu tespit etmiştir. Sağlık açısından varlığı önemli olan yağ asitlerinden eikosapentaenoik asit %0.95-1.78, dokosahekzaenoik asit %3.78-7.62 arasında değişiklik göstermektedir. Yine aynı çalışmada ω-3/ω-6 oranı hiçbir mevsimde 1’i geçmemiştir. Yaptığımız çalışmada, her mevsim oleik asit en yüksek yüzdeye sahip yağ asidi olarak tespit edilmiştir. Eikosapentaenoik asit %2.11-3.95 arasında, dokosahekzaenoik asit %7.30-10.38 arasında değişiklik göstermektedir. ω-3/ω-6 oranı ise ilkbahar mevsimi haricinde hiçbir mevsimde 1’in altına inmemiştir (0,82-1,93). Bu değerlerdeki farklılıklara balıkların gölde veya denizde yaşaması ve beslenme biçimi neden olabilir. Öztürk’ün çalışmasında kullanılan T. tinca Beyşehir gölünde yaşayan omnivor bir türdür. Çalışmamızda kullanılan S. aurata ise Mersin yöresinden yakalanmış karnivor bir türdür.
Balıkların karakteristik yağ asitlerinden olan eikosapentaenoik asit ve dokosahekzaenoik asit gibi aşırı doymamış yağ asitlerinin yüzdeleri karnivor türler ile alg ve yüksek bitkilerle beslenen herbivor türler için daha yüksek iken, omnivor türler için daha düşük seviyelerdedir (Roy ve ark. 1999). Bir karnivor tür olan S.aurata’nın total yağ asidi bileşiminde dokosahekzaenoik asit yüksek sayılabilecek yüzdelere sahipken, eikosapentaenoik asit düşük yüzdelerdedir.
Yapılan çalışmalar balıkların eşeysel olgunluk dönemlerinde lipidleri daha fazla tercih ettikleri ve bu dönemlerde enerji ihtiyaçlarını kas dokusundan sağladıkları ifade edilmiştir (Nevsoma ve Leduc 1975, Medford ve Mackay 1978). Tatlı su balıklarından olan Aplodinotus grunniens, Coregonus artedii, Lota lota ve Alosa pseudoharengus türleri üzerinde yapılan çalışmada, üreme periyodu öncesinde lipid miktarının arttığı, üreme periyodu sonunda da azaldığı bildirilmiştir (Ackman 1967). Yaptığımız çalışma da bu sonuçlara uygunluk göstermektedir. S. aurata’nın üreme dönemi olan ekim ve aralık aylarına denk gelen sonbahar mevsiminden önceki dönem olan yaz mevsiminde total lipid oranı yüksek seviyede iken, üreme dönemi sonuna denk gelen kış mevsiminde total lipid oranı en düşük seviyededir.
Sıcak denizlerde yaşayan çinakop (Pomatomus saltatrux), barbun (Mullus barbatus) ve kefal (Mugil cephalus) balıklarında en yüksek yağ asidi yüzdesi oleik asit olarak tespit edilmiştir. Oleik asidi palmitik asit izlemiştir (Kıraç 2004). Mersin
yöresinden yakalanan çipurada da en yüksek yağ asidi oranına oleik asit sahiptir. Bu yağ asidini palmitik asit izlemiştir. Bu sonuçlara göre yaptığımız çalışma Kıraç’ın çalışmasını desteklemektedir.
Balık yağlarının bileşiminde yer alan fakat sentezleyemedikleri linoleik asit, linolenik asit ve arakidonik asit aynı zamanda insanlar için de esansiyel yağ asitleridir. Yani bu yağ asitleri vücutta sentezlenemediği için besinle dışarıdan hazır olarak alınma zorunluluğu vardır (Gunstone 1986). Çalışmamızda arakidonik aside rastlanmazken, linoleik asit ve linolenik asit önemli miktarlarda bulunmuştur.
Balığın yağ asidi bileşimini etkileyen en önemli faktörlerden biri de besininin ihtiva ettiği yağ asidi bileşimidir. Genellikle linoleik asit, linolenik asit ve eikosapentaenoik asit gibi yağ asitleri bakımından zengin olan tatlı su algleri, sucul böcek larvaları balıklar için önemli besin kaynaklarıdır (Farkas 1970, Wood 1974, Takahashi ve Yamada 1976, Hanson ve ark. 1985). Eikosapentaenoik asit ve dokosahekzaenoik asit bakımından zengin fakat ω-6 aşırı doymamış yağ asitleri bakımından fakir olan planktonlar (Sargent ve ark. 1989) deniz balıklarının temel yiyecekleridir (Kelly ve ark. 1959, Kayama ve ark. 1963). Planktonlarla beslenen deniz balıklarında ω-3 seviyesi daha yüksektir. Öztürk (2003)’ün tatlı su balığı olan T. tinca ile yapmış olduğu çalışmada ω-3/ω-6 oranı hiçbir mevsimde 1’i geçmemiştir. Çalışmamızda deniz balığı olan S. aurata’nın ω-3/ω-6 oranı genel olarak 1’in üzerinde bulunmuştur.
Balık yağı insan sağlığı açısında son derece önemlidir (Connor ve Connor 1986). Balık yağları başta kalp-damar hastalıkları olmak üzere ölüme sebebiyet verebilen birçok hastalığın iyileştirilmesinde rol oynamaktadır. Balık yağını sağlık açısından önemli kılan özellik, balıkların yağ asidi bileşiminde karbon sayısı ve doymamışlığı yüksek yağ asitlerinin bulunmasıdır. Bu yağ asitlerinin başında eikosapentaenoik asit ve dokosahekzaenoik asit gelmektedir (Steffens 1997). Yapmış olduğumuz bu çalışmada, sağlık açısından önemli olan bu iki yağ asidinden dokosahekzaenoik asit %7.30-10.38 gibi yüksek sayılabilecek yüzdelerde iken, eikosapentaenoik asit ise daha düşük yüzdelere sahiptir (%2.11-3.95).
Sağlığın korunması için özellikle soğuk sularda yaşayan balıkların tüketiminin yüksek olduğu Grönland tipi beslenme şekli önerilmekte ve kardiovasküler sistem bozukluklarından ölümlerin engellenmesi için ister deniz balığı, ister tatlı su balığı olsun balık tüketiminin arttırılması teşvik edilmektedir (Bang ve Dyerberg 1972, Dyerberg ve ark. 1975, 1978, Puwastien ve ark. 1999, Jonkers ve ark. 2000).
Bu çalışmada, S. aurata’nın total lipid ve total yağ asidi bileşimlerinin mevsimsel değişimi incelenmiştir. Her mevsimde doymamış ve aşırı doymamış yağ asitleri toplam yüzdeleri doymuş yağ asitleri yüzdelerinden yüksek olarak bulunmuştur. Total lipid miktarı sonbahar mevsiminde en yüksek değerde, ω-3 miktarı ise kış mevsiminde en yüksek değerde bulunmuştur. Bu değişimlere, suyun sıcaklık derecesi, besin çeşidi ve üreme durumları gibi etkenler neden olabilir.
Bu tez, S. aurata türünün total lipid ve total yağ asidi bileşimine mevsimsel değişimin etkilerinin araştırıldığı ilk çalışma olması nedeni ile önemlidir.
6. KAYNAKLAR
Ackman, R.G., 1967. Characteristics of the Fatty Acid Composition and Biochemistry of Some Freshwater Fish Oils And Lipids in Comparison with Marine Oils and Lipids. Comp. Biochem. Physiol., 22, 907-922.
Ackman, R.G., 1988. Concerns for utilization of marine lipids and oils, Food Technology; 151-160.
Agren, J., Mute, P., Hanninen, O., Herranen, J., Pentila, I., 1987. Seasonal Variations of Lipid Fatty Acids of Boreal Freshwater Fish Species. Comp. Biochem. Physiol., 88B: 905-909.
Ahlgren, G., Blomquist, P., Bober, M., Gustafsson, I.B.,1994. Fatty acid content of the dorsal muscle an indikator of fat quality in freshwater fish. Journal of Fish Biology, 45, 131-157.
Ahlgren, G., Carlstein, M., Gustafsson, I.B., 1999. Effects of natural and commercial diets on the fatty acid content of European grayling. Journal of Fish Biology, 55, 1142-1155.
Akpınar, M.A., 1987. Cyprinus carpio L. (Osteichthyes, Cyprinidae)’nın kas dokusu yağ asitlerinin mevsimsel değişimi. Doğa Türk Biyoloji. 11(1).
Akşiray, F., 1954. Türkiye Deniz Balıkları Tayin Anahtarı. Pulhan Matbaası, İstanbul, 257.
Andrade, A.D., 1995. Omega-3 fatty acids in freshwater fish from South Brazil. Jour. Of The American Oil Chemists, 72 (10), 1207-1210.
Bang, H.O., Dyerberg, J., 1972. Plasma lipids and lipoproteins in Greenlandic West Coast Eskimos. Acta Med. Scand., 192, 85-94.
Bell, J.G., Mcevoy, J., Tocher, D.R., McGhee, F., Campbell, P.J., Sargent, J.R., 2001. Journal of nutrition, 131, 5 , 1535-1543.
Boggio, S.M., Hardy, R.W., Rabbitt, J.K., Brannon E.L., 1985. Aquaculture, 51, 13-24.
Brenner, R.R., 1989. Factors Influencing Fatty Acid Chain Elongation and Desaturation. In The Role of Fats in Human Nutrition (Vergroesen, A.J., Crawford, M., eds), p. 45-79. London: Academic Pres.
Burr, M.L., Fehily, A.M., 1990. Fatty Fish and Heart Disease. World Review of Nutrition and Dietetics., 256-257.
Cai, Z., Curtis, L.R., 1989. Effects of Diet on Consumption, Growth and Fatty Acid Composition in Young Grass Carp. Aquaculture, 81, 47-60.
Charalambakis, G., Divanach, P., 1993. International Journal of Food Sciences and Nutrition, 45, 135-139.
Cho, C.Y., Cowey, C.B., Watanabe, T., 1985. Finfish nutrition in Asia. Methodological aproaches to reseach and devolopment Int. Devolop. Res. Cent. 1:26-33.
Connor, W. E., Connor, S.L., 1986. Dietary cholestorol and fat the prevention of coronary heart disease: Risk and benefits of nutritional change. In diet and prevention of coronary heart disease and cancer (Hallgren, B., Leven, O., Rossner, S., Vessby, B. Eds.), pp. 113-147.
Crawford, M., Marsh, D., 1989. The driving force. London: Heinemann.
Csengeri, I., Albrecht, M.L., Steffens, W., Olah, J., 1986. Archiv Fur Tierernahrung, 36, 7, 653-663.
Deng, J.C., Orthefer, F.T., Dennison, R.L., Watson, M., 1976. Lipids and Fatty Acids in Mullet (Mugil cephalus): Seasonal and Locational Variations. Jour of Food Science, 4, 1479-1483.
Domaizon, I., Desvilettes, C., Debroas, D., Bourdier, G., 2000. Journal of Fish Biology, 57, 2, 417-432.
Dönmez, M., Tatar, O., 2001. Fleto ve Bütün Olarak Dondurulmuş Gökkuşağı Alabalığının (Oncorhynchus mykiss W.)Muhafazası Süresince Yağ Asitleri Bileşimlerindeki Değişimlerin Araştırılması. Ege Üniversitesi, Su Ürünleri Dergisi, Cilt 18, Sayı (1-2): 125-134.
Dyerberg, J., Bang, H.O., Hjqrne, N., 1975. Fatty acid composition of the plasma lipids in Greenland Eskimos. Am. J. Clin. Nutr. 28, 958-966.
Dyerberg, J., Bang, H.O., Stoffersen, E., Moncada, S., Vane, J.R., 1978. Eikosapentaenoic acid prevention and thrombosis and atheroselerosis. Lancet, 2, 117-119.
Farkas, T., 1970. Fats in freshwater crustaceans, 1, Fatty acid composition of lipids obtained from Eudiaptomus gracilis G.O. Sars (Copepoda) and Daphnia cucullata G.O. Sars (Cladocera), Acta Biol. Acad. Sci. Hung., 21, 225-233. Forss, D.A., 1967. Role of Lipids of Flavors. J. Agr. Food Chem., 17:681.
Fremont, L., Marion, D., 1982. A comparison of hhe lipoprotein profiles in male trout (Salmo gairdneri) before maturity and during spermiation. Comp. Biochem. Physiol. B., 73, 849-855.
Galli, C., Butrum, R., 1990. Dietary omega-3 fatty acids and cancer. World Review of Nutrition and Dietetics., 66, 446-461.
Gallagher. M.L., Harrell. M.L., Rulfson R.A., 1991. Variation in lipid and fatty acid contents of Atlantic croakers striped mullet and summer flounder. Trans Am. Fish. Soc. 120:614-619.
Gibson, R.A., 1983. Ausralian Fish-an Excellent Source of Both Arachidonic Acid and Omega-3 Polyunsaturated Fatty Acids. Lipids, 18 (11), 743-750.
Gibson, R.A., Kneebone, R., Kneebone, G.M., 1984. Comparative levels of arachidonic acide in Malasian fish. Comp. Biochem. Physiol. 78 C: 325-328. Gögüş, A.K., 1988. Su Ürünleri İşleme Teknolojisi, K.T.U. Sürmene Deniz
Bilimleri ve Teknolojisi Yüksekokulu, Ders Kitabı No:19, Trabzon.
Gunasekara, R.M., Silva, S.S.D., Ingram, B.A., 1999. Early ontogeny-related Changes of the fatty acid composition in the Percichthyid fishes trout cod, Maccullochella macquariensis and Murray cod, M. peelii peeli. Aqua Living Resour., 12(3), 219-227.
Gunstone, F., 1986, The Lipid Handbook, Mc Graw and Hill.
Hagen, W., Kattner, G., Graeve, M., 1995. On The Lipid Biochemistry of Polar Copepodes: Compositional Differences in The Antartict, Marine Biology, 123, 451-457.
Halver, J.E., 1972. Fish Nutrition, Academic Pres Inc., Second Ed., New York, 723p. Halver, J.E., 1989. Fish Nutrition, Academic Pres Inc., Second Ed., New York, 798p. Hanson, B.J., Cummins, K.W., Cargill, A.S., Lowry, R.R., 1985. Lipid content, fatty acid composition and the effect of diet on fats of aquatic insects, Comp. Biochem. Physiol. B, 80, 257-276.
Hawkins, P., 1997. Essential Fatty Acids in Fishes and Time Course of Changes in Fatty Acids Composition of Liver, Blood and Carcass Induced by a Diet Deficient in n-3 and n-6 fatty Acids, 75.
Hayashi, K., Takagi, T., 1977. The Seasonal variation in lipids and fatty acids of sardine (Sardinaps melanosticta). Bull. Fac. Fish. 28(2):83-94.
Henderson, R.J., Tocher, D.R., 1987. The lipid composition and biochemistry of freshwater fish. Prog. Lipid Research 26, 281-347.
Henderson, R.J., Burkow, I.C., Buzzi, M., Bayer, A., 1998. Biochimica et Biophsica Acta, 1392, 309-319.
Huss, H., 1988. Fresh Fish Quality and Quality Changes. Ministery of Fisheries Technical Universty Press , Copanhagen, Denmark.
Jangaard, P.M., Brockerhoff, H., Burger, R.D., Hoyle, R.J., 1967. Seasonal Changes in General Condition and Lipid Content of Cod From Inshare Waters. Jour Fisheries Research Board of Canada, 24, 607-612.
Johnson, L. L., Casillas, E., 1991. The use of Plasma parameters to predict ovarian maturation stage in English sole Parophrys vetulus Girard., j. Exp. Mar. Biol. Ecol., 151, 257-270.
Johansson, L., Kiessling, A., Kiessling, K.H., Berglund, L., 2000. Food Quality and Preference, 11, 247-254.
Jonkers, I.J.A.M., Ledeboer, M.D.M., Steens, M.D.J., Smelt, A.H.M., Masclee, A.A.M., 2000. Effects of very long chain versus long chain trigliserides on gastrointestinal motility and hormone release in humans. Digestive Diseases and Sciences, 45, 1719-1726.
Kandemir, Ş., 1999. Derbent Baraj Gölünde Kültürü Yapılan Gökkuşağı Alabalığında (Oncorhynchus mykiss W., 1792) Total Yağ ve Yağ Asidi Miktarı ile Yağ Asidi Cinslerinin Aylara ve Mevsimlere Göre Değişimi. Ondokuz Mayıs Üniversitesi, Fen Bilimleri Enstitüsü. Biyoloji A.B.D., Doktora Tezi, 150s, Samsun.
Kanders, B., Kowalchuk, M., 1990. Omega-3 Fatty Acids and Cancer Metostasis in Humans. World Review of Nutrition and Dietetics, 66, 477-487.
Kayama, M., Tsuchiya Y., Mead J.F., 1963. A model experiment of aquatic food chain whit special significance in fatty acid conversion. Bulletin of The Japanese Society of Scientific Fisheries, 29, 452-458.
Kelly, P.B., Riser, R., Hood, D.W., 1959. The origin of the marine polyunsatureted fatty acids. Compositions of marine plankton, J. Am. Oil Chem. Soc. 36, 104-106.
Kıraç, E., 2004. Konya’da Satılan Bazı Balık Türlerinin Yağ Asidi bileşimi. Selçuk Üniversitesi, Fen Bilimleri Enstitüsü. Biyoloji A.B.D., Yüksek Lisans Tezi, 47s, Konya.
Kiessling, A., Pickova, J., Johansson, L., Asgard, T., Storebakken, T., Kiessling, K.H., 2001. Food Chemistry, 73, 271-284.
Leaf, A., Weber, P.C., 1988. Cardivasküler Effects of n-3 Fatty Acids. New England Journal of Medicine, 318.
Li, X., Steiner, M., 1990. Fish Oil, a Potent Inhibitor of Platelet Adhesiveness. World Review of Nutrition and Dietetics, 66, 562-563.
Love, R.M., 1970. The Chemical Biology of Fishes. Vol.2. Academic Press. London and New York.
MacFarlane, R..B., Norton, F.C., Bowers, M.J., 1993. Lipid Dynamics in Relation to The Annual Reprotuctive Cycle in Yellowtail Rockfish (Sebastes flavitus) Can. J. Fish. Aquat. Sci., 50, 391-400.
Medford, B.A., Mackay, W.C., 1978. Protein and lipid content of gonads, liver and muscle of Notherm pike (Esox lucuis) in relation to gonad growth. J. Fish Research, 35: 213, 219.
Neuhaus, O.W., Halver, J.C., 1969. Fish in Research, s.135, Academic Press. New York.
Nevsome, E.G., Leduc, G., 1975. Seasonal changes of fat content in the yellow perch (Perca flavens) of two laurentin lakes. J. Fish Red Bd. Can. 32 (11): 2214-2221.
Öztürk, A.K., 2003. Beyşehir Gölü’ndeki Kadife Balığı, Tinca tinca L.(Osteichthyes: Cyprinidae)’nın Total Yağ Asidi Bileşiminin Mevsimsel Değişimi. Selçuk Üniversitesi, Fen Bilimleri Enstitüsü, Biyoloji A.B.D., Yüksek Lisans Tezi, 43s, Konya.
Puwastien, P., Judprasong, K., Ketwann, E., Vasanachitt, K., Nakngamanong, Y., Bhattaccharjee, L., 1999. Proximate Composition of raw and Cooked Thai Freshwater an Marine Fish. Journal of Food Composition and Analysis, 12, 9-16.
Roy, R., Fodor, E., Kitajka, K., Farkas, T., 1999. Fatty acid composition of the ingested food only slightly affects physicochemical properties of liver total phospholipids and plasma membranes in cold-adapted freshwater fish. Fish Physiology and Biochemistry, 20, 1-110.
Sağlık, S., 1994. Bazı Balık, Midye ve Karides Türlerinin Yağ Asidi Kompozisyonları ve Kolesterol İçeriklerinin Gaz Kromatografik İncelenmesi. İstanbul Üniversitesi Sağlık Bilimleri Enstitüsü, Analitik Kimya A.B.D., Doktora Tezi, 60s, İstanbul.
Sargent, J.R., Henderson, R.J., 1980. Lipid Metabolism In Marine Animals. Biochem. Soc. Trans., 3, 296-297.
Sargent, J.R., Henderson, R.J., Tocher, D.R., 1989. The Lipids in Fish Nutrition. Pp 1554-218. edited by J.E. Halver. Academic Pres, New York.
Sargent, J.R., Bell, J.G., Bell, M.V., Henderson, R.J., Tocher, D.R., 1995. Requirement Criteria for Essential Fatty Acids. Journal of Applied Ichthyology, 11, 183-198.
Sinclair, A.J., 1982. Elevated Levels of Arachidonic Acid in Fish From Northern Australian Coastal Waters. Lipids, 18 (12), 877-881.
Skorski, Z., 1990. Sea Food, Resources, Nutritional Composition and Preservation. Crc. Press. Inc. Boca Rota, Florida , 41-44p.
SPSS, 1999. SPSS 10.0 Statistics. SPSS, Chicago, IL.
Steffens, W., 1997. Effects of variation in essential fatty acids in fish feeds on nutritive value of freshwater fish for humans. Aquaculture, 151: 97-119.
Takahashi, H., Yamada, M., 1976. Lipid composition of seven Species of crustacean plankton. Bull. Jpn. Soc. Sci. Fish., 42, 769-776.
Tornaritis, M., 1993. Departmant of Social Medicine, Preventative Medicine and Nutrition Clinic, University of Crete, Heraklio, Crete, Greece, Academic Pres, 43-45.
Uysal, K., 2000. Eğirdir Gölü Sudak (Stizostedion luciperca L. 1758) Balıklarının Total Lipid, Total Yağ Asidi ve Yağ Asidi Bileşiminin Mevsimsel İncelenmesi. Süleyman Demirel Üniversitesi, Su Ürünleri Temel Bilimleri Anabilim Dalı, Doktora Tezi, 66s, Isparta.
Ünlüsayın, M., 1999. Yılan Balığı (Anguilla anguilla L. 1766), Gökkuşağı Alabalığı (Oncorhynchus mykiss W. 1792) ve Sudak Balığı (Stizostedion luciperca L. 1758)’nın Sıcak Dumanlama Sonrası Lipid ve Protein Bileşimleri. Süleyman Demirel Üniversitesi, Su Ürünleri Temel Bilimleri Anabilim Dalı, Doktora Tezi, 57s, Isparta.
Wang, Y.J., Miller, L.A., Perren, M., Addis, P.B., 1990. Omega -3 Fatty Acids in Lake Superior Fish. Jour of Food Science, 55, 72-73.
Weatherley, A.H., Gill, H.S., 1989. The Biology of Fish Growth, Academic Press, London, 442p.
Weber, P.C., Leaf, A., 1990. Cardiovaskuler Effects of n-3 Fatty Acids. World Review of Nutrition and Dietetics, 66, 218-232.
Williams, E.E., Hazel, J.R., 1992. The role of docosahexaenoic acid-containing molecular species of phospholipids in the thermal adaptation of biological membranes. In essential fatty acids and eicosanoids, pp. 128-133. Edited by A. Sinclair and R. Gibson. Am. Oil Chemists’ Society, Champaign, Illinois. Wood, B.J.B., 1974. Fatty acid and saponifiable lipids. In: W.D.P. Stewart (Editor).
Algae Physiology and Biochemistry. Blackwell, Edinburgh, pp. 29, 236-265. Yılmaz, Ö., Konar, V., Çelik, S., 1996. Elazığ Hazar Gölünde Yaşayan Capoeta
capoeta umbla (Heckel, 1843)’nın (Siraz) Total Lipid ve Yağ Asidi Miktarlarının Aylara ve Mevsimlere Göre Değişimi. Tr. J. Of Biology, 20:245-257.
Ziboh, V.A., 1990. Omega3 Polyunsaturated Fatty Acid Constituends of Fish Oil and the Management of Skin Inflammatory and Scaly Disorders. World Review of Nutrition and Dietetics, 66, 425-435.