T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
Aralık-2019 KONYA Her Hakkı Saklıdır FARKLI B VE NaCl DOZU
UYGULAMALARINDA Puccinellia distans (Jacq.) Parl. BĠTKĠSĠNĠN B VE Na TRANSPORTER
GENLERĠNĠN ĠFADE DÜZEYLERĠNĠN GERÇEK ZAMANLI ANALĠZĠ
Merve ĠNANÇ YÜKSEK LĠSANS
iv ÖZET YÜKSEK LĠSANS
FARKLI B VE NaCl DOZU UYGULAMALARINDA Puccinellia distans (Jacq.) Parl. BĠTKĠSĠNĠN B VE Na TRANSPORTER GENLERĠNĠN ĠFADE
DÜZEYLERĠNĠN GERÇEK ZAMANLI ANALĠZĠ Merve ĠNANÇ
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Toprak Bilimi ve Bitki Besleme Anabilim Dalı
DanıĢman: Prof. Dr. Erdoğan EĢref HAKKI 2019, 60 Sayfa
Jüri
Prof. Dr. Erdoğan EĢref HAKKI Prof. Dr. Sait GEZGĠN
Doç. Dr. Ceyda ÖZFĠDAN KONAKÇI
Dünyada ve ülkemizde, abiyotik ve biyotik stres faktörleri tarımsal verimi önemli ölçüde olumsuz yönde etkilemektedir. Tarım alanlarındaki bu abiyotik stres faktörlerinden toprak tuzluluğu ve bor toksisitesi sorunu kurak ve yarı kurak iklim bölgeleri başta olmak üzere, bataklıklar ve deniz kıyıları gibi aşırı nemli bölgelerde de görülmektedir. Tarım topraklarında tuzluluk ve bor toksisitesi sorununun giderilmesi toprak ıslahı yoluyla mümkün olmakla birlikte yetersiz kaliteli yıkama suyu ya da arazinin topoğrafik konumu gibi nedenlerle gerçekleştirilmesi mümkün olmamaktadır. Bu nedenle tuzluluk ve B toksisitesi sorununun giderilebilmesi ve verimli bitkisel üretimin gerçekleştirilebilmesi için bu alanlarda B ve tuza toleranslı kültür bitkilerinin tespit edilerek geliştirilmesi veya bu geliştirme aşamalarında akraba türlerden yararlanılması gerekmektedir.
Graminae (buğdaygil) familyasının Poaideae çim alt familyasından olan Puccinellia distans ‗çorak çim bitkisi‘ tuz ve B toksisitesine tolerans gösteren ve Türkiye‘de bor madenleri civarında da yaşayabilen hiperakümülatör ve çok yıllık bir bitki olarak karşımıza çıkmaktadır. Buğdaygiller familyasında olmasından dolayı da önemli tahıl grupları için model bir organizma olarak değerlendirilmektedir. Ancak, söz konusu bitkinin B ve NaCl stresi toleransı hakkında yeterince moleküler düzeyde çalışma bulunmamaktadır.
Araştırma materyali olan P. distans (Jacq.) Parl. tohumları Eskişehir Kırka‘da bulunan bor madenlerinde doğal olarak yetişen bir popülasyondan toplanmış ve iklim odasında kontrollü koşullarda su kültürü ortamında yetiştirilmiştir. Bitkiler bu çalışmada üç farklı dozda bor (0 mM, 0.003 mM ve 4 mM B) ve üç farklı dozda tuz (0 mM, 100 mM ve 250 mM NaCl) uygulamalarına maruz bırakılmıştır. Bu uygulama sonucunda elde edilmiş olan RNA örneklerinden kontrol ve stres şartlarında tespit edilmiş olan 4 adet B ve 1 adet Na ile ilişkili gen kullanılarak bu genlerin ifade düzeylerinin qRT-PCR ile belirlenmesi hedeflenmiştir. Referans gen olarak ise hedef genlerin normalizasyonunda optimize edilmiş olan Actin geni kullanılmıştır. Tez çalışması kapsamında P.distans‘ın yaprak örneklerinden bor ile ilişkili olabilecek Gene-21 ve tuz stresi ile ilişkili olabilecek NHA1 genlerinin ifade düzeyleri ölçülmüştür. P. distans‘ın göstermiş olduğu tolerans mekanizmasının moleküler düzeyde anlaşılması neticesinde ilgili bilgilerin buğday gibi ekonomik değeri yüksek bitkilere veya bu kültür bitkilerine daha yakın akrabalık ilişkisinde bulunan farklı bitkilerde anlaşılması ile ilgili hedef bitkilerde moleküler ıslahta kullanılabilecek karakterlerin belirlenmesine katkı sağlayabilecektir.
v ABSTRACT
MS THESIS
REAL-TIME EXPRESSION OF Puccinellia distans (Jacq.) Parl. B AND Na TRANSPOTER GENES UNDER DIFFERENT B AND NaCl TREATMENTS
Merve ĠNANÇ
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY
THE DEGREE OF MASTER OF SCIENCE IN SOIL SCIENCE AND PLANT NUTRITION
Advisor: Prof. Dr. Erdoğan EĢref HAKKI 2019, 60 Pages
Jury
Prof. Dr. Erdoğan EĢref HAKKI Prof. Dr. Sait GEZGĠN
Assoc. Dr. Ceyda ÖZFĠDAN KONAKÇI
In the world and in our country, abiotic and biotic stress factors significantly adversely affect agricultural productivity. The problem of soil salinity and boron toxicity among these abiotic stress factors in agricultural areas is seen in arid and semi-arid climates, as well as in extremely humid areas such as marshes and seashores. Although salinity and boron toxicity in agricultural soils can be solved through soil improvement, it cannot be realized due to reasons such as insufficient quality washing water or topographic location of the land. Therefore, in order to eliminate the salinity and B toxicity problem and to realize productive plant production, it is necessary to identify and develop B and salt tolerant culture plants in these areas or to use related species in these development stages.
Puccinellia distans ―arid grass plant‖ from the Poaideae grass subfamily of the Graminae (Poaceae) family is a perennial plant and hyperaccumulator that shows tolerance to salt and B toxicity and is also able to live around boron mines in Turkey. It is considered as a model organism for important cereal groups since it is in the grasses family. However, there is not enough molecular level studies on B and NaCl stress tolerance of the plant in question.
The research material P. distans (Jacq.) Parl. seeds were collected from a population that grows naturally in boron mines in Eskişehir Kırka and grown under controlled conditions in a climate chamber in a water cultivated environment. In this study, the plants were exposed to three different doses of boron (0 mM, 0.003 mM and 4 mM B) and three different doses of salt (0 mM, 100 mM and 250 mM NaCl). It was aimed to determine the expression levels of these genes by qRT-PCR by using 4 B and 1 Na related genes which were determined under control and stress conditions from RNA samples obtained as a result of this application. Actin gene, which is optimized for normalization of target genes, was used as reference gene. Within the scope of thesis, expression levels of Gene-21, which may be related to boron, and NHA1 genes, which may be related to salt stress, were measured from the leaf samples of P. distans. As a result of understanding the tolerance mechanism of P. distans at molecular level, it can contribute to the determination of the characters that can be used in molecular breeding in the target plants by understanding the relevant information in different plants that have close affinity relations with high economic value plants such as wheat or these cultivated plants.
vi ÖNSÖZ
Yüksek Lisans eğitim-öğretim hayatıma başladığım ilk günden beri bana yol gösterdiği, bilgi ve tecrübelerini, bulunduğum ortamı ve sahip olduğu çalışma imkanlarını benimle paylaştığı, her zaman bana yardımcı olduğu ve hiçbir zaman desteğini esirgemediği için çok kıymetli saygıdeğer danışmanım Prof. Dr. Erdoğan Eşref HAKKI‘ya sonsuz saygı ve teşekkürlerimi sunarım.
Yüksek lisans tez çalışmamın her aşamasında ve her ayrıntısında bilgi, tecrübe ve deneyimlerini benden hiçbir şekilde esirgemeyen, her zaman yanımda olduklarını hissettiğim, her konuda fikir ve düşüncelerine saygı duyduğum ve önemsediğim çok kıymetli ve değerli Dr. Öğr. Üyesi Anamika PANDEY ve Dr. Öğr. Üyesi. Mohd Kamran KHAN hocalarıma sonsuz teşekkür ederim.
Toprak Bilimi ve Bitki Besleme Bölümü öğretim üyelerinden olan saygı değer hocam Prof. Dr. Sait GEZGİN‘e ve çalışmamın tamamlanmasında yapmış olduğu katkılarından dolayı Doç. Dr. Mehmet HAMURCU‘ya ve çalışma ekibine teşekkür ederim. Ekip ruhu ile bütünleşip bilgi ve desteklerini esirgemeyen laboratuvar arkadaşlarıma, desteklerini esirgemeyen diğer hocalarıma ve bölüm arkadaşlarıma teşekkür ederim.
Ayrıca, hayatımın her aşamasında yanımda olan, beni her zaman destekleyen ve dinamik tutmaya çalışan Sosyolog Tuğba PEKMEZCİ arkadaşıma, çocukluğumdan beri her koşulda yanımda olan dostum Av. Merve BAKAR AMAÇ ile desteklerini her zaman hissettiğim Zir. Yük. Müh. Ayşegül KORKMAZ ve adını sayamadığım tüm arkadaşlarıma teşekkür ederim.
Vatanı ve milleti için ahlaklı ve ilkeli olarak bugünlere gelmemi sağlayan, maddi ve manevi desteğini esirgemeyen ve her koşulda yanımda olan değerli annem Nuray İNANÇ ve babam Hüseyin İNANÇ‘a, her zorlukta ve koşulda beni cesaretlendiren destekçim sevgili kardeşim Seval İNANÇ‘a ve sevgili kardeşim Emin İNANÇ‘a minnettarım.
Çalışmamda; finansman ve burs desteği sağladığı için 116Z343 nolu proje ile TÜBİTAK‘a ve 19201016 nolu proje ile tez çalışmamı maddi anlamda destekleyen ve burs sağlayan Selçuk Üniversitesi Fen Bilimleri Enstitüsü BAP Kordinatörlüğü‘ne teşekkür ederim.
Merve İNANÇ KONYA-2019
vii ĠÇĠNDEKĠLER ÖZET ... iv ABSTRACT ... v ÖNSÖZ ... vi ĠÇĠNDEKĠLER ... vii SĠMGELER VE KISALTMALAR ... ix ġEKĠLLER DĠZĠNĠ ... xi ÇĠZELGE DĠZĠNĠ ... xii 1. GĠRĠġ ... 1 2. KAYNAK ARAġTIRMASI ... 5
2.1. Bor Noksanlığı ve Toksisitesi ... 5
2.2. Bitkide Bor Alımı, Taşınımı ve Tolerans Mekanizması ... 7
2.3. Toprak Tuzluluğu ve NaCl Toksisitesinin Bitkiler Üzerine Etkisi ... 11
2.4. NaCl Tolerans Mekanizması ... 14
2.5. Puccinellia distans (Jacq.) Parl. ... 18
2.6. Tez Çalışmasının Amacı ... 28
3. MATERYAL VE YÖNTEM ... 30
3.1. Bitkinin Yetiştirilmesi ve Uygulamaların Yapılması ... 30
3.2. Yaprak Örneklerinden Total RNA İzolasyonu ... 31
3.3. İzole Edilen Total RNA Örneklerinin Konsantrasyonlarının Belirlenmesi ... 32
3.4. Ters Transkripsiyon (cDNA) ... 32
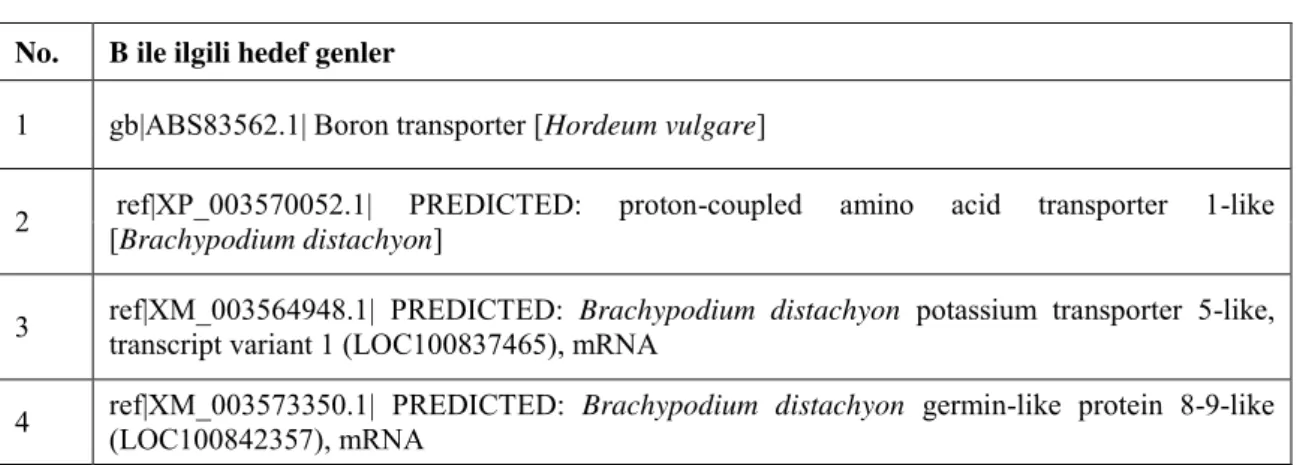
3.5. Kontrol ve Stres Şartları Altında B ve Na İle İlişkili Seçilmiş Hedef Genler ... 33
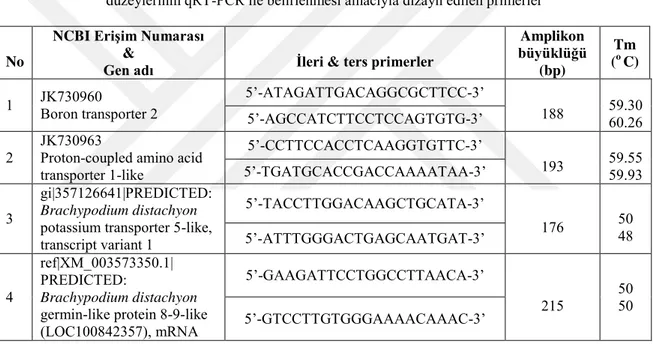
3.6. Genlerin Amplifikasyonunda Kullanılan Primerler ... 34
3.7. Kantitatif Eş Zamanlı Polimeraz Zincir Reaksiyonu (qRT-PCR) ... 35
3.8. İstatistiksel Analiz Metodu ... 36
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 37
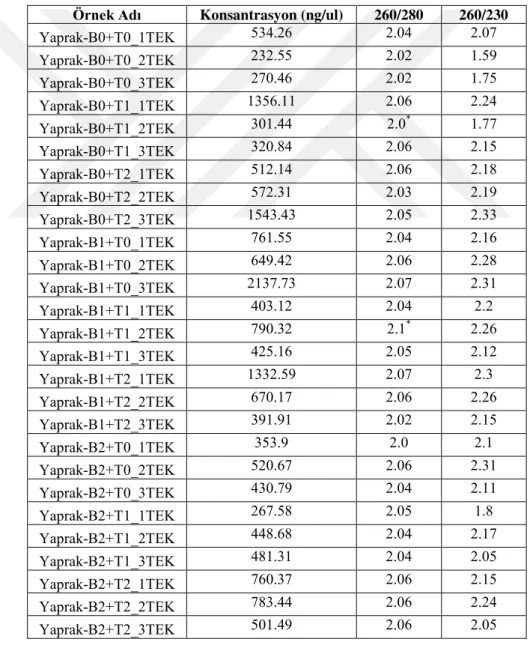
4.1. İzole Edilen Total RNA Örneklerinin Konsantrasyonları ... 37
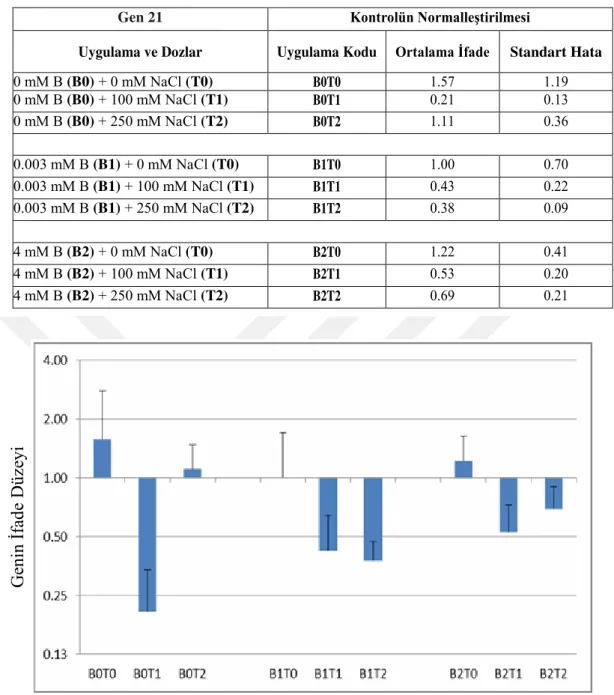
4.2. B-efflux Transporter Proteinlerine Ait Hedef Genlerin qRT-PCR Analizi Sonuçları ... 38
4.2.1. PREDICTED: Brachypodium distachyon potassium transporter 5-like, transcript variant 1 (LOC100837465), mRNA, Gene 21- ... 38
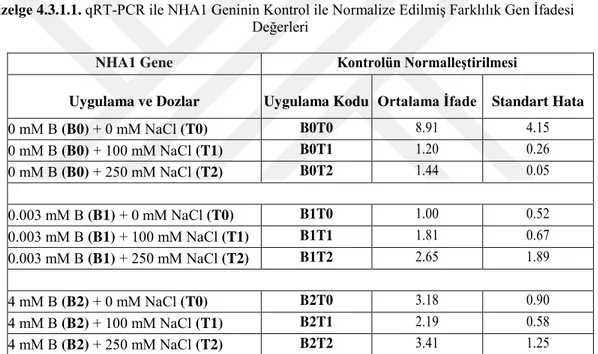
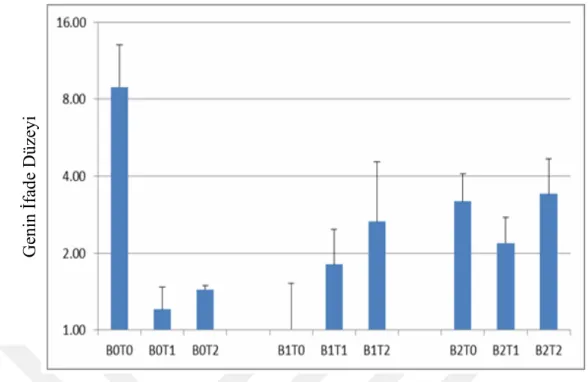
4.3. Na-efflux Transporter Proteinlerine Ait Hedef Genlerin qRT-PCR Analizi Sonuçları ... 41
4.3.1. gb|AY785147.1| Oryza sativa (japonica kültüvar-grubu) Na+ /H+ antiporter (SOS1) GENİ, NHA1- ... 41
viii 5. SONUÇLAR VE ÖNERĠLER ... 44 5.1 Sonuçlar ... 44 5.2 Öneriler ... 45 KAYNAKLAR ... 46 EKLER ... 56 ÖZGEÇMĠġ ... 58
ix SĠMGELER VE KISALTMALAR Simgeler % : yüzde < : küçük > : büyük ≤ : küçük eşittir B : Bor
B(OH)3 : Borik Asit B(OH)4- : Borat B2O3 : Bor Oksit
Ca : Kalsiyum
CaSO4 : Kalsiyum Sülfat
Cd : Kadmiyum
Cl : Klor
dS m-1 : desi Siemens/metre
H : Hidrojen
H3BO3 : Borik Asit
K : Potasyum
KCl : Potasyum Klorür
KNO3 : Potasyum Nitrat
Na : Sodyum
NaCl : Sodyum Klorür (Tuz)
Ni : Nikel o C : Santigrat Pb : Kurşun pH : Hidrojen gücü ppm : milyonda bir kısım psi (b/s) : İnçkare başına libre
Si : Silisyum
x Kısaltmalar
ABD : Amerika Birleşik Devletleri AMF : Arbusküler Mikorizal Mantar APX : Askorbat peroksidaz
AtBOR1 : Arabidopsis thaliana Bor1
B0 : 0 mM B
B1 : 0.003 mM B
B2 : 4 mM B
bç : baz çifti
BOREN : Ulusal Bor Araştırma Enstitüsü
CAT : Katalaz
cDNA : Komplementer Deoksiribo Nükleik Asit DEPC : dietilpirokarbonat
dk : dakika
DNA : Deoksiribonükleik asit
FAO : Food and Agricultural Organisation
GB : glycinebetaine
ha : hektar
HKT : yüksek afiniteli K+taşıyıcıları HvBor2 : Hordeum vulgare Bor2 IAA : indolasetik asit
ICP-AES : Inductively Coupled Plasma-Atomic Emission Spectrometer LCT : düşük afiniteli katyon taşıyıcıları
mg : miligram
mg kg-1 : miligram kilogram mg L-1 : miligram Litre
mL : mililitre
mM : milimolar
mol m-3 : mol metreküp
mRNA : mesajcı RNA
MS : Murashige ve Skoog, 1962
NCBI : National Center for Biotechnology Information NSCC : seçici olmayan katyon taşıyıcıları
PCR : Polimeraz Zincir Reaksiyonu ppm : milyonda bir kısım
qRT-PCR : Kantitatif Eş Zamanlı Polimeraz Zincir Reaksiyonu QTL : Quantitative Trait Loci
RNA : Ribonükleik asit rpm : Revolutions Per Minute
RT-PCR : Reverse Transcriptase - Polymerase Chain Reaction SOD : Süperoksit Dismutaz
T0 : 0 mM NaCl
T1 : 100 mM NaCl
T2 : 250 mM NaCl
TaBor2 : Triticum aestivum Bor2
USDA : Amerika Birleşik Devletler Tarım Bakanlığı WHO : World Healt Organization (Dünya Sağlık Örgütü)
μL : mikrolitre
xi
ġEKĠLLER DĠZĠNĠ
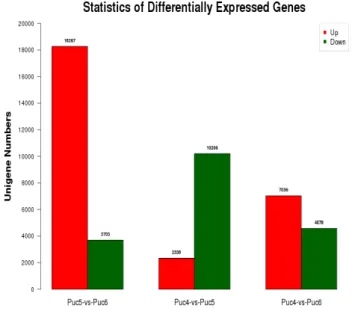
ġekil 2.5.1. Puccinellia distans (Jacq. Parl.)‘a ait sınıflandırma……….18 ġekil 2.5.2. Bor noksan (0 ppm B) koşullar ile Bor toksik (500 ppm B) koşulların kontrol (Hoagland solüsyonu) şartları ile mukayese edildiği ortamda yetiştirilen Puccinellia distans‘ın De-novo sekanslama sonuçlarına göre artış ve azalış gösteren unigenler………..27 ġekil 4.2.1.1. Gen 21 geninin referans gen Actin‘e göre Relatif transkript düzeyi. Relatif ifade düzeyi 3 bağımsız tekrarın ortalamasıdır. Standart sapmaların ortalaması kontrole göre normalize edilmiştir……….39 ġekil 4.3.1.1. NHA1 geninin referans gen Actin‘e göre Relatif transkript düzeyi. Relatif ifade düzeyi 3 bağımsız tekrarın ortalamasıdır. Standart sapmaların ortalaması kontrole göre normalize edilmiştir……….42
xii
ÇĠZELGE DĠZĠNĠ
Çizelge 3.2.1. Bor ve Tuz stresine tabi olan RT-PCR denemesindeki yaprak
örnekleri………...31
Çizelge 3.4.1. RT miks bileşenleri………..33
Çizelge 3.5.1. B ile ilişkili proteinleri kodlayan qRT-PCR çalışmasında kullanılan hedef genler………...33
Çizelge 3.5.2. Na ile ilişkili proteinleri kodlayan qRT-PCR çalışmasında kullanılan hedef genler……….34
Çizelge 3.6.1. B-atılımı (B-efflux) sağlaması muhtemel taşıyıcı proteinleri kodlayan genlerin ifade düzeylerinin qRT-PCR ile belirlenmesi amacıyla dizayn edilen primerler………..34
Çizelge 3.6.2. Na-atılımı (Na-efflux) sağlaması muhtemel taşıyıcı proteini kodlayan genin ifade düzeyinin qRT-PCR ile belirlenmesi amacıyla dizayn edilen primer………...34
Çizelge 3.6.3. qPCR deneylerinde kullanılan referans gene ait dizi bilgisi………35
Çizelge 3.7.1. qRT-PCR Master miksi bileşenleri………..35
Çizelge 3.7.2. PCR reaksiyon döngüsü………...35
Çizelge 4.1.1. P. distans yaprak örneklerinin RNA konsantrasyonları ve saflık değerleri………...37
Çizelge 4.2.1.1. qRT-PCR ile Gen 21‘in Kontrol ile Normalize Edilmiş Farklılık Gen İfadesi Değerleri………..39
Çizelge 4.3.1.1. qRT-PCR ile NHA1 Geninin Kontrol ile Normalize Edilmiş Farklılık Gen İfadesi Değerleri………...41
1. GĠRĠġ
Bitkilerin optimum düzeyde gelişim gösterebilmeleri için bulundukları ortamda yeterli düzey ve oranda bitki besin elementlerinin bulunması ve gerekli koşulların sağlanması gerekmektedir. Normal gelişim süreçlerinde istenmeyen koşullar ile karşılaşıldığı durumlarda bitkiler için stres söz konusudur. Bitkileri etkileyen bu elverişsiz koşullar altında oluşan stres faktörleri ikiye ayrılmaktadır. Bunlar; biyotik (bitkiler, mikroorganizmalar, hayvanlar v.b.) ve abiyotik stres (radyasyon, sıcaklık, su, gazlar, mineraller vb.) etmenleridir (Çulha ve Çakırlar, 2011). Abiyotik stres faktörleri içerisindeki mineral stresinin büyük bir oranını tuzluluk oluşturmakta ve dünyada 9 milyon ha alandan daha fazla alanın tuzluluğa maruz kaldığı bilinmektedir (Tuteja, 2007). Türkiye‘de ise çorak alanların yaklaşık olarak 12 bin ha alanının tuzlu topraklardan oluştuğu bildirilmiştir (Kendirli ve ark., 2005). Bazı kaynaklarda ise Türkiye topraklarının yaklaşık 1.5 milyon ha alanında tuzluluk ve alkalilik sorununun mevcut olduğu ifade edilmektedir (Kanber ve ark., 2005). B toksisitesi ve tuzluluk gibi bitki gelişimi ile verimliliği etkileyen stres etmenleri, kurak ve yarı kurak iklim bölgelerinde genellikle birlikte görülen etmenlerin başında gelen problemlerdendir. Ürün verimliliğini etkileyen B stresine genellikle toprak tuzluluğu eşlik etmektedir. B‘un kolay çözünebilmesi sebebiyle özellikle zayıf drenajlı bölgelerde sodyum tuzları olarak birikir. Bor-tuz sorunu çoğunlukla bitkilerde yüksek düzeyde B ve tuz içeren sulama suyu ile sulama veya doğal olarak B ve tuz bakımından zengin topraklarda bitkilerin büyümesi nedeniyle birlikte görülmektedir (Marschner, 1995). Bu nedenle, B toksisitesi ile tuzluluk problemlerinin yoğun olduğu arazilerde kaliteli ürün elde edilmesi ve iyi bir tarım yapılabilmesi için bu arazilerin ıslah edilmesi gerekmektedir.
Dünya bor rezervlerinin dağılımına bakıldığında (%B2O3) Türkiye %73.4, Rusya %7.7, ABD %6.2‘lik bir paya sahip olduğu bilinmektedir. Dünya bor rezervlerinin %73.4‘üne sahip olan ve ilk sırada yer alan Türkiye‘de bor yatakları; Eskişehir/Kırka, Kütahya/Emet, Balıkesir/Bigadiç, Bursa/Kestelek‘te bulunmaktadır. Ayrıca, Türkiye topraklarının %31.1‘inde yeterli seviyede B bulunurken, %22.7‘sinde toksik veya fazla, %46.2‘sinde ise çok az ve yetersiz düzeyde B bulunduğu tespit edilmiştir (BOREN, 2017). B toksisitesi sadece tarım alanlarının aşırı ve dengesiz gübrelenmesinin sonucu olarak değil aynı zamanda yeraltı bor rezervlerinden de kaynaklanmaktadır. Türkiye
topraklarının dünya bor rezervleri arasında %73.4‘lük bir paya sahip olması ekonomik açıdan faydalı olmasına rağmen tarımsal alanlarda büyük sorunlara neden olmaktadır.
Dünya topraklarının çoğunluğunda ülkemiz dahil noksanlığı gözlenen B elementinin, ülke topraklarında toksisitesi de gözlenmektedir. Topraklarda bulunan B elementi, gerek mevcut rezervler gerekse yeraltı su kaynakları ve bor madenlerinin etrafındaki sulama suyu ile toprak yüzeyine taşınmasıyla bitkiler üzerindeki etkileri artmaktadır (Stiles ve ark., 2010). B elementinin fazlalığı ile toksisite artmakta ve bitkilerin büyüme ve gelişmesi için sınırlandırıcı bir faktör olarak Orta Anadolu topraklarında önemli bir mikro besin elementi problemi olarak karşımıza çıkmaktadır (Cakmak ve ark., 1997).
Tüik (2018) verilerine göre, Türkiye tarım topraklarının nadas alanları hariç 15.421 milyon hektar alanın tahıl ve diğer bitkisel ürünlerin ekimi için kullanılmakta olduğu bilinmektedir. Bu alanın da yaklaşık olarak 10.8 milyon hektarında hububat ekimi yapılmakta ve hububat ekim alanı içerisinde ilk sırada buğday yer almaktadır. Türkiye, dünya buğday ekim alanın %3.5‘ine sahiptir. Ayrıca, buğday ülkemizde en fazla ekim alanına sahip olup insan beslenmesinin esas kaynağı olan ve günlük enerji kaynağının büyük bir kısmının karşılandığı kültür bitkilerinden olduğu bilinmektedir. Türkiye tarım topraklarında buğday üretiminin en fazla gerçekleştirildiği Orta Anadolu tarım arazilerinde B toksisitesi verimi önemli ölçüde düşürmekte ve kısıtlamaktadır.
Gezgin ve ark. (2002)‘nın yaptığı bir çalışmada Aksaray, Afyon, Karaman, Kayseri, Konya, Niğde ve Nevşehir il topraklarının B içeriği incelenmiştir. Bu inceleme sonucunda Orta Anadolu tarım topraklarının önemli bir kısmı (%18‘inde) özellikle tahıllar için kritik değer olarak kabul edilen 3 mg kg-1‘dan fazla B içerdiği rapor edilirken önemli bir kısmında da bor noksanlığı olduğu belirlenmiştir. Verimi yüksek oranda düşüren B toksisitesi sorununun çözümü için yapılacak olan çalışmalar doğrultusunda insan beslenmesinde büyük etkisi olan tahıl grubu buğday bitkilerinin verimliliğinin artırılması gerekmektedir.
B elementi noksanlığının 80‘den fazla ülkede 132 adet bitkide gözlendiği ve dolayısıyla yaygın bir tarımsal problem olarak karşımıza çıktığı rapor edilmiştir (Shorrocks, 1997). Bor elementi hem düşük hem de yüksek konsantrasyonlarda bitkinin
vejetatif organlarının yanında generatif organlarının da etkilenmesine, bitkide verim ve kalitenin düşmesine sebep olmaktadır. B eksikliği genellikle ilk olarak hızlı büyüme gösteren dokularda görülmekte ve büyümede azalmaya ve ayrıca steriliteye neden olmaktadır (Dell ve Huang, 1997). Ancak bitkilerin gereksinim duyduğu B miktarı oldukça sınırlı bir aralığa sahiptir. Bu yüzden, B gübrelerinin uygulanmasında oldukça dikkatli olunmalıdır (Marschner, 1995).
B, toprakta ve yer altı sularında doğal olarak bulunur ve birikebilir (Nable ve ark., 1997). Yüksek B içeren materyaller ile gübreleme sonucunda da B birikimi gerçekleşebilir. Ayrıca, B içeren atık materyalleri depolamak amacıyla kullanılmış topraklarda B birikimi ortaya çıkabilir (Soy, 2002). Sulama suyu, kuyu suları, drenaj suyunun toprağa uygulanması, endüstriyel atıklar ve kimyasallar gibi faktörler B toksisitesinin başlıca sebeplerindendir (Dhankhar ve Dahiya, 1980). Bunların yanı sıra çürümüş organik madde içeren gübrelerin uygulanması, kompost gübrelerinin aşırı kullanılması, fosil yakıtların yanması ve toprak pH‘sını arttırmak amacıyla yüksek oranda B uygulamasının yapılması yüksek B konsantrasyonlarının oluşmasına sebep olmaktadır (Türkan ve Öncel, 2006)
B‘a toleranslı bitkiler geliştirmek için öncelikle böyle bir strese karşı olan tolerans mekanizmalarının anlaşılması gerekmektedir. Daha önceden yapılan bazı çalışmalarda farklı bitki türlerinin farklı tolerans mekanizmaları geliştirdikleri belirlenmiştir. Bazı bitkiler köklerinde B birikimini sınırlandıracak mekanizmalara sahiptirler. Bu gibi bitkiler aynı zamanda B‘a dirençli bitkiler olarakta adlandırılmaktadır. Hordeum vulgare L. ve Triticum aestivum L. türü bitkilere ait genotiplerin B‘u dışlayabildiği rapor edilmiştir (Nable, 1988; Nable ve Paull, 1990; Nable ve ark., 1997). Bunun yanı sıra bazı bitkiler ise B‘ u dokularından dışlamaz, güvenli bir şekilde biriktirerek tolere ederler. Bu nedenle bu gibi bitkiler bor akümülatör bitkiler olarak adlandırılırlar. Stiles ve ark. (2010)‘nın Puccinellia distans ve Gypsophila arrostil ile yaptığı bir çalışmada, B alımının köklerde değişmediği ve bitkilerin B‘u yaprak dokularında toksik etkileri gözlemlenmeksizin depolayabildiği rapor edilmiştir. Aynı zamanda bu bitkiler yaprak dokularında B biriktirme ve depolama konusunda çok iyi olduklarından B akümülatör bitkileri olarak adlandırılmıştır. Tarımı yapılmakta olan birçok bitkiye göre buğdayın (özellikle de ekmeklik buğdayın)
mukayeseli olarak, tuzluluğa toleransı daha yüksek olmakla birlikte, genel olarak düşünüldüğünde, yerel Türk buğday çeşitlerinin tuza tolerans düzeyi yetersizdir.
Temel besin kaynağı olarak tüketilen ve tahıllar grubu içerisinde önemli bir yere sahip olan buğday bitkisi, insan beslenmesinde önde gelen enerji kaynaklarından birisi olarak bilinmektedir. Dünya‘da ve ülkemizde artan nüfus göz önüne alındığında insan beslenmesindeki önemi giderek artmaktadır. Artan ihtiyacın karşılanması için günümüzde çeşitli ıslah programları pratik ve ekonomik bir yöntem olarak karşımıza çıkmaktadır. Ancak buğday kompleks bir genom yapısına sahip poliploid bir bitkidir ve aynı familyada yer alan daha az karmaşık ve model özelliğine sahip bir bitkinin kullanılması bilgi birikiminin hızlanmasını sağlayacaktır. Puccinellia distans bu özelliklere sahip olmasının ötesinde tez konusu B toksisitesi ve tuz stresine karşı çok yüksek düzeyde dayanıma da sahiptir. Bu nedenle çalışmaların öncelikle bu bakir bitkide yürütülmesi araştırmaların özgünlük kazanmasının ötesinde mekanizmaların sınırlarının belirlenmesinde de belirleyici olacaktır. Bu bitkinin yüksek B ve tuz tolerans mekanizmasında görev aldığını düşündüğümüz B- ve Na- efflux proteinleri hakkında daha önce yapılmış herhangi bir çalışma bulunmamaktadır.
2. KAYNAK ARAġTIRMASI 2.1. Bor Noksanlığı ve Toksisitesi
Mikro elementlerin eksikliğinde veya fazlalığında tüm bitkiler strese girmektedirler. Strese giren bu bitkilerde noksanlık veya toksisite belirtileri gözlemlenir.
Bitkilerde gözlenen bu besin elementi noksanlığı veya toksisitesi bitki gelişimini ve ürün verimini önemli ölçüde etkilemektedir. B, bitkilerin gelişebilmesi için gerekli olan mikro besin elementlerinden birisi olarak bilinir (Marschner, 1995). Bitkilerde noksanlık ve toksisite belirtilerine neden olan toprak B seviyeleri birbirine oldukça yakındır (Chapman ve ark., 1997). Bu nedenle bilinen mutlak gerekli mikro besin elementlerinin arasında noksanlık ve toksisite belirtileri en yaygın olarak görülen elementlerin başında gelmektedir. Ülkemiz topraklarının en az %25‘inde eksikliği görülürken, %20‘sinin üzerinde ise toksisitesi görülmektedir (Gezgin ve ark., 2002).
Bergmann (1992), aynı toprakta yetişen bitki türleri arasında B gereksinimleri bakımından farklılıklar gözlemlendiğini belirtmiştir. Örneğin, buğday gibi Gramineae familyasındaki monokotil türler için 5-10 mg kg-1
kritik seviye iken, yonca gibi dikotiledon türlerde ise 20-70 mg kg-1
kritik aralık olarak belirtilirken, lateks üreten bitkilerde 80-100 mg kg-1kritik aralık olarak rapor edilmiştir.
B toksisitesine sebep olan etmenlerin başında B içeriği yüksek sulama suyunun kullanımı, yüzey gübrelemesi, endüstriyel atıklar ve kimyasallar gelmektedir (Harite, 2008). B elementinin toprakta 5 mg kg-1‘dan fazla bulunması ve sulama suyunun 1-3 mg L-1 arasında B içerdiği koşullarda yetişen bitkiler için toksik etkiye yol açmaktadır. Tarım alanlarında B içeriği yüksek sulama sularının kullanılması B elementinin toprakta tutulmasına ve tarım alanlarında B toksisitesi etkilerinin gözlenmesine sebep olmaktadır (Cartwright ve ark., 1984). Dolayısıyla gübreleme yapılırken sulama suyunun B içeriğine dikkat edilmesi gerekmektedir (Karaman, 2012).
Fazla B içeren kurak ve yarı-kurak iklimin hakim olduğu bölge topraklarında yetiştirilen özellikle tahıl ve kültür bitkilerinde B toksisitesi çok yaygın bir beslenme sorunu olarak karşımıza çıkmaktadır. Örneğin, Güney Avustralya'da kurak ve yarı kurak
bölgelerden alınan bitki ve toprak örneklerinde çok yüksek B konsantrasyonlarına rastlanmıştır (Cartwright ve ark., 1984). Kültür bitkilerindeki B noksanlığı, diğer iz elementler ile karşılaştırıldığında daha yaygın olarak karşımıza çıkmaktadır. Topraktaki B noksanlığı, gübreleme ile toprağın zenginleştirilmesi gibi kolay yöntemlerle giderilebilmesine rağmen gübrelerin dikkatsiz bir şekilde kullanılması bitki gelişim ve verimini olumsuz bir şekilde etkileyen B toksisitesine sebep olabilmektedir. B toksisitesi ile mücadele etmek noksanlık ile mücadeleye göre oldukça zor ve masraflıdır. Aşırı B içeren toprakların ıslah edilmesinde, tuzlu toprakların ıslahına oranla 3 kat daha fazla su kullanılmasına ihtiyaç duyulmaktadır (Keren ve Bingham, 1958). B içeriği düşük sulama suları ile toprağı yıkayarak fazla B‘dan arındırma uygun bir yöntem olmasına rağmen uzun vadede kalıcı ve ekonomik bir çözüm oluşturmaz. Bunun için B toksisitesine dayanıklı çeşitlerin geliştirilmesi, B toksisitesi ile mücadelede kullanılabilecek en etkin ve en faydalı yöntemdir.
B toksisitesi ile mücadelede toksisiteye dayanıklı çeşitlerin seçimi ve ıslahının yapıldığı bilinmektedir. Bunun yanı sıra B‘u bünyesinde tutan ve bazı dokularında depolayan bitkilerin yetiştirilmesi ve geliştirilmesi, toprakta bulunan mevcut B‘u bağlayan organik madde içeriği yüksek olan materyallerin toprağa ilavesinin sağlanması ve B elementinin bitkilerce alımını sınırlayan elementleri içeren gübreler ile gübrelemek Gezgin ve ark. (2005) tarafından önerilen bir yöntem olarak karşımıza çıkmaktadır.
Dell ve Huang (1997)‘a göre bor noksanlığı koşullarında bitki kök ucunun gövdeye oranla gelişimi azalmakta, dolayısıyla gövde/kök oranı artması nedeniyle bitkinin stres koşullarına (diğer bitki besin elementleri veya su noksanlığı vb.) hassasiyeti önemli düzeyde artmaktadır. Bunların yanı sıra, yaprak büyümesi, kök uzaması gerilemekte veya durmakta, çiçek gelişimi, meyve ve çiçek oluşumu bozulmakta ve tohum verimi de düşmektedir.
B noksanlığı monokotil bitkilerde erkek kısırlığına neden olurken, dikotil bitkilerde ise dişi kısırlığına neden olmaktadır. Bu durum bitkide meyve veriminin düşmesi ile sonuçlanmaktadır (Gupta, 1993).
B‘un farklı bitki çeşitlerinde yapısal, fizyolojik ve biyokimyasal olaylarda önemli rol oynadığı düşünülmektedir. B, lignifikasyon olgusunda, hücre duvarı sentezi
ile yapısının oluşumunda, hücre bölünmesi, uzaması ve bitkide büyüme açısından önemlidir. Ayrıca, bitkilerde karbonhidrat metabolizmasında, solunumda, RNA (ribonükleik asit) metabolizmasında, IAA (indolasetik asit) metabolizmasında, fenol metabolizmasında görev alırlar. Bitki içerisindeki şekerin hareketine yardımcı olur. Polen çimlenmesinde ve polen tüpü büyümesinde biyolojik membranların yapısal ve fonksiyonel özellikleri üzerinde önemli rollere sahip olduğu ileri sürülmektedir (Parr ve Loughman, 1983). B elementinin bitkideki işlevleri üzerine çok sayıda araştırma yapılmış olmasına rağmen bitkideki etkileri henüz netlik kazanmış değildir (Demirtaş, 2005).
Nükleik asitlerin sentezi için önemli bir element olan B‘un eksikliği durumunda protein sentezi olumsuz bir şekilde etkilenir. B noksanlığında deoksiribonükleik asit (DNA) sentezinin, ribonükleik asit (RNA) miktarının ve hücre bölünmesi ile kök uzamasının da önemli düzeyde azaldığı tespit edilmiştir (Moore ve Hirsch, 1983). Dolayısıyla B uygulamasının protein sentezini artırması beklenir.
B elementinin fazla olduğu durumlarda ise hücre bölünmesi etkilenir ve özellikle kök büyümesi azalır. Ayrıca, yaprakların klorofil içeriğinde azalmaya sebep olur. B gövdede biriktiği için genellikle yaprak tabanından başlayıp yaprak ucuna doğru devam eden kloroz ve yaprak kenarlarında nekrozlara sebep olur. Ayrıca, verimde ve meyve kalitesinde düşüşlere neden olmaktadır (Nable ve ark., 1997; Reid ve ark., 2004).
2.2. Bitkide Bor Alımı, TaĢınımı ve Tolerans Mekanizması
Bitkilerde B alımı, topraktaki B konsantrasyonu ve B‘un bitkiler açısından elverişliliği ile yakından ilişkilidir. Bitkilerin B gereksinimlerini karşılama ve bünyelerine alımda toprak pH‘ı, nemi ve sıcaklığının da etkili olmasının yanı sıra tekstür, kil mineralleri ve organik madde gibi toprak faktörleri de etkilidir (Goldberg, 1997). Bitkinin B alımında, toprak özelliklerinin yanı sıra, bitkisel yapı farklılıkları ve iklim gibi bazı faktörlerin de etkilediği bilinmektedir (Güzel ve ark., 2002).
pH‘ya göre değişiklik gösteren B, asidik pH seviyerinde düşük adsorpsiyon gösterirken, alkalin karakterli topraklarda ise maksimum adsorpsiyon göstermektedir. Bu durum kireçli topraklarda yetişen kültür bitkilerinde gözlenen B noksanlığının
sebebidir. pH arttıkça sulama suyundaki borat ve borik asit iyonlarının oranları değişmektedir. Nötr pH‘dan sonra baziklik arttıkça borat iyonları artış gösterirken, borik asit iyonlarında azalma gerçekleşmektedir (Keren ve Bingham, 1958). Yani, toprağın pH‘sı arttıkça, bitkilerin topraktan B alım düzeyi genelde düşmektedir (Gupta, 1993). Bu durum, yüksek pH koşullarında B‘un kalsiyumla birleşerek kalsiyum boratlar halinde çökelmesiyle açıklanabilir.
B alımı bitkiler arasında ve aynı tür içinde genotipler arasında da önemli düzeyde farklılık göstermektedir. B ihtiyaç durumuna göre bitkileri 3 gruba ayrılmıştır: 1. B‘a en az ihtiyaç duyan (monokotil bitkilerden buğdaygiller familyası)
2. İlk gruba göre daha fazla B‘a ihtiyaç duyan (buğdaygiller familyası dışında kalan monokotil ve dikotil bitkiler)
3. B‘a en fazla ihtiyaç duyan lateks üreten bitkiler (Mengel ve ark., 2001). Genellikle tek çenekli (monokotil) bitkilerin B alım kapasitelerinin çift çenekli (dikotil) bitkilere göre daha az olduğu bilinmektedir (Tanaka, 1967).
Bitkilerde B‘un taşınması türler arasında değişkenlik gösterebilmektedir. Temelde besin elementlerinin taşınması ksilem ve floem aracılığı ile iletim demetlerinde gerçekleştiği bilinmektedir. İletim demetlerinde taşınmayı sağlayan anahtar molekül ise sudur. B elementinin kökten gövdeye doğru taşınması su akımının yardımıyla ksilem cansız hücrelerinde gerçekleşmektedir. Bitki yapraklarında gerçkeleşen su kaybı ile birlikte bir akım gücü meydana gelmektedir. Öncelikle ksilem translokasyonu, bitkide transpirasyonun en yoğun olduğu bölgeye, genellikle yapraklara doğru olmaktadır. Floem translokasyonu ise transpirasyondan tamamen bağımsız olarak gerçekleşmektedir. Floem akım yönü ise genç yapraklar ve kolayca su kaybına uğramayan meyve ile tohumlara doğru, büyüme bölgelerinde gerçekleşmektedir (Brown ve Shelp, 1997).
B'un bitki organlarında hareketliliğinin immobil olduğu düşünülmekteydi. (Marschner, 1995). Ancak, elma gibi fotosentez ürünü şeker alkolü olan bitkilerde ise B‘un taşınmasında transpirasyon akımı aktif rol oynamamıştır. Yaprakta şeker alkolü miktarının artması ile B-polyol kompleksleri oluşmakta ve bunun sonucunda da B‘un bitkide kolayca hareket ettiği rapor edilmiştir. B, sorbitol ve mannitol şeker alkolleri ile
B-polyol kompleksleri oluşturabilmekte ve böylece şeker alkollerince zengin olan bitkilerde B kolayca taşınmaktadır (Brown ve Shelp, 1997).
B‘un bitkilerce kullanılabilen formu toprakta çözünebilen, bağımsız, iyonize olmamış borik asit (H3BO3), borik asit (B(OH)3-) veya iyon halindeki borat (B(OH) -4)‘dır (Hu ve Brown, 1997). Bitkilerce B temelde borik asit (H3BO3) ve B(OH)3 -formunda pasif yolla alınmasının yanı sıra çok daha az oranda da olsa B(OH)
-4 formunda aktif yolla da alındığı bilinmektedir. Bitki kök bölgesine taşınımda ise yüksek oranda (%65) kitle akışı ve daha düşük oranda da (%32) difüzyon rol oynadığı bildirilmiştir (Güzel ve ark., 2002). B‘un bitki köklerine yüksüz haldeki ayrışmamış borik asit şeklinde girdiği bilinmektedir (Woods, 1996). Borik asitin toprak solüsyonunda ayrışmamış halde olması sebebiyle bitkiler kökleri aracılığı ile pasif taşınım ile alabilmektedir (Hu ve Brown, 1997). Ayrışmamış ve yüksüz durumda olan borik asit suda ve lipitte çözünebilir olduğu için bitki hücrelerinden geçirgenlik özelliğine sahiptirler (Mengel ve ark., 2001).
B çoğunlukla pasif yolla taşındığı için yaşlı yapraklardan genç yapraklara B elementinin taşınımı kolayca gerçekleşememektedir. Bitkilerin yeterli düzeyde besin elementlerini alımı, verim ile birlikte kaliteli ürün elde etmek için önemlidir. Bitkilerin besin element alımında noksan, yeterli veya toksik düzeylerden bahsederken net rakamlar yerine aralık belirtilir. B ihtiyaç aralığı ise bitkilerin organlarında farklılık gösterebilmektedir. B‘un yetersiz alındığı durumlarda bitkilerin vejetatif gelişiminde herhangi bir etki görülmemesine rağmen, meyve ve tohum gibi generatif organların gelişimlerinin daha fazla etkilediği bilinmektedir (Mengel ve ark., 2001).
Bitkinin ihtiyacı olan B elementinin noksan olduğu durumlarda, bitkinin bor gereksinimini karşılayan B taşıyıcılarının (B taşınımını sağlayan membran proteinlerinin) varlığı dikkat çekmektedir. B hareketliliğinden sorumlu genler ilk olarak model organizma olan Arabidopsis thaliana yoluyla tanımlanmıştır (Lin ve ark., 1999). A. thaliana bitkisinde B‘un kök yüzeyinden gövdeye iletilmesinden sorumlu en az 2 adet membranlardan taşınım mekanizması olduğu bilinmektedir. İlk olarak B, epiderma, korteks ve endoderma hücrelerine taşınmakta (alınım) ve daha sonra perisikl veya ksilem parankima hücrelerinden öz bölgesindeki hücrelerin apoplastına iletildiği (salınım veya ksileme yükleme) şeklinde olduğu rapor edilmiştir. B noksanlığına son
derece duyarlı olan BOR1-1 mutantının analiz edilmesiyle yüksek afiniteli bir taşınım sistemi olarak ksileme boşaltımda rol alan BOR1 tanımlanmıştır (Noguchi ve ark., 2000).
Reid ve ark. (2004)‘nın yaptığı bir çalışmada arpanın B-tolerans mekanizmasının köklerden B atılımı şeklinde olduğunu ileri sürmüşlerdir. Bu ve bunun gibi mevcut çalışmalar B toleransının oluşumundan sorumlu köklerde B atılımını sağlayan taşıyıcıların varlığına işaret etmektedir. A. thaliana‘da ilk defa tanımlanan BOR1 taşıyıcı proteini, sınırlı B bulunan ortamlarda köklerden gövdeye doğru etkin bir şekilde B taşınımını sağlayan ve mayada ifade edildiği zaman hücre içi B miktarını azaltabilen efflux (atılım) yapan bir taşıyıcı olarak tanımlanmıştır (Takano ve ark., 2002). Bunun yanında (Arabidopsis thaliana) AtBOR1‘in veya mayadaki homoloğu olan BOR1‘in mayada aşırı ifadesi hücrelerdeki B içeriğini düşürerek yüksek B toleransı sağlar (Nozawa ve ark., 2006; Takano ve ark., 2007). AtBOR1‘in bitkilerde aşırı ifadesi ise yüksek B toleransı sağlamadığı fakat düşük B toleransı sağladığı belirtilmiştir (Miwa ve ark., 2006). Bu durum BOR1‘in yüksek B dozlarında parçalanması ve böylece köklerden gövdeye doğru B taşınımının sadece düşük B dozlarında gerçekleşmesinin teşvik edilmesinden kaynaklandığı şeklinde açıklanmıştır (Takano ve ark., 2005; Miwa ve ark., 2006).
B-(efflux) atılımını sağlayan bir transporter kullanıldığı taktirde bitkilerde yüksek B toleransı kazandırılabilinir. Bunun için A. thaliana genomunda yer alan 6 adet BOR1 benzeri genlerden (Nakagawa ve ark., 2007) BOR4‘ün yüksek B dozlarında kararlı bir şekilde ifade olduğu belirlenmiştir (Miwa ve ark., 2007). Benzer şekilde (Sutton ve ark., 2007) 2 arpa kültivarı arasında B toleransı bakımından QTL haritalaması yaptıklarında haritalanan bölgede BOR1 homoloğu olan Bot1‘i tespit etmişlerdir. Toleranslı arpa kültivarında Bot1 mRNA seviyelerinin hassas olan kültivara göre daha yüksek olduğu bulunmuştur. Reid (2007), BOR1 homologları olan arpada (Hordeum vulgare) HvBor2 ve de buğdayda (Triticum aestivum) TaBor2 genlerini klonlamış ve arpa ve buğdayın köklerinde yüksek düzeylerde mRNA tespit etmiştir. Ayrıca mRNA seviyeleri ile köklerdeki B konsantrasyonlarındaki azalma ve bitkilerin büyümelerindeki artış arasında pozitif bir korelasyon olduğunu rapor etmişlerdir. Bu sonuçlar, BOR1 homologlarının köklerdeki B konsantrasyonlarını düşürerek yüksek B toleransı sağladıklarına dair rollerini desteklemektedir. Arpa ve buğdayın gövdesindeki
Bor2 mRNA seviyeleri köklerden daha düşük miktardadır. Kök B konsantrasyonlarındaki azalmanın yüksek B toleransı sağlamada yarattığı farklılık, Miwa ve Fujiwara (2011) tarafından BOR4‘ü gövdesinde aşırı ifade eden A. thaliana‘nın gövdesinde de incelenmiştir. Gövdede aşırı BOR4 ifade eden transgenik A. thaliana bitkilerinin toksik B konsantrasyonlarına maruz bırakıldığında yapraklarındaki B dağılımını, sitoplazmadan apoplasta doğru B atılımı sağlamak suretiyle düzenleyerek yüksek B toleransı geliştirdikleri görülmüştür (Miwa ve Fujiwara, 2011).
Buğday ve arpanın yapraklarında yüksek B birikimine karşı farklı duyarlılıklar rapor edilmiştir. Reid ve Fitzpatrick (2009), toleranslı arpa ve buğday kültivarlarının yapraklarındaki nekrotik bölgelerin B konsantrasyonlarının, duyarlı kültivarlara göre daha yüksek miktarda olduğunu bulmuşlardır. Bu gözlem, yüksek B toleransının dokudaki B konsantrasyonuyla doğrudan ilişkilendirilemeyeceğini ve gövde dokularının B-toksisitesine karşı tolerans mekanizmalarının olduğunu göstermektedir.
2.3. Toprak Tuzluluğu ve NaCl Toksisitesinin Bitkiler Üzerine Etkisi
Bitkisel üretimi sınırlandıran en önemli abiyotik stres etmenlerinden birisi de tuzluluk problemidir. Tuz stresi; toprakta ya da suda bulunan NaCl ve diğer çözünebilir tuz miktarının artışına bağlı olarak bitkinin gelişimini ve büyümesini olumsuz yönde etkilemektedir. Söz konusu bu tuzlar; genelde klorürler, sülfatlar, karbonatlar, bikarbonatlar ve boratlar olarak bilinmektedir. Fakat doğada sıklıkla karşılaşılan tuz formu NaCl‘dür (Dölarslan ve Gül, 2012). Çeşitli çözünebilir tuzların çok yüksek konsantrasyonlarını içeren ortamlarda yaşamlarını sürdürebilme yeteneğine sahip bitkiler ise ‗tuza toleranslı bitkiler‘ olarak bilinmektedirler. Salicornia herbacea, Atriplex vericaria, Suaeda maritima gibi halofitler genellikle küçük yapılı olup, yüksek tuz şartlarına adapte olmuş ve bu şartlarda yaşamını sürdürebilen bitkilerdir (Yilmaz ve ark., 2011; Dölarslan ve Gül, 2012).
Ülkemizde 1,5 milyon ha alanda tuzluluk probleminin var olduğu bilinmekte (Uras ve Sonmez, 2010) ve ayrıca Orta Anadolu tarım kuşağında tuz sorunu büyük bir problem olarak karşımıza çıkmaktadır. Tuzluluk problemi bitkilerin yetiştirilmesine engel olmakta ve ileri seviyedeki tuzluluk bitkilerde ölüme neden olmaktadır.
Ülkemizde ve dünya topraklarında kültür bitkilerinin gelişimini ve büyümesini önemli ölçüde engelleyen ve sınırlandıran çözünebilir tuzlar, değişebilir Na veya her iki unsuru da içeren problemli topraklar ile yaygın bir şekilde karşılaşılmaktadır (Bahtiyar, 2002). Topraklarda bulunan fazla miktarlardaki değişebilir Na, toprağın su geçirgenliğini ve havalanmasının azalmasına sebep olarak bitki gelişimini olumsuz yönde etkilemekte ve engellemektedir. Bu gibi tuzluluk sorunun olduğu topraklarda bulunan mevcut tuzların cinsi ve miktarına bağlı olarak, kültür bitkilerinde çimlenme, büyüme ve ürün miktarında azalma gözlenmektedir (Richards ve Allison, 1954; Dizdar, 1978).
Tuz stresinin bitkiler üzerindeki etkileri; bitkinin çeşidine, uygulanan tuz çeşidi ile miktarına ve maruz kalma süresine bağlı olarak değişmektedir. Tuzlu ortamlarda bitkiler genotipik farklılıklara bağlı olarak çok farklı cevaplar verirler (Dajıc, 2006). Tuzluluğa karşı verilen bu farklı büyüme cevapları sadece farklı bitki türleri için değil aynı türün farklı çeşitleri için de geçerlidir (Munns, 2002).
Bitkiler sodyumu, Na+ iyonu şeklinde alırlar. Aldıkları Na miktarı ve Na‘a tepkileri yönünden bitkiler iki gruba ayrılmaktadır. Bunlar:
1. Natrofilik bitkiler, Na seven bitkiler olup Na‘u kolaylıkla alırlar. Na bitki organlarında üniform bir şekilde dağılır.
2. Natrofobik bitkiler ise Na‘u sevmeyen bitkilerdir ve bu bitkilerde Na kökte toplanır (Karaman, 2012).
Siegel ve ark. (1980), tuz stresi koşullarında bitkilerde aşırı miktarlarda biriken Na, K (Potasyum)‘un alımını engelleyerek bitkilerin iyon dengesinde bozulmalara neden olabileceğini bildirmiştir. Tuzlu şartlarda yetiştirilen buğdayda tuza dayanıklılık kriteri olarak bitkinin Na+ ve K+ içeriği baz alındığı bilinmektedir (Ahsan ve Khalid, 1999). Bitki bünyesinde gerçekleşen K konsantrasyonunun artışı, bitkinin tuza karşı dayanaklılığının artmış olmasının göstergesi olarak bilinmektedir (Hsiao ve Lauchli, 1986). Ayrıca, bitkinin sahip olduğu K+/Na+ oranının yüksek olması tuza dayanıklılığı ile doğru orantılı olduğu bildirilmiştir (Gorham, 1990; Ashraf ve Khanum, 1997; Yağmur ve ark., 2006).
Karanlık ve ark. (1999)‘ı 20 farklı ekmeklik ve makarnalık buğday genotipini kullanarak NaCl tuzuna karşı daynıklılıkları yönünden bitkilerin Na (Sodyum), K (Potasyum), Ca (Kalsiyum) içeriklerinin yanısıra K/Na ve Ca/Na oranı gibi parametreleri de değerlendirmiş ve NaCl uygulamasından verim kaybının en az Dağdaş (ekmeklik buğday) ile en çok verim kaybının ES-14 (ekmeklik buğday) ve Kunduru (makarnalık buğday) genotiplerinde gerçekleştiğini rapor etmişlerdir.
Harmer ve ark. (1953)‘ı bitkileri Na‘a karşı gösterdikleri tepkimeye göre dört grupta toplamışlardır. Bunlar:
a. K noksanlığında Na‘dan çok fazla yararlanan bitkiler; Yonca, Arpa, Yulaf, Domates, Brüksel Lahanası ve Havuç
b. K noksanlığında Na‘dan az yararlanan bitkiler; Mısır, Kırmızı Üçgül, Marul, Soğan, Patates
c. Yeterli düzeyde K‘un bulunması durumunda Na‘dan az yararlanan bitkiler; Buğday ve Bezelye
d. Yeterli düzeyde K‘un bulunması durumunda Na‘dan çok fazla yararlanan bitkiler; Kereviz, Şalgam ve Pancar
Na ile ilgili yapılan çalışmalarda bazı C-4 bitkileri (mısır ve şeker kamışı gibi bitkiler dışında) ile belli koşullar altında CAM bitkileri, mavi-yeşil algler, bazı bakteri ve mantarlar için mutlak gerekli bir besin elementi olduğunu bildirmesine rağmen C-3 bitkileri için mutlak gerekli besin elementi olmadığını bildirmiştir (Asher, 1991).
Brownell (1968) ‘in yapmış olduğu bir çalışmaya göre, besin çözeltisinde Na konsantrasyonu ≤ 100 μM olduğu zaman C-4 bitkilerinde maksimum gelişme gözlenirken, C-3 bitkilerinin Na konsantrasyonu 0.07 μM düzeyine indirilmiş besin çözeltilerinde optimum düzeyde gelişme gösterdikleri gözlenmiştir.
Hem dünya genelinde, hem de ülkemizde buğday yetiştirilen alanlarda büyük oranda tuzluluk problemleri ile karşılaşılması söz konusu olduğuna göre, gerek halihazırda atıl durumda kalmış tarım potansiyelli alanlar, gerekse, yakın gelecekte bu riski taşıyan geniş alanlar düşünüldüğünde, bu ortamların kaderine terkedilmesi kabul edilebilir değildir. Tuzluluk, bitki büyümesini, bitkilerin ürün kalitesi ve verimliliğini önemli ölçüde düşüren ve sürekli artan bir sorun olarak karşımıza çıkmaktadır. Tuzluluk
problemi olan tarım topraklarının ıslahının zor olması sebebiyle ve verimli bitkisel üretim gerçekleştirebilmek için tuza toleranslı bitkilerin tespit edilmesi veya geliştirilmesi gerekmektedir. Tuzluluk problemi olan topraklarda verimli bitkisel üretim yapabilmek için tuza toleranslı bitkilerin geliştirilmesi ve genetik ıslahın kullanılması ideal bir alternatif sağlayacaktır.
2.4. NaCl Tolerans Mekanizması
B içeriği yüksek olan topraklarda, borat formunda çözünebilir tuzların bulunması sebebiyle tuzluluğun bulunması kaçınılmaz bir gerçektir. Ayrıca yüksek tuz ve B içeren yer altı sularıyla yapılan sulama ile bitkiler aynı anda iki farklı strese maruz kalmaktadırlar (Nable ve ark., 1997). Örneğin, G. Avustralya (Marcar ve ark., 1999), İsrail ve Ürdün‘de Ürdün Nehri Vadisi topraklarının (Yermiyahu ve ark., 2008) yüksek tuz ve B birikimi gösterdiği; Kaliforniya‘da San Joaquin Vadisi‘nde (Grieve ve ark., 2010) ve Şili‘de Lluta Vadisi‘nde (Bastías ve ark., 2004) yüksek oranda tuz ve B içeren sulama sularıyla sulamanın yapıldığı bilinmektedir.
Tuz stresi ve B toksisitesi arasındaki karşılıklı (sinerjistik veya antogonistik) ilişkilerle ilgili araştırmacılar arasında henüz tam olarak bir fikir birliğine varılabilmiş değildir (Grieve ve ark., 2010). Tuz stresi ve B toksisitesi arasındaki antogonistik etki, yüksek B dozlarında NaCl‘nin düşük toksik etki göstermesi veya yüksek NaCl dozlarında B‘un düşük toksik etki göstermesi şeklinde veya her iki durum nedeniyle de ortaya çıkabilmektedir. Bazı araştırmacılar, B‘un, yapraklardaki Cl-
iyonu birikimini azaltmak suretiyle tuz stresine karşı pozitif etki gösterdiğini bildirmiştir (Holloway ve Alston, 1992; Yermiyahu ve ark., 2008). Fakat bu iyileşmenin tam olarak mekanizması bilinmemektedir. Diğer yandan, sulama suyundaki NaCl konsantrasyonunun artışıyla yapraktaki B miktarında görülen düşüşün, transpirasyonda oluşan azalma nedeniyle ksilem tarafından yapraklara taşınarak biriktirilen B miktarındaki azalmadan kaynaklandığı ileri sürülmüştür (Yermiyahu ve ark., 2008).
Tuz stresi ve B toksisitesi arasındaki ilişkiyle ilgili olarak yapılan bir diğer açıklama akuaporin fonksiyonuyla ilgilidir (Ballesta ve ark., 2008; Martínez-Ballesta ve ark., 2009). Dış ortamda aşırı B varlığında, plazma membranında yer alan akuaporinler aracılığıyla önemli oranda B taşınımının gerçekleştiği gösterilmiştir
(Dordas ve Brown, 2000). Tuz stresi altında bazı özel membran bileşenlerinin aktivitesinin, tuz tolerans mekanizmasında rol oynayan bazı akuaporin izoformlarının işlevselliğini düzenleyen B tarafından etkilenebileceği rapor edilmiştir (Martínez-Ballesta ve ark., 2009). Ayrıca tuza toleranslı bitkilerin tuzun atılımını sağlama yeteneklerinin içsel B konsantrasyonlarının azalmasına da katkı sağlayabileceği düşünülerek toksik B düzeylerine karşı daha toleranslı olabilecekleri ileri sürülmüştür (Alpaslan ve Gunes, 2001).
Bitkiler, tuz stresine maruz kaldıkları zaman ilgili biyokimyasal ve moleküler mekanizmaları devreye girdiği bildirilmiştir (Parida ve Das, 2005). Bu mekanizmaları üç farklı kategoride sınıflandırmak mümkündür (Roy ve ark., 2014). Bunlar:
Osmotik Tolerans Dokusal Tolerans İyon Atımı
Bitkiler yüksek tuz konsantrasyonu bulunan ortamlarda hücre içi Na+
/K+ oranını uygun seviyede tutabildikleri sürece büyüme ve gelişim süreçlerine devam edebilmektedirler. Bitkiler bu dengeyi koruyabilmek için kök, gövde ve yapraklarında ortak çalışan veya bu dokulardan birine özgü olarak çalışan birbirinden farklı üç sistem kullanmaktadır. Bu sistemler aşağıdaki gibi açıklanabilir;
1. SOS1 (Salt Overly Sensitive 1): Tuza hassasiyeti yüksek taşıyıcılar
2. NHX1 (Tonaplast Na+/H+ exchangers): Tonoplast (Vakuol) Na+/H+ iki yönlü taşıyıcısı
3. HKT (High affinity potassium (K) Transporter): Yüksek afiniteli potasyum taşıyıcılar
Bunların her biri bir veya daha fazla gen ile ilişkilendirilerek bitkilerin tuza karşı göstermiş oldukları moleküler direncin temelini oluşturduğu bildirilmiştir.
Toprak çözeltisinde bulunan NaCl‘e direkt olarak maruz kalan kökten Na+‘un girişi pasif olarak gerçekleşmektedir. Bu Na+‘un bitki köküne girişi kolaylaştırılmış difüzyon ile veya iyon kanalı şeklinde bulunan taşıyıcı proteinler [HKT (yüksek afiniteli K+ taşıyıcıları), LCT (düşük afiniteli katyon taşıyıcıları) ve NSCC (seçici olmayan katyon taşıyıcıları)] aracılığıyla sağlandığı rapor edilmiştir (Apse ve Blumwald, 2007). Bu şekilde köke giriş yapmış olan Na+
apoplastik yol ile kaspari şeridine kadar ilerlemektedir (Botella ve ark., 2005). Köke giriş yapan Na+ iyonlarının tamamı sürgüne taşınmaz. Bu iyonların bir kısmı kökteki hücrelerden geçiş sırasında vakuolde tutulur ya da bir kısmı kökten dışarıya atılır. Bu dışarıya atma faaliyetlerine katkı sağlayan yolaklardan biri de SOS yolağıdır. A. thaliana‘da iyon dengesi ile tuz toleransında rol oynayan ve genetik olarak birbirine bağlı SOS1, SOS2, SOS3 olmak üzere üç lokus teşhis edilmiştir. SOS genlerinin kontrolündeki SOS sinyal yolu, Na+
ve K+ iyon dengesini düzenlemektedir. Bu yolakta bulunan 3 adet gen ve genlerin ürünleri aktif olarak çalışmakta ve Na+ iyonları hücre dışına atılarak hücre içi yüksek K+
/Na+ oranı korunmaktadır (Yokoi ve ark., 2002). SOS genlerinin moleküler yapısı belirlenmiş olup, SOS3 geni kalsiyum bağlayıcı bir proteini, SOS2 geni bir serin/treonin protein kinazı, SOS1 geni ise plazma membranı Na+/H+ antiporterini kodladığı bilinmektedir (Borsani ve ark., 2003). SOS3‘ün, SOS2 ile etkileşirek ve SOS2‘yi aktive ettiği bilinmektedir. SOS2 ve SOS3 ise SOS1‘i aktive ederek SOS1‘in ekspresyon düzeyini kontrol ettiği rapor edilmiştir (Sairam ve Tyagi, 2004). Sonuç olarak SOS1‘in aktive edilmesiyle tuzluluk şartlarında iyon dengesi ve tuz toleransı sağlanmış olmaktadır (Yokoi ve ark., 2002).
Stres şartlarında sitoplazmalarında yüksek tuz bulunduran bitkiler, metabolik fonksiyonlarını devam ettirebilmek için Na+‘un fazlasını vakuollerinde depoladığı bilinmektedir. Bitkiler sitozolde düşük düzeyde Na+oranı sağlayabilmek için sodyumun kompartımanlaşması ve Na+
/H+ antiporter‘ları aracılığıyla dışarı atılması yoluna giderler. Tonoplasta lokalize olmuş olan Na+
/H+antiporter‘ları (Na+/H+ değiş-tokuşunu sağlayan transporter‘lar; NHX) aşırı miktardaki Na+‘un kompartımanlaşması için Na+‘u tonoplast boyunca sitozolden vakuole doğru aktif bir şekilde taşırlar ve çeşitli bitkilerde NHX‘leri aşırı ifade eden bazı çalışmalarda NHX‘ların bitkilerin tuz toleransında çok önemli bir role sahip olduklarını göstermiştir (Apse ve ark., 1999; Zhang ve Blumwald, 2001). Na+‘un sitozolden dış ortama doğru atılımı (efflux) plazma membranında lokalize olmuş olan Na+
/H+ antiporter‘ları (Freudenreich ve ark., 1997) sayesinde gerçekleştirilir. A. thaliana‘da bulunan tuza aşırı duyarlı gen (Salt-Overly Sensitive; SOS) olan AtSOS1 geni, bakteri ve mayadaki NHA genleriyle homologdur ve plazma membranında bulunan bir NHA proteinini kodlar. Bu gen plazma membranı boyunca net Na+ akışlarının kontrolünde görev alır (Shi ve ark., 2000). AtSOS1 geninin aşırı ekspresyonu hücrelerde Na+ birikimini sınırlayarak tuz toleransını arttırır (Katiyar-Agarwal ve ark., 2006; Verslues ve ark., 2007).
Kökte, Na+‘un ksilemden köke tekrar yüklenmesi Na+ iyonuna karşı seçici olmayan uniportlar (HKT ve NAX) aracılığıyla gerçekleştirilir. Bu durum yapraklarda ise Na+‘un ksilemden parankima hücrelerine taşınımı Na+‘a karşı seçici olmayan uniportlarla NSCC kanallarıyla gerçekleştiği bilinmektedir (Apse ve Blumwald, 2007). Yaprakta biriken fazla Na+, SOS1 ile ksileme tekrar yüklenebilceği gibi floem döngüsüne katılarakta köke tekrar gönderilebilir veya NHX aracılığı ile vokuolde biriktirilerek de azaltılacağı ifade edilmiştir (Botella ve ark., 2005). Na+‘un floemden köke geri taşınmasıyla ilgili mekanizma tam olarak aydınlatılamamış olmasına rağmen, Berthomieu ve ark. (2003)‘nin Arabidopsis ile yaptıkları çalışmada, Na+‘un sürgünden floeme aktarılarak köke geri gönderilmesine, AtHKT1 geninin ürünü olan Na+ taşıyıcılarının aracılık ettiği belirtilmiştir. Ayrıca, NaCl kaynaklı tuzluluk, hücrelerde Na+, Cl-, Ca+2 ve K+ gibi iyonların stabil durumlarının bozulmasına sebep olmaktadır. Bu durumda, hücre dışında artan Na+iyonlarının hücreye alınımı, hücre zarında yer alan çeşitli taşıyıcılar aracılıyla gerçekleşir. Hidrate Na+
ile K+‘un yarıçaplarının yakınlığı, K+ taşıyıcılarının bu iyonlar arasında ayrım yapmasını zorlaştırır ve bu durumda Na+, K+ ile rekabete girer. Bu gibi durumlarda çeşitli K+ kanalları Na+‘un hücre zarından sitoplazmaya geçişini sağlarlar (Aharon ve ark., 2003; Çulha ve Çakırlar, 2011).
Marschner (1995)‘e göre, eğer bir bitkide tuzdan sakınım (exclusion) ve tuzu kabullenme (inclusion) mekanizmalarından birisi iyi gelişmiş ise, bu bitki genotipinin tuza toleransı yüksektir. Tuzdan sakınım mekanizmasına sahip olan bitkiler, tuzu bünyesinden uzak tutarak tuzun alınmasını sınırlama yoluyla hücre içindeki tuz konsantrasyonunu sabit tutabilir ve bu şekilde toksisite zararından korunabilir. Kabullenme mekanizmasına sahip bitkiler ise Na+
ve Cl- iyonlarına doku toleransı göstermektedirler. Bitki Na+
iyonunu fazlaca aldığı halde, zararlanma belirtisi göstermiyor veya çok az etkileniyorsa doku toleransından söz edilebilir. Bu tip bitkilerde tuzun hücreler içinde tutulduğu, metabolizmasına dahil edilmediği ve tuz bezleri gibi özelleşmiş hücrelerde biriktirildiği bilinmektedir. Bu iki tolerans mekanizması işleyiş bakımından kabul ediliyor olsa da, tuza karşı toleransın mekanizması henüz tam olarak açıklanabilmiş değildir (Babourina ve ark., 2000).
2.5. Puccinellia distans (Jacq.) Parl.
Poaceae (buğdaygil) familyasının Poaideae çim alt familyasından olan (Choo ve ark., 1994) Puccinellia distans (Jacq.) Parl. bitkisi ―European alkali grass‖, ―weeping alkali grass‖ ve ―çorak çim bitkisi‖ olarak bilinmektedir (Wikipedia; Keskin, 2010).
ġekil 2.5.1. Puccinellia distans (Jacq. Parl.)‘a ait sınıflandırma (www.plants.usda.gov‘dan alınmıştır.)
P. distans bitkisi Orta Doğu, Asya, Avrupa, Amerika ve İran‘da tuzlu topraklarda doğal olarak yayılış gösteren bir C3 bitkisi olarak bilinmektedir (Clayton ve Renvoize, 2013).
Yüksek konsantrasyonlarda ağır metal içeren topraklarda yaşayabilen ve bu ağır metalleri kökleri aracılığıyla alarak diğer dokularında biriktirebilen bitkiler ‗hiperakümülatör bitkiler‘ olarak adlandırılırlar. Hiperakümülatör olarak bilinen bu bitkiler gövdelerinde, özellikle de yapraklarında fitotoksik hasar oluşmaksızın olağanüstü düzeylerde ağır metal biriktirebilme özelliğine sahiptirler (Öztürk, 2016). Hiperakümülatör olmayan türlerle kıyaslandığında hiperakümülatör bitkilere ait kökler topraktan ağır metalleri çok yüksek bir oranda alır, gövdeye daha yüksek bir hızda iletir ve gövde ile yapraklarda yüksek miktarlarda depolarlar (Rascio ve Navari-Izzo, 2011) . Brassicaceae (Lahanagiller) familyasının üyeleri en iyi bilinen hiperakümülatör bitkiler olup, bu tür bitki gruplarının yaklaşık olarak %25‘ini oluşturduğu rapor edilmiştir. Örneğin Thlaspi caerulescence (Cd, Ni, Pb ve Zn), T. goesingense (Ni ve Zn), T. ochroleucum (Ni ve Zn) ve T. rotunolifolium (Ni, Pb ve Zn) bünyesinde birden fazla ağır metal biriktirdiği bildirilmiştir. Ayrıca, bu familyaya Arabidopsis thaliana‘nın da dahil olduğu bilinmektedir (Özbek, 2015).
Sınıflandırma Bilimsel ve Yaygın Kullanılan Ġsmi
Alem Plantae – Plants
Alt Alem Tracheobionta – Vascular plants Üst Şube Spermatophyta – Seed plants Şube Magnoliophyta – Flowering plants Sınıf Liliopsida – Monocotyledons Alt Sınıf Commelinidae
Takım Cyperales
Familya Poaceae – Grass family Cins Puccinellia Parl. – alkaligrass
Genetik araştırmalarda sıklıkla tercih edilen ve bir bağırsak bakterisi olan Escherichia coli; genetik ve mikrobiyoloji alanında kullanılan ve bir maya türü olan Saccharomyces cerevisiae; bitki araştırmalarında sıklıkla kullanılan, çok küçük bir genoma sahip olma ve genom haritası çıkarılmış ilk bitki olan A.thaliana; bir nematod türü olan Caaenorhabditis elegans; moleküler genetik, popülasyon genetiği gibi araştırmalarda kullanılan Drosophila melanogaster; fare araştırmalarında kullanılan Mus musculus ve zebra balığı olan Danio rerio en sık kullanılan model organizmalar olarak bilinmektedir.
Çalışmalarda kullanılacak olan model organizma seçiminde; söz konusu materyal genomu ve homolojisinin yüksek olması, jenerasyonlar arası sürenin kısa olması, kolay kültüre alınması, deney manipülasyonlarına uygun olması, genetik analizlere uygun olması vb. gibi faktörler dikkate alınmaktadır. Bu doğrultuda kullanılan söz konusu model organizma olarak karşımıza çıkan A. thaliana‘nın ucuz ortamlarda gelişmesi, özel iklimsel isteklerinin olmaması, az miktarda mineral isteği nedeniyle genetik çalışmalar için uygundur. A. thaliana bitkisinin çok sayıda tohum üretmesi, hücrelerindeki toplam DNA miktarının oransal azlığı, mutant bireylerinin varlığı ve bu organizmanın gen aktarım çalışmalarına uygunluğu gibi özellikleri nedeniyle moleküler çalışmalar için ideal bitki olarak karşımıza çıkmaktadır. Bu özelliklerinin yanı sıra genetik transformasyonun kolay olması, DNA polimorfizminin yüksek olması, gen izolasyonunda ve diğer genom çalışmalarında kullanılması açısından laboratuvarda çalışmak için uygun bir organizmadır. Ayrıca Arabidopsis‘in genom dizisi 2000 yılının sonlarında tamamlanarak dünya üzerinde genom sekansı tamamlanan ilk bitki olduğu bildirilmiştir (Graham ve May, 2011). Ayrıca, A. thaliana bitkisinin en küçük genom (120 Mbp) yapısına sahip olduğu bilinmektedir (Bennetzen ve Hake, 2009).
P. distans, tuz stresi ve yüksek konsantrasyonda B bulunan topraklara karşı uyum göstermiş ve Türkiye‘de bor madenleri civarında da yaşayabilen, buğdayın tolere edebileceği maksimum B dozlarından çok daha yüksek dozlara ve B içeren topraklarda doğal olarak yaşayabilen hiperakümülatör ve çok yıllık bir bitki olarak bilinmektedir. Buğdaygiller familyasında olmasından dolayı önemli tahıl grupları için model bir
organizma olarak değerlendirilmektedir (Gezgı n ve ark., 2004; Stiles ve ark., 2010; Hamurcu ve ark., 2016).
Ülkemizde Orta Anadolu bor madenlerindeki 277 mg kg-1 düzeylerinde yüksek B içeren topraklarda rahatlıkla yaşayabildiği bilinmektedir. Birçok bitkinin 5 mg kg-1‘ın üzerinde B içeren topraklarda yaşayamadığı (Nable ve ark., 1997), bazı bezelye varyetelerinin ise toprakta en fazla 100 mg kg-1 düzeylerindeki B miktarına tolerans gösterebildikleri (Bagheri ve ark., 1994) göz önüne alındığında P. distans’ın sergilediği tolerans son derece dikkat çekicidir.
P. distans bitkisinde NaCl ve B uygulamaları ile yapılan çalışmaların sınırlı sayıda olduğu bilinmektedir. Söz konusu uygulamalar ile ilgili ulaşılabilen çalışmalar tarih sırasına göre aşağıda belirtilmiştir:
Venables ve Wilkins (1978), tuzlu alanlarda doğal olarak yetişebilen Agropyron repens, Agrostis stolonifera, Festuca rubra, Hordeum secalinum, Lolium perene ve P. distans türleri üzerinde yürüttükleri çalışmada, maksimum kök büyüme oranlarının tuza bağlı olarak farklılık gösterdiğini, Lolium ve Hordeum‘un tuz içermeyen kontrol grubunda en uzun köke sahip olduklarını, Puccinellia‘nın 100 mM tuz konsantrasyonunda en uzun kök oluşturduğu halde diğer türlerin 50 mM NaCl tuzda en uzun kök oluşturduklarını belirlemişlerdir. Araştırmacılara göre, bitki gelişimi dikkate alındığında diğer türler en fazla 100 mM NaCl‘de gelişebilirken Puccinellia‘nın 200 mM‘da etkilenmeden gelişmeye devam ettiğini ifade etmişlerdir.
Ashraf ve ark. (1986), Agrostis stolonifera L., Agrostis capillaris L., Holcus lanatus L., Lolium perenne L., Dactylis glomerata L., Festuca rubra L. ve Puccinellia distans (L.) Parl. türlerini 0, 150 ve 250 mol m-3 NaCl içeren besin çözeltisi içinde 14 gün boyunca yetiştirmiş ve bitkileri ortama verdikleri tepki açısından seçerek sınıflandırmışlardır. Yaptıkları bu çalışma sonunda artan tuz konsantrasyonunun tüm türlerin ortalama kök uzunluğunda azalmaya sebep olduğunu, 250 mol m-3 NaCl içeren besin çözeltisinde P. distans (L.) Parl. dışında tüm türlerde kök büyümesinin engellendiğini belirlemişlerdir.
Parker ve ark. (1991) tarafından gerçekleştirilen bir çalışmada Astragalus, Leucaena, Medicago, Trifolium, Elymus, Elytrigia, Festuca, Leymus, Oryzopsis, Psathyrostachys, Puccinellia ve Sporobolus cinslerinden pek çok tür, çeşit ve hattı B ve tuza tolerans bakımından incelediklerinde hem tür içinde hem de türler arasında önemli varyasyon olduğunu belirlemişler, ümit veren 15 türü fide gelişimi döneminde tekrar test ettiklerinde bunlar içerisinde en ümitvar olanların A. bisulcatus, A. racemosus, E. pontica, P. distans ve S. airoides olduğunu, bunlarda tuzluluk seviyesi (EC50 değerinin) 20 dS m-1 olduğunda çimlenmenin %50 azaldığını ve fide gelişim döneminde uygulanan 4 mM B‘dan etkilenmediklerini ifade etmişlerdir.
Ashraf ve Yasmin (1997), Cenchrus pennisetiformis Hochst. & Steud., Leptochloa fusca (L.) Kunth., Panicum turgidum Forssk., Pennisetum divisum (Gmel.) Henr. ve Puccinellia distans (L.) Parl. çim türlerinin tuza dayanıklılıklarını incelemek amacıyla bir çalışma gerçekleştirmiştir. Bu çalışma için; dört farklı tuz kullanarak (NaHCO3, MgSO4.7H2O, CaCl2.2H2O ve NaCl) hagland besin solüsyonu ile karışım hazırlamış ve 6 haftalık fidelere 2.4 (kontrol), 8, 16 ve 24 dS m-1 şeklinde tuz uygulaması gerçekleştirmişlerdir. Çalışma sonucunda; L. fusca ve P. distans‘ın tüm tuz uygulamalarından etkilenmediği, C. pennisetiformis ve P. turgidum‘un ise tuz toleransına karşı hassas olduğunu belirtmişlerdir. Ayrıca, P. distans‘ın sürgünlerinde yüksek Cl-ve düşük Na+
ile K+içerdiğini rapor etmişlerdir.
Moravcova ve Frantik (2002), P. distans ve P. limosa türlerinin çimlenmesi üzerine kuru depolama, sıcaklık ve NaCl tuzluluğuna cevabını incelemişlerdir. İstatistiksel analiz için, toplam çimlenme yüzdesi ile çimlenme hızı indeksini kullanmışlardır. Kuru depolama koşullarında sıcaklığın 10-30°C olduğu durumda çimlenme üzerine etkisinin en yüksek olduğu gözlenmiştir. Ancak, P. distans‘ın taze hasat tohumlarının 15°C sabit sıcaklıkta daha iyi çimlendiği rapor edilmiştir. Ayrıca, P. limosa‘nın tuzluluğa toleransının P. distans‘tan daha yüksek olduğu gözlenmiştir.
Alshammary ve ark. (2004)‘nın sera (saksı) ve su kültürü (hidroponik) ortamda gerçekleştirdikleri çalışmalarında Poa pratensis L., Festuca arundinacea Screb., Puccinellia distans (L.) Parl. ve Distichlis spicata (Torr.) bitkilerinin tuza karşı toleranslarını ve büyüme tepkilerini karşılaştırmalı olarak araştırmışlardır. Saksı denemelerinde yetiştirilen bitkilere farklı seviyelerde tuz içeren sulama suyu