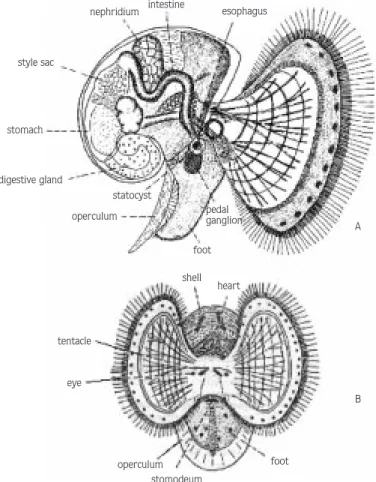
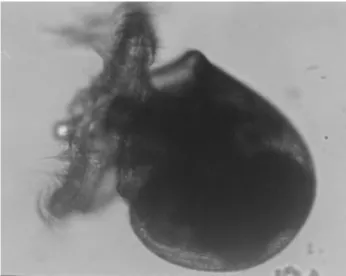
The larval development stages of the Japanese Snail, Rapana thomasiana, Gross 1861, in the egg capsule
Tam metin
Şekil

Benzer Belgeler
İşyer- lerinde işin yürütümü sırasında özellikle elle taşıma sırasında doğan olumsuz koşullardan işgöreni korumak, üretimin devamını, işletmenin güvenliğini
bu katkl maddelerinin bUyumeyi te~vik etmeleri ya- nlnda insan ve hayvan sagllglnl ciddi olarak tehdit eden bazl Van etkileri de ortaya ~lkml~llr (Teller ve
İstanbul örnekleminde 6 lisede toplam 927 lise bir, iki ve üçüncü sınıf öğrencisiyle yapılan başka bir çalışmada yaşam boyu bir kez kullanım % 5.2, son bir yıl
Bir aralık me’zunen Trabzonda bulunduğu sırada Türkçe ve Usuli Muhabere ve Neşriyat Muallimliği ilâvesile Trabzon Yüksek Ticaret Mektebi Müdirliğine nasb
— Şark ile Garbi barıştırmak için üzerimize düşen vazifeyi
Devrinin üstad hocaları olan Adil Bey ve Alman Hem VVemiya'dan ve yine meşhur ressam Valery’den ders almış olan Hayri Çizel, Akademiyi birincilikle bitirmiş
An cak birinci derecede tarihi eser sayılan otelde böyle bir değişiklik için İstanbul 1 No’lu Tabiat ve Kültür Varlıklarını Koruma Ku- rulu'nun
Gazete ve dergi yazılarını düzenli olarak takip etme oranı değişkeninin; öğrencilerin evrensel değerlere ilişkin tutumları üzerinde öntest sonuçlarına göre manidar
