Tüberküloza karşı bilgisayar destekli ilaç tasarımı; fosfodiesteraz için inhibitör geliştirilmesi
Tam metin
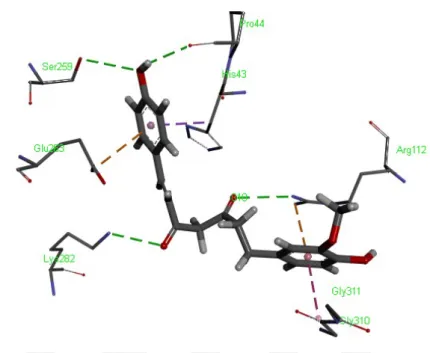
Şekil
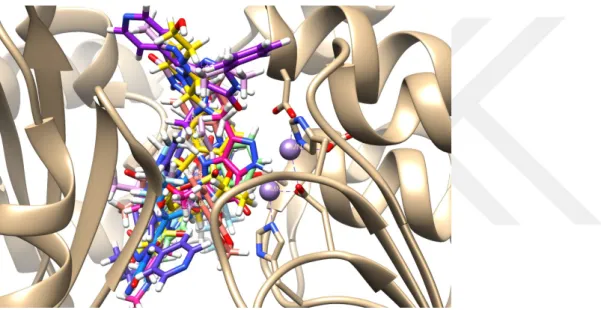
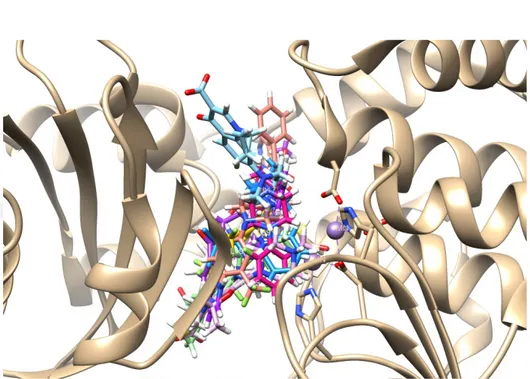
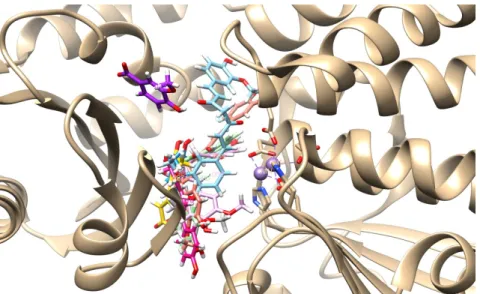
![Tablo 1. Literatürlerden Seçilen ve Tasarlanan Ligantlar O S NHN N NOOO NN (L-01)- Sildenafil (L-02)- Vardenafil (L-03)- (R)-Rolipram (4R)-4-[3-(cyclopentyloxy)-4-methoxyphenyl]pyrrolidin-2-one (L-04)- (L-05)- 3-[6-bromo-3-(cyclopentylamino)imidaz](https://thumb-eu.123doks.com/thumbv2/9libnet/3372464.12297/27.892.107.766.152.1061/literatürlerden-tasarlanan-ligantlar-sildenafil-vardenafil-cyclopentyloxy-methoxyphenyl-cyclopentylamino.webp)

Benzer Belgeler
Mekanik hareketi dairesel bir hareket olmayıp da yatay eksen ( x veya y eksenleri ) üzerinde hareket eden motorlara lineer motor denir.. Bu tür motorların tasarımı yapılırsa
Hastalıkla ilgili etmenin ve/veya hastalık-hedef-efektör üçgenindeki ilişkilerle ilgili bilgilerin araştırılıp, tanımlanarak açıklığa kavuşturulması ve elde edilen
Kimyasal bileşiklerin nicel olarak saptanan moleküler nitelikleri ile biyolojik etkileri arasındaki ilişkilerden yararlanılarak;.. «İdeal ilaç etken maddesi olabilecek yeni
• Indinavir: HIV-1 Protease Inhibitor developed via X- ray crystallography,. molecular mechanics
As a result of the paired t-test, the p-value was 0.002526, which is less than 0.05, so the null hypothesis was rejected at the significance level of 0.05, and it can be said there
Analiz sonuçlarını detaylı olarak incelediğimizde; çalışanların örgütsel destek algılarının işe angaje olmaları üzerinde anlamlı ve olumlu bir etkisi
Cases with a decrease in BCVA (change ≥5 letters) and a ≥10% increase in CFT received a repeat injection. Focal laser therapy was applied to patients who were detected to
Babası yarın yine aynı yere gidip balık tutmaya çalışacaktı.. Sakin kafayla
