www.biodicon.com Biological Diversity and Conservation
ISSN 1308-8084 Online; ISSN 1308-5301 Print
4/3 (2011) 58-70
Research article/Araştırma makalesi
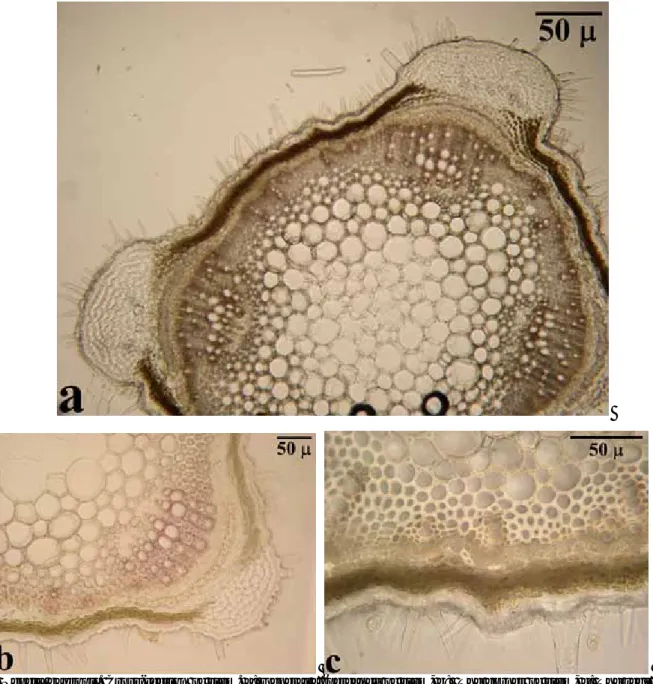
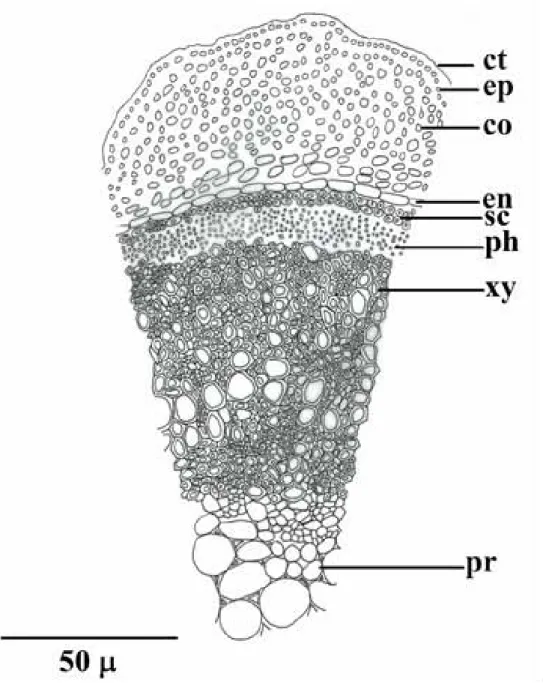
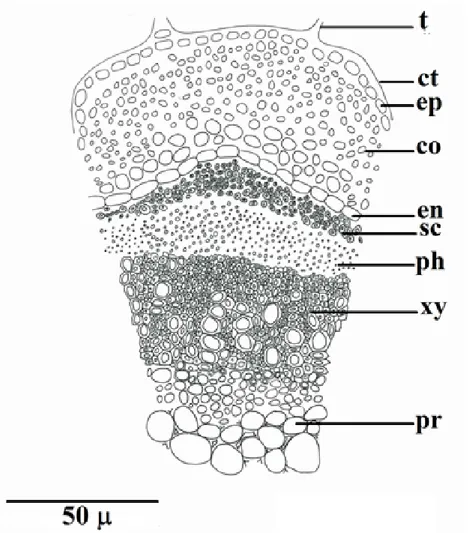
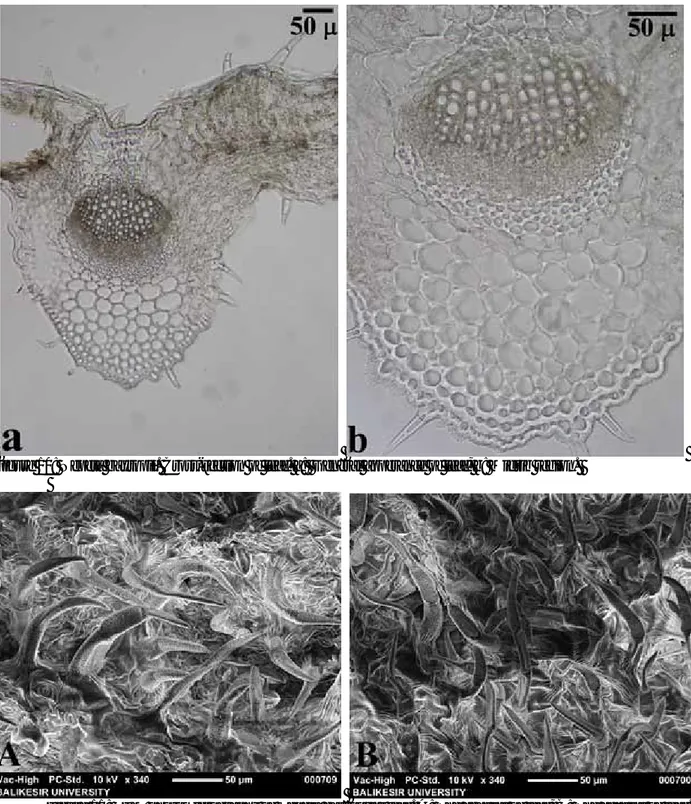
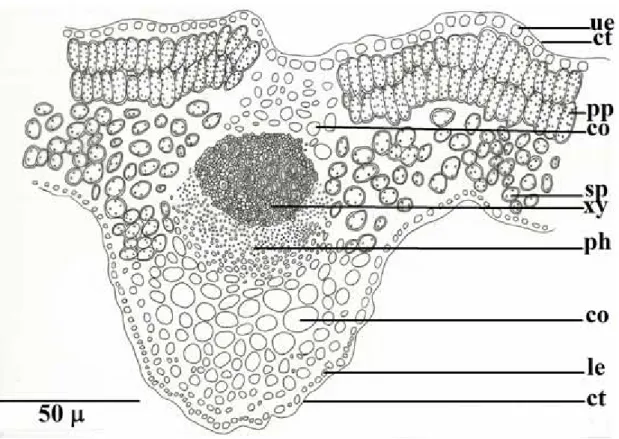
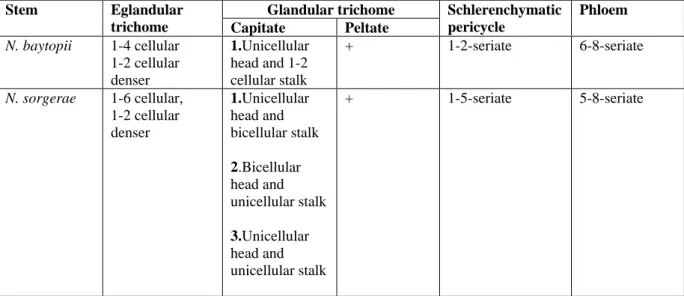
A comparative anatomical study on two endemic Nepeta L. species (N. baytopii and N. sorgerae)
Mikail ACAR
1,
Taner OZCAN
*2, Fatih SATIL
1, Tuncay DIRMENCI
21
Balıkesir University, Arts and Science Faculty, Department of Biology, 10145 Balıkesir, Turkey
2