Retinotopic sensitisation to spatial scale: evidence for flexible spatial frequency processing in scene perception
Tam metin
Şekil

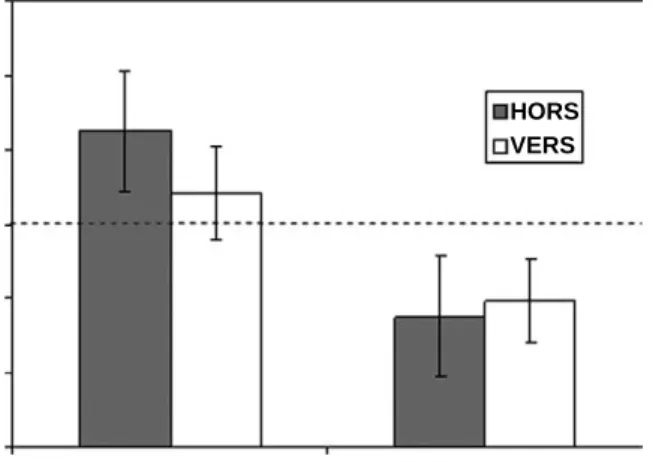
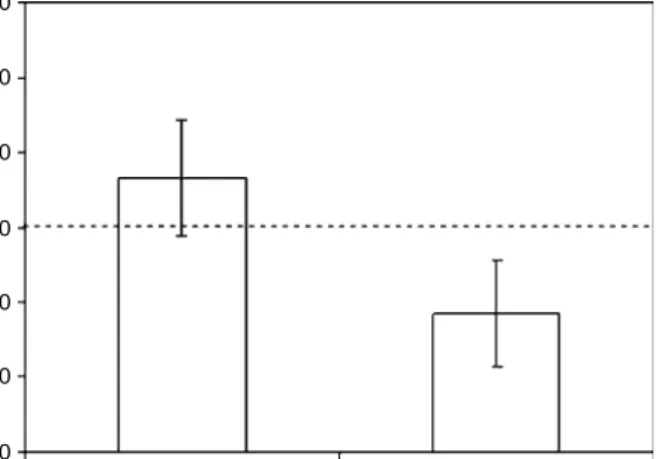
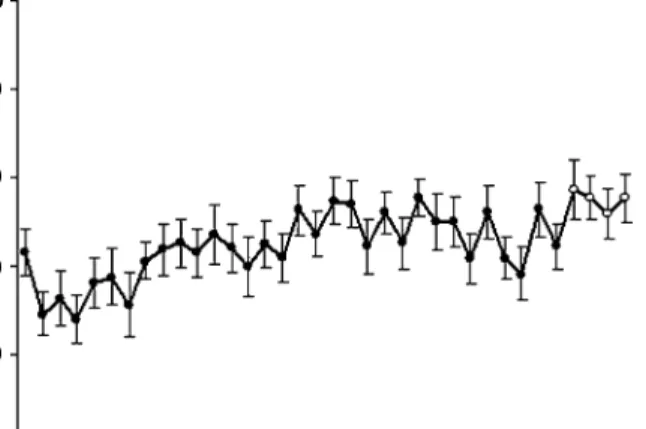
Benzer Belgeler
According to the findings obtained in regard of educational level variable in the related literature, it was seen that teachers’ pupil control ideology differed
In this paper, I have explored a few major examples of the use of music by composers and viewers to associate and assist in recalling the identity of major characters in the
Türk mitolojisinde Şamanist inanç ile pekiştirilmiş anahanlı örgütlenmenin, varoluşun fenomenal bilinç düzeyinde kavranmasına engel olması ve buna dayalı olarak
Chest CT images (Figs 1 and 2) showed peripheral, multilobar areas of ground-glass opacity sign suggesting diagnosis of novel coronavirus (COVID-19) pneumonia (Fig 2)..
34 The figures of merit we consider here for quantifying interfacial friction are the magnitude of the corrugation in the PES of interaction between Au and h-BN and that of the
We present a new dominance rule for the single machine total weighted tardiness problem with job dependent penalties. The proposed dominance rule provides a
4.17 Memory scale plots of three different sphere meshes for array struc- tures, which are related with far-field computations (load balancing algorithms are applied).. 54 4.18
Ahmet Cebeci, ideolojik bir bakış açısı ve hesaplamayla Türk nüfusunun Bulgaristan’da kalmaya devam etmesinin Türklerin lehine olacağını fakat yapılan hatanın
