Full Terms & Conditions of access and use can be found at
https://www.tandfonline.com/action/journalInformation?journalCode=thsb20
The Journal of Horticultural Science and Biotechnology
ISSN: 1462-0316 (Print) 2380-4084 (Online) Journal homepage: https://www.tandfonline.com/loi/thsb20
S
_allele identification and genetic diversity
analysis of apricot cultivars
Zehra Tuğba Murathan, Salih Kafkas, Bayram Murat Asma & Hayat Topçu
To cite this article: Zehra Tuğba Murathan, Salih Kafkas, Bayram Murat Asma & Hayat Topçu (2017) S_allele identification and genetic diversity analysis of apricot cultivars, The Journal of Horticultural Science and Biotechnology, 92:3, 251-260, DOI: 10.1080/14620316.2016.1255568 To link to this article: https://doi.org/10.1080/14620316.2016.1255568
Published online: 13 Jan 2017.
Submit your article to this journal
Article views: 208
View related articles
View Crossmark data
ARTICLE
S_allele identification and genetic diversity analysis of apricot cultivars
Zehra Tuğba Murathana, Salih Kafkasb, Bayram Murat Asmacand Hayat Topçub
aFaculty of Engineering, Food Engineering Department, Ardahan University, Ardahan, Turkey;bFaculty of Agriculture, Department of Horticulture, Çukurova University, Adana, Turkey;cFaculty of Agriculture, Department of Horticulture,İnönü University, Malatya, Turkey
ABSTRACT
In this study, the sexual incompatibility andS-allele diversity of 24 Turkish apricot cultivars, Paviot and Sakıt-1 as parents and 127 F1 progenies were identified by polymerase chain
reaction (PCR) and sequencing techniques. Additionally, genetic diversity and relatedness among the 24 cultivars were determined using 18 simple sequence repeat (SSR) markers from the genusPrunus. PCR for S_alleles identified nine different S-RNase alleles in the 24 apricot cultivars, namelySc,S1,S2,S8,S9,S20,S24,S52, andS53. All primers amplified only oneS_allele in
the cultivars Adilcevaz-1, Adilcevaz-3, Ethembey, Pasamismisi, Canakkale, and Soganci. Most of the Turkish cultivars were self-incompatible. TheScallele was present in only three cultivars
(Canakkale, Ethembey, Imrahor) that are, therefore, self-compatible. TheS_alleles of cultivars Paviot and Sakıt-1 displayed homology with the Sc,S2andS20, andS52alleles. In the 127 F1
genotypes, the twoS_alleles of Paviot were inherited by roughly half of the offspring, while about 76% of the offspring inherited theS52allele from Sakıt-1, and less than 24% inherited
S20. The amplification using all SSR 18 primers was successful and produced 128 polymorphic
alleles with an average of 7.11 alleles per locus. Among the apricot cultivars studied, expected heterozygosity (He) ranged from 0.33 to 0.72, observed heterozygosity (Ho) ranged from 0.42 to 1.00, PIC values were between 0.28 and 0.89, and similarity rates were between 0.30 and 0.68. The cultivars Levent and Ozal were genetically closest (0.68) while cultivars Sakıt-3 and Sogancı were the most distinct (0.30).
ARTICLE HISTORY Accepted 26 October 2016 KEYWORDS Genetic diversity; PCR; Prunus armeniaca L.; Self-incompatibility; SSR Introduction
Apricot (Prunus armeniaca L., 2n:16) belongs to the Rosaceae family and is one of the most important stone fruit crops in the world (Rehder, 1967). The global apricot production was about 4 million tons in 2013; and Turkey, Spain, Italy, France, and Greece accounted for more than 75% of the total production (Faostat, 2013). Turkey produces 812,000 tons of fresh and 162,000 tons of dry apricots annually and is the global leader in dry apricot production and export. Eastern Anatolia, especially Malatya, is the most important apricot growing region in Turkey (Asma & Ozturk,2005).
Apricots are classified into four major eco-geogra-phical groups, namely Central Asian, Irano-Caucasian, European, and Dzungar-Zailij (Kostina,
1969). While the European group is the youngest with least phenotypic variation, the Central Asian group shows highest phenotypic variability. The Irano-Caucasian group comprises apricots from Armenia, Georgia, Azerbaijan, Dagestan, Iran, Iraq, Syria, and Turkey (Mehlenbacher, Cociu, & Hough,
1990), and most of these cultivars are self-incompa-tible. Turkish apricots are also predominantly
gametophytically self-incompatible (Kostina, 1970) and this gametophytic self-incompatibility is con-trolled by a highly polymorphic but simple S-locus (De Nettancourt, 1977), whose gene product is an S-RNase enzyme in the styles or an F-box protein in the pollen (McClure et al.,1989). While self-compat-ibility alleles would allow the pollen tubes to grow in the style and reach the ovules, the self-incompatibility alleles stop this pollen tube growth if the same allele is present both in the pollen grain and in the pistil (Burgos, Ledbetter, Perez-Tornero, Ortin-Parraga, & Egea,1997). Commercial apricot cultivars in Turkey are predominantly self-incompatible and, therefore, require a pollinator for an economic fruit set. Furthermore, the breeding programs for generating new apricot cultivars using superior genotypes from the orchards have been in place for several decades and resulted in the development of a few cultivars with good quality fruits; however, they are all still self-incompatible (Asma, 2012; Bolat & Guleryuz,
1995; Gulcan et al., 2006; Misirli, Saglam, Gulcan, Olmez, & Sahin, 2006; Yilmaz, 2008). Therefore, identification of their S_alleles is necessary before they can be widely used for commercial cultivation.
CONTACTSalih Kafkas [email protected]
THE JOURNAL OF HORTICULTURAL SCIENCE AND BIOTECHNOLOGY, 2017 VOL. 92, NO. 3, 251–260
http://dx.doi.org/10.1080/14620316.2016.1255568
Traditionally, self-incompatibility is determined by monitoring the fruit set rate of pollinated flowers; but as this method is influenced by environmental con-ditions, fruit set varies annually and is both labor-and time-intensive (Milatovic & Nikolic, 2007). Recently, molecular methods, especially PCR, have been used to determine self-incompatibility in apri-cots (Badanes et al., 2000; Halasz et al., 2013; Jie, Shupeng, Jixiang, Manru, & Huairui, 2005; Romero et al.,2004). Even though there are several studies on S-allele identification in the Turkish apricot germ-plasm (Halasz, Pedryc, Ercisli, Yılmaz, & Hegedus,
2010; Paydas, Eti, Gülcan, Derin, & Yilmaz, 2006; Yilmaz, Kafkas, & Kargı Paydas,2013), their reassess-ment is necessary to clarify certain disparities. Molecular markers are valuable for germplasm char-acterization, genetic mapping, parental identification, and fingerprinting in plants, and have been exten-sively used for studying stone fruits (Krichen, Bourguiba, Audergon, & Trifi-Farah,2010; MirAli & Nabulsi,2003; Sa´nchez-Pe´rez, Ruiz, Dicenta, Egea, & Martı´nez-Go´mez, 2005). Of these, the simple sequence repeat (SSR) markers or microsatellites are advantageous compared to other marker systems as they are PCR-based, simple, independent of environ-mental effects, highly polymorphic, co-dominant, abundant, and highly reproducible (Gupta, Balyan, Sharma, & Ramesh, 1996; Li, Wang, Sun, Zhang, & Fang, 2012; Tautz, 1989). Additionally, the genetic characterization studies in apricots performed in the last two decades have predominantly used microsa-tellite markers (Bourguiba et al., 2012,2013; Pedryc et al., 2009; Rajia, Jannatizadehb, Fattahia, & Esfahlanica,2014; Sa´nchez-Pe´rez et al.,2005).
Therefore, in the present study, we reassessed S_alleles in 24 Turkish apricot cultivars and in the F1 progenies of a Paviot×Sakıt-1 cross using PCR
analysis and sequencing. We also mapped the genetic diversity among these 24 apricot cultivars using SSR markers to assess their potential use in cultivar breed-ing programs.
Material and methods
Plant material
Leaf samples from the 24 apricot cultivars, and 127 F1
progenies from the Paviot-Sakıt-1 cross and their parents were collected from the Inonu University, Apricot Research and Application Central Directorate. The F1 progeny of the Paviot-Sakıt-1
cross was raised in 2008. The cultivar Paviot was chosen for its large fruit size, orange pulp color, resistance to Sharka disease, and self-compatibility, whereas Sakıt-1 is a local cultivar with high fruit set, high Brix content, and sweet seeds.
DNA extraction, S_Allele PCR analysis, and sequencing
DNA isolation from leaf samples was performed using the CTAB (cetyl trimethyl ammonium bro-mide) protocol according to Doyle and Doyle (1987) but with minor modifications (Kafkas & Perl-Treves,2001). The concentration of the isolated DNA was determined by comparison with λ-DNA quantified using the agarose gel electrophoresis sys-tem (Thermo electron corporation EC135-90, Madison, WI, USA). DNA was diluted to a final concentration of 10 ng µL–1 for SSR and S_allele PCR.
PCR for S_alleles used primer pairs (Table 1) developed by Romero et al. (2004), Vilanova, Romero, Llacer, and Badenes (2005), Halasz, Pedryc, and Hegedus (2007), and Tao et al. (1999). PCR reactions were performed in a final volume of 25 µL and the reaction mixture contained 75 mM Tris-HCl (pH = 8.8), 20 mM (NH4)2SO4, 2 mM MgCl2, 0.1%
Tween 20, 100 µM each of dATP, dTTP, dGTP, dCTP, 0.2 µM forward and reverse primers, 1.0 unit of Taq DNA polymerase, and 20 ng of DNA. PCR amplification conditions were as follows: samples were pre-denatured at 94°C for 3 min, followed by 35 cycles of denaturation for 45 s at 94°C, annealing for 45 s at the amplification temperature of each primer, and extension for 60 s at 72°C. Samples were maintained at 72°C for 10 min for final exten-sion. PCR products were separated on the basis of band sizes by electrophoresis on a 2–3% agarose gel with 0.5X TBE, stained with ethidium bromide, and visualized under UV light.
Bidirectional Sanger sequencing of the amplified PCR fragments for the 24 apricot cultivars and the F1
progeny was performed at Medsantek Company (İstanbul, Turkey). The obtained sequences were aligned using the BLASTn (Basic Local Alignment Search Tool) program and the National Center for Biotechnology Information (NCBI) database (Altschul et al.,1997).
SSR amplification and data analysis
Eighteen primer pairs from different Prunus species (Aranzana, Garcia-Mas, Carbó, & Arús, 2002;
Table 1.The primers used to determineS_alleles of apricot cultivars.
Primers Sequence (5ʹ–3ʹ) Tm (°C) SRc-F CTC GCT TTC CTT GTT CTT GC 54 SRc-R GGC CAT TGT TGC ACA AAT TG 54 Pru C2-F CTTTGGCCAAGTAATTATTCAAAC 58 PruC4-R GGATGTGGTACGATTGAAGCG 58 PaConsI-F (C/A)CTTGTTCTTG(C/G)TTT(T/C)GCTTTCTTC 58 PaConsI-R CATG(A/G)ATGGTGAA(A/G)T(T/A)TTGTAATGG 58 AprSFB-F AAGAAWGARAYYTTRRTCGACAT 50 AprSFB-R ATYGAGWAAAACCAWRCTYTC 50
Cipriani, Lot, Huang, Marrazzo, & Peterlunger,1999; Dirlewanger, Cosson, Tavaud, Aranzana, & Poizat,
2002; Messina, Lain, Marrazzo, Cipriani, & Testolin,
2004; Mnejja, Garcia-Mas, Howad, & Arús, 2005; Sosinski, Gannavarapu, Hager, Beck, & King, 2000; Testolin et al.,2004) were used for SSR amplification, and the PCR reactions utilized a three-primer strat-egy reported by Scheulke (2000), but with minor modifications (Topcu, Coban, & Kafkas, 2016; Zaloglu, Kafkas, Dogan, & Guney, 2015). PCR reac-tions were performed in a total volume of 12.5 µL containing 10 ng DNA, 75 mM Tris-HCl (pH 8.8), 20 mM (NH4)2SO4, 2.0 mM MgCl2, 0.01% Tween 20,
200 µM of each dNTP, 10 nM of M13-tailed forward primer at the 5ʹ end, 200 µM reverse primer, 200 nM
universal M13 tail primer
(5-TGTAAAACGACGGCCAGT-3) labeled with FAM, VIC, NED or PET dye, and 0.8 U of Taq DNA polymerase.
PCR amplification was carried out in two conse-cutive steps. The first step consisted of an initial denaturation at 94°C for 2 min, followed by 30 cycles of 94°C for 30 s, 56–60°C for 45 s, and 72°C for 45 s. The second step involved eight cycles of 94°C for 30 s, 52°C for 45 s, and 72°C for 45 s, with a final extension at 72°C for 5 min. The PCR reaction pro-ducts were subjected to denaturation for subsequent capillary electrophoresis in an ABI 3130xl genetic analyzer (Applied Biosystems Inc., Foster City, CA, USA) using a 36-cm capillary array and POP7 matrix. Fragments were resolved using the ABI data collec-tion software (v. 3.0) and SSR fragment analysis was performed using the GeneScan Analysis Software (v. 4.0).
After fragment analysis of the SSR loci, the num-ber of alleles per locus (Na), effective numnum-ber of alleles (Ne), polymorphism information contents (PIC), expected heterozygosity (He) and observed heterozygosity (Ho) were calculated using the GenAlEx software (v. 6.5; Peakall & Smouse, 2006) and the PowerMarker software (v. 3.25; Liu & Muse,
2005). Genetic similarities between apricot cultivars were calculated using Jaccard’s coefficient (Jaccard,
1908), and the unweighted pair group method with arithmetic average (UPGMA) was used to construct a dendrogram using the NTSYSpc 2.11 V program (Exeter Software, Setauket, NY) (Rohlf, 2004).
Results
S_Allele PCR analysis
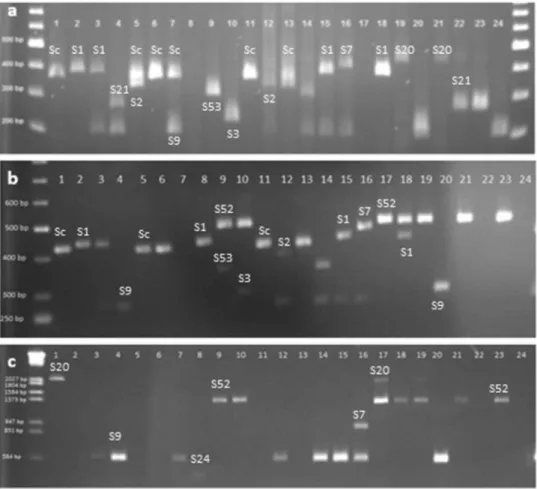
Initially, SRc-F/SRc-R primer pairs for the first intron were used in S-allele identification in the 24 apricot cultivars, and PCR detected the presence of an approximately 353-bp allele in the Adilcevaz-1, Alyanak, Cologlu, Canakkale, Ethembey, and
Imrahor cultivars (Figure1). After that, we used the SFB primer set to analyze further the 353-bp sequence amplified by the SRc-F/SRc-R primer pairs to identify whether the alleles were Sc, S8, or both.
Results indicated that the Canakkale cultivar had an Sc allele, and the Adilcevaz-1, Alyanak, and Cologlu
cultivars had the S8 allele, while the Imrahor and
Ethembey cultivars had both types of S_alleles. As we were not able to definitely identify all S_alleles in the 24 cultivars using the SRc-F/SRc-R primer pair, the PaConsI-F/PaConsI-R primer pair was used to amplify the first intron region of the apricot S-RNase and the PruC2-F/PruC4-R primer set was used to amplify the second intron. Initially, we analyzed the S-alleles of the 24 culti-vars by measuring the size of the bands; subse-quently, the bands amplified by all three primer pairs were sequenced for confirmed identification. Finally, the type of S_alleles in all 24 Turkish apricot cultivars was determined (Table 2). Cultivars Adilcevaz-3, Pasamismisi, Canakkale, Sakıt-3, and Soganci were found to have only one allele even when all primer pairs were used for amplification.
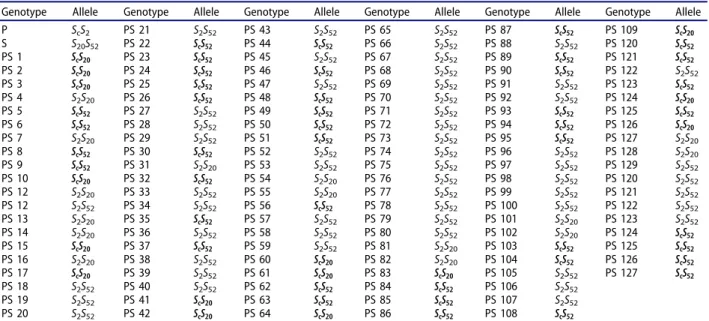
The S genotypes of the parental cultivars Paviot and Sakıt-1, and their 127 F1 progenies were also
determined and are listed in Table 3. A BLAST analysis of the S_allele sequences of the Paviot and Sakıt-1 cultivars showed 100% homology with the Sc/S2 and the S20/S52 alleles, respectively; and the
Paviot cultivar had a self-compatible allele. Of the 127 F1 progenies tested, 57 had the Sc allele, and
thus were classified as self-compatible. The other 70 F1 progenies were self-incompatible as they had
either an S2S52 or an S2S20 allele combination
(Figure 2). Each of Sc and S2 from Paviot was inherited by roughly half of the offspring; how-ever, very skewed inheritance was obtained in the S_alleles of Sakıt-1 as more than 76% of the off-spring inherited S52, and less than 24% inher-ited S20.
SSR fingerprinting
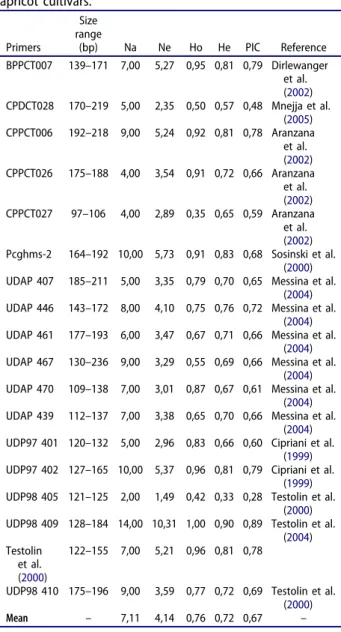
The SSR fingerprinting results for the 24 apricot cultivars are given in Table 4. The size of the SSR varied between 97 and 236 bp. A total of 128 alleles were obtained from 18 SSR loci, and all alleles were polymorphic among 24 apricot culti-vars. The average number of alleles per locus was 7.11. The locus UDP98-409 had the highest num-ber of alleles (Na = 14), while the UDP98-405 locus had the least (Na = 2). The PIC values ranged between 0.28 (UDP98-405) and 0.89 (UDP98-409) with an average value of 0.66.
Expected heterozygosity (He) values among the SSR loci ranged between 0.33 (UDP98-405) and
0.90 (UDP98-409) with an average value of 0.72, while the observed heterozygosity (Ho) values ran-ged from 0.42 (UDP98-405) to 1.00 (UDP98-409)
with an average value of 0.76. UPGMA analysis of the 24 apricot cultivars using the 128 polymorphic SSR alleles grouped the cultivars into two main clusters (Figure 3). All cultivars originating from the Malatya region were clustered together. The cultivars Levent with Ozal (0.68), and Adilcevaz-5 with Kabaasi (0.65) were the closest pairs of culti-vars in this cluster, while Sakıt-3 and Soganci were the most distant (0.30) cultivars.
Discussion
S genotype identification
In this study, the primer pairs for sequences in the first (SRc-F/SRc-R, SFBF/SFBR, PaConsI-F/PaConsI-R) and second (PruC2-F/PruC4-PaConsI-F/PaConsI-R) introns were used for S_allele identification in the 24 apricot cultivars, and a 353-bp allele was identified in the Adilcevaz-1, Alyanak, Cologlu, Canakkale, Ethembey, and Imrahor cultivars. While Vilanova et al. (2005) pro-posed this 353-bp allele to be the self-compatibility allele (Sc); however, Halasz et al. (2007) reported that
the SRc primer set could not distinguish between the Sc and S8 alleles in apricots due to their identical
RNase sequences. These authors also suggested
Figure 1.S_allele PCR analysis for the 24 apricot cultivars with the primers Src-F and Src-R (a), PaConsI-F and PaConsI-R (b) and PruC2-F and PruC4-R (c). Samples are 1: Adilcevaz-1, 2: Adilcevaz-3, 3: Adilcevaz-5, 4: Akcadag Gunay, 5: Alyanak, 6: Canakkale, 7: Cologlu, 8: Sam, 9: Gec aprikoz, 10: Guz aprikozu, 11: Ethembey, 12: Hasanbey, 13: Imrahor, 14: Ismailaga, 15: Kabaasi, 16: Kurukabuk, 17: Levent, 18: Mahmudun erigi, 19: Ozal, 20: Pasamismisi, 21: Sakıt-1, 22: Sakıt-3, 23: Sekerpare, 24: Soganci. Table 2.S genotypes in apricot cultivars.
Cultivar Origin Paydas et al. (2006) Gulcan et al. (2006) Yilmaz et al. (2013) Halasz et al. (2010) This study Adilcevaz-1 Bitlis – – SC S8S19 S8S20 Adilcevaz-3 Bitlis – – SI S13S19 S1 Adilcevaz-5 Bitlis – – SI S9S13 S1S9 Akçadag Gunay Malatya SI SI SI S3S9 S9S21 Alyanak Izmir SI SI SC S2S8 S2S8 Canakkale Canakkale SC SC SC ScSc ScSc Cologlu Malatya SI SI SC S8S9 S8S9 Sam Izmir – SC SI ScS2 S1S24
Gec Aprikoz Igdir SI SI SI S6S11 S52S53
Guz Aprikozu Igdir SI SI SI S6 S3S52
Ethembey Edirne SC SC SC ScS8 ScS8 Hasanbey Malatya SI SI SI S2S9 S2S9 Imrahor Izmir SC SC SC S3 ScS8 Ismailaga Malatya SI SI SI S9S11 S9S53 Kabaasi Malatya SC SI SI S9S13 S1S9 Kurukabuk Malatya SI SI SI S7S9 S7S9 Levent Malatya – – SI S6S19 S20S52 Mahmudun erigi Malatya SI SI SI S13 S1S52 Ozal Malatya – – SI S6S9 S20S52 Pasamismisi Malatya SI SI SI ScS8 S9 Sakit-1 Hatay SC SC SI S6S19 S20S52 Sakit-3 Hatay – SI SI S13S19 S21 Sekerpare Igdir – SI SI S3S6 S21S52 Soganci Malatya – SI SI S6S9 S9
using the SFB primer set for successful distinction between these two alleles. Therefore, we used the SFB primer set during PCR reactions to distinguish between the S8and Scalleles and found that while the
cultivar Canakkale had the Sc allele and cultivars
Adilcevaz-1, Alyanak, and Cologlu had the S8 allele,
cultivars Imrahor and Ethembey had both the Scand
the S8alleles. The S-allele sequences obtained in this
study showed homology with reported allele sequences for Sc, S1, S2, S3, S7, S8, S9, S20, S21, S24,
S52, and S53 of Prunus armeniaca available in the
NCBI database (Gu, Wu, Du, Yang, & Zhang, 2013; Jiang et al., 2010; Raz, Stern, Bercovich, & Goldway, 2009; Romero et al., 2004; Vilanova et al., 2006; Zhang et al., 2008). We found that S9 was the most
commonly found allele as it was amplified from nine cultivars, and previous reports also state that the S9
allele is the most commonly amplified allele in Turkish apricots (Halasz et al., 2010). Allele S52 was
the next most common allele and was detected in seven of the 24 cultivars.
There are several reports in the literature that describe S_alleles in Turkish cultivars; however, there are disparities among the studies with respect to S_allele identification. A comparison of the S_alleles present in the 24 cultivars (reported here) with those reported in the literature is provided in
Table3. While Gulcan et al. (2006) and Paydas et al. (2006) ascertained self-compatibility of the cultivars by self and artificial pollination in the orchard, Yilmaz et al. (2013) used the SRc-F/SRc-R primer combination to check for the presence of the Scallele
in the cultivars. Comparably, even though Gulcan et al. (2006) and Halasz et al. (2010) found that the cultivar Sam was self-compatible, we and others (Yilmaz et al.,2013) report it to be self-incompatible. Similar results were also obtained with the Imrahor and Pasamismisi cultivars. Halasz et al. (2010) reported the presence of the S3allele in the Imrahor
cultivar, we and others (Gulcan et al., 2006; Paydas et al., 2006; Yilmaz et al., 2013) have detected the presence of the self-compatibility allele. The reported differences during field studies among these cultivars might be due to the fact that ribonuclease synthesis is blocked depending on environmental conditions, which result in the temporary removal of self-incom-patibility by activating the self-incomself-incom-patibility gene (Bassi, Bartolini, & Viti,2006).
However, this argument does not explain simi-lar differences observed in laboratory studies, and it is thought that these differences may have occurred due to the mistakes in labeling or hand-ling the leaf and DNA samples afterward. Additionally, self-incompatibility alleles of certain
Table 3.S_alleles of parents and F1genotypes in this study.
Genotype Allele Genotype Allele Genotype Allele Genotype Allele Genotype Allele Genotype Allele P ScS2 PS 21 S2S52 PS 43 S2S52 PS 65 S2S52 PS 87 ScS52 PS 109 ScS20 S S20S52 PS 22 ScS52 PS 44 ScS52 PS 66 S2S52 PS 88 S2S52 PS 120 ScS52 PS 1 ScS20 PS 23 ScS52 PS 45 S2S52 PS 67 S2S52 PS 89 ScS52 PS 121 ScS52 PS 2 ScS20 PS 24 ScS52 PS 46 ScS52 PS 68 S2S52 PS 90 ScS52 PS 122 S2S52 PS 3 ScS20 PS 25 ScS52 PS 47 S2S52 PS 69 S2S52 PS 91 S2S52 PS 123 ScS52 PS 4 S2S20 PS 26 ScS52 PS 48 ScS52 PS 70 S2S52 PS 92 S2S52 PS 124 ScS20 PS 5 ScS52 PS 27 S2S52 PS 49 ScS52 PS 71 S2S52 PS 93 ScS52 PS 125 ScS52 PS 6 ScS52 PS 28 S2S52 PS 50 ScS52 PS 72 S2S52 PS 94 ScS52 PS 126 ScS20 PS 7 S2S20 PS 29 S2S52 PS 51 ScS52 PS 73 S2S52 PS 95 ScS52 PS 127 S2S20 PS 8 ScS52 PS 30 ScS52 PS 52 S2S52 PS 74 S2S52 PS 96 S2S52 PS 128 S2S20 PS 9 ScS52 PS 31 S2S20 PS 53 S2S52 PS 75 S2S52 PS 97 S2S52 PS 129 S2S52 PS 10 ScS20 PS 32 ScS52 PS 54 S2S20 PS 76 S2S52 PS 98 S2S52 PS 120 S2S52 PS 12 S2S20 PS 33 S2S52 PS 55 S2S20 PS 77 S2S52 PS 99 S2S52 PS 121 S2S52 PS 12 S2S52 PS 34 S2S52 PS 56 ScS52 PS 78 S2S52 PS 100 S2S52 PS 122 S2S52 PS 13 S2S20 PS 35 ScS52 PS 57 S2S52 PS 79 S2S52 PS 101 S2S20 PS 123 S2S52 PS 14 S2S20 PS 36 S2S52 PS 58 S2S52 PS 80 S2S52 PS 102 S2S20 PS 124 ScS52 PS 15 ScS20 PS 37 ScS52 PS 59 S2S52 PS 81 S2S20 PS 103 ScS52 PS 125 ScS52 PS 16 S2S20 PS 38 S2S52 PS 60 ScS20 PS 82 S2S20 PS 104 ScS52 PS 126 ScS52 PS 17 ScS20 PS 39 S2S52 PS 61 ScS20 PS 83 ScS20 PS 105 S2S52 PS 127 ScS52 PS 18 S2S52 PS 40 S2S52 PS 62 ScS52 PS 84 ScS52 PS 106 S2S52 PS 19 S2S52 PS 41 ScS20 PS 63 ScS52 PS 85 ScS52 PS 107 S2S52 PS 20 S2S52 PS 42 ScS20 PS 64 ScS20 PS 86 ScS52 PS 108 ScS52
P = Paviot, S = Sakıt-1, PS = Paviot×Sakıt-1 F1genotypes.
Figure 2.S_allele PCR analysis for F1genotypes with the primers Src-F and Src-R (a), and PruC2-F and PruC4-R (b).
cultivars reported here are different from those reported by Halasz et al. (2010). Specifically, while cultivars Alyanak (S2S8), Canakkale (ScSc),
Cologlu (S8S9), Ethembey (ScS8), Hasanbey (S2S9),
and Kurukabuk (S7S9) appear to harbor identical
alleles, as reported here and by Halasz et al. (2010), the other 18 cultivars showed differences in at least one S_allele (Table 4). It is also possible that the low resolution of agarose gels in identify-ing S_alleles may have led to misidentification of
the S_alleles in some cultivars (Sambrook & Russell, 2001). Thus, sequencing the S_alleles obtained after separation on agarose gels would be a better method for a confirmed identification of S_allele in case of discrepancies.
S_allele sequencing revealed that the Paviot and Sakıt-1 cultivars have the Sc/S2 and S20/S52, allele
combinations, respectively. While the cultivar Paviot and 57 of the 127 F1progenies had the Sc allele, the
remaining F1 progeny and Sakıt-1 were self-Figure 3.UPGMA dendrogram of the 24 apricot cultivars based on 18 SSR markers.
incompatible. Asma (2012) reported that progeny with no. 42 and no. 96 have high yield and high fruit quality and are now being appropriately regis-tered; we report that progeny no. 42 has the Scallele
and that progeny no. 96 is self-incompatible.
SSR polymorphism
We analyzed SSR polymorphism to identify genetic diversity among 24 apricot cultivars; many other stu-dies have also used SSRs and other marker systems such as RAPD, RFLP, and ISSR (Gogorcena & Parfitt,
1994; Hurtado et al., 2001; Li, Zhao, & Miao, 2014; Yilmaz, Paydas Kargi, & Kafkas, 2012; Yuan et al.,
2007). Previous studies suggest that, compared to other marker systems, SSRs are better suited for char-acterizing genetic diversity among apricot cultivars because of their high degree of polymorphism, repeatability, and co-dominant nature (Gupta et al.,
1996; Sa´nchez-Pe´rez et al., 2005; Yilmaz et al.,
2012).
Eighteen SSR loci produced 128 polymorphic alleles with an average of 7.11 alleles per locus. Other studies reported the average number of alleles per locus to be 5.3 from 39 cultivars (Rajia et al.,
2014), 4.1 from 48 cultivars by fingerprinting (Hormaza, 2002), and 7.95 from 183 cultivars (Bourguiba et al., 2013). The values reported here are in confirmation with those previously reported probably because we have characterized 24 highly diverse apricot genotypes.
We observed that the UDP98-409 locus had the highest number of alleles per locus (Na = 14) while the UDP98-405 locus had the least (Na = 2). Similar results have been reported by other authors. Yilmaz et al. (2012) reported the presence of 84 polymorphic alleles using 16 SSR primers and UDP98-409, where the pchgms–2 locus had the highest number of alleles (Na = 7). Sa´nchez-Pe´rez et al. (2005) also amplified 66 polymorphic alleles using 17 peach SSR primers, and the number of alleles per locus was 3.9.
A high PIC value depends on the number and frequency distribution of the alleles measured, mar-kers with PIC values exceeding 0.500 being consid-ered more informative (Botstein, White, Skolnick, & Davis, 1980). The PIC value ranged from 0.28 (UDP98-405) to 0.89 (UDP98-409) with an average value of 0.66 in our study, and it was found to be 0.56 by Rajia et al. (2014), 0.59 by Bourguiba et al. (2012), and 0.73 by Yilmaz et al. (2012).
In studies, the most important criterion is the high level of heterozygosity. Expected heterozygosity (He) ranged from 0.33 to 0.90 with an average value of 0.72 and observed heterozygosity (Ho) ranged from 0.42 to 1.00 with an average value of 0.76. Our results are higher than the mean values reported by previous studies on apricots. The expected heterozygosity value was reported 0.59 by Bourguiba et al. (2013) and 0.75 by Pedryc et al. (2009), while the observed heterozygosity values were reported as 0.64 by Zhebentyayeva, Reighard, Gorina, and Abbott (2003) and 0.56 by Bourguiba et al. (2013).
The apricot genotypes analyzed in this study were grouped into five main clusters with a similarity level of 0.24. The first major group (Cluster 1) was further divided into two subgroups with three accessions, including two accessions from Bitlis. Cluster 2 con-tained two subgroups and four accessions, and included two Iğdır cultivars. The largest cluster (Cluster 3) contained most of the economically important apricot cultivars grown in Malatya/ Turkey, and was distinct from the other two sub-groups, and comprised 10 Malatya cultivars and three other (Bitlis, Iğdır, Hatay) cultivars (Figure 3). The late-ripening cultivars Levent and Ozal (0.68), and Adilcevaz-5 and Kabaasi (0.65) (which is the second most widely grown dried apricot cultivar in Malatya/Turkey) were the closest pairs of cultivars in
Table 4.Variability parameters of polymorphic SSR loci for 24 apricot cultivars. Primers Size range (bp) Na Ne Ho He PIC Reference BPPCT007 139–171 7,00 5,27 0,95 0,81 0,79 Dirlewanger et al. (2002) CPDCT028 170–219 5,00 2,35 0,50 0,57 0,48 Mnejja et al. (2005) CPPCT006 192–218 9,00 5,24 0,92 0,81 0,78 Aranzana et al. (2002) CPPCT026 175–188 4,00 3,54 0,91 0,72 0,66 Aranzana et al. (2002) CPPCT027 97–106 4,00 2,89 0,35 0,65 0,59 Aranzana et al. (2002) Pcghms-2 164–192 10,00 5,73 0,91 0,83 0,68 Sosinski et al. (2000) UDAP 407 185–211 5,00 3,35 0,79 0,70 0,65 Messina et al.
(2004) UDAP 446 143–172 8,00 4,10 0,75 0,76 0,72 Messina et al.
(2004) UDAP 461 177–193 6,00 3,47 0,67 0,71 0,66 Messina et al.
(2004) UDAP 467 130–236 9,00 3,29 0,55 0,69 0,66 Messina et al.
(2004) UDAP 470 109–138 7,00 3,01 0,87 0,67 0,61 Messina et al.
(2004) UDAP 439 112–137 7,00 3,38 0,65 0,70 0,66 Messina et al.
(2004) UDP97 401 120–132 5,00 2,96 0,83 0,66 0,60 Cipriani et al.
(1999) UDP97 402 127–165 10,00 5,37 0,96 0,81 0,79 Cipriani et al.
(1999) UDP98 405 121–125 2,00 1,49 0,42 0,33 0,28 Testolin et al.
(2000) UDP98 409 128–184 14,00 10,31 1,00 0,90 0,89 Testolin et al.
(2004) Testolin
et al. (2000)
122–155 7,00 5,21 0,96 0,81 0,78
UDP98 410 175–196 9,00 3,59 0,77 0,72 0,69 Testolin et al. (2000) Mean – 7,11 4,14 0,76 0,72 0,67 –
cluster 3. Gürcan et al. (2015) investigated the Turkish apricot germplasm using SSR markers and found that cultivars Adilcevaz 5 and Kabaasi were located close to each other in the tree with identical sequences at 16 of the 17 loci. Cluster 4 was divided into two subgroups and cultivars Sakıt-3 and Soganci (0.31) were the most distinct. Cluster 5 was further divided to form two subgroups with two accessions, namely Alyanak and Ethembey, which originated from İzmir and Edirne, respectively.
Conclusion
In the present study, S_alleles of 24 apricot culti-vars and 127 F1progenies from Paviot x Sakit-1 F1
population were analyzed. In addition, genetic diversity and relatedness among the 24 cultivars were also determined using SSR markers. The results indicated that only three (Canakkale, Ethembey, Imrahor) of the 24 cultivars have the Sc allele and are, therefore, self-compatible. Allele
S52 is the most common allele and was detected in
seven of the 24 cultivars. The observed discrepan-cies in the results reported herein and those in the literature were reassessed by sequencing the S_alleles. The alleles of Sakıt-1 had a skewed seg-regation, while the alleles of Paviot segregated as expected. A total of 57 F1 progenies had the Sc
allele and the allelic distribution in F1 genotypes
was detected to be 11.8% for S2S20, 43.3% for S2S52,
11.8% for ScS20, and 33.1% for ScS52. The F1
pro-geny no. 42 was self-compatible with high yield and good fruit quality and is a candidate for registra-tion as a new cultivar. The use of S_alleles for identifying superior genotypes has great potential to be beneficial for fruit-farming in the future and can also contribute economically, both to manufac-turers and farmers. The amplification of 18 SSR primer pairs in 24 cultivars produced 128 poly-morphic alleles and the ability to distinguish them. A high degree of polymorphism and genetic diversity was obtained from SSRs, which further substantiates that SSRs are one of the best marker systems for genetic diversity analysis.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was funded by The Scientific and Technological Research Council of Turkey (TUBITAK) [grant number: 113Z540].
References
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., & Lipman, D.L. (1997). Gapped BLAST and PSI-BALST: A new generation of protein database search programs. Nucleic Acids Research, 25, 3389–3402. doi:10.1093/nar/25.17.3389
Aranzana, M.J., Garcia-Mas, J., Carbó, J., & Arús, P. (2002). Development and variability analysis of microsatellite markers in peach. Plant Breeding, 121, 87–92. doi:10.1046/j.1439-0523.2002.00656.x
Asma, B.M. (2012). Determination of durability to Sharka Disease (Plum Pox Virus, PPV) of hybrid apricots, Investigations onbreeding of new fresh and dry apricots (II. Part) (Project report no: 2010/34). Turkey: Inonu University, Malatya.
Asma, B.M., &Ozturk, K. (2005). Analysis of morpholo-gical, pomological and yield characteristics of some apricot germplasm in Turkey. Genetic Research and Crop Evolution, 52, 305–313. doi: 10.1007/s10722-003-1384-5
Badanes, M.L., Hurdato, M.A., Sanz, F., Archelos, D.M., Burgos, L., Egea, J., & Llacer, G. (2000). Searching for molecular markers linked to male sterility and self-com-patibility in apricot. Plant Breeding, 119, 157–160. doi:10.1046/j.1439-0523.2000.00463.x
Bassi, D., Bartolini, S., & Viti, R. (2006). Recent advances on environmental and physiological challenges in apri-cot growing. Acta Horticulture, 717, 23–32. doi:10.17660/ActaHortic.2006.717.1
Bolat, I., & Guleryuz, M. (1995). Selection of late matura-tion wild apricot (Prunus armeniaca L.) forms on erzin-can plain. Acta Horticulture, 384, 183–188. doi:10.17660/ ActaHortic.1995.384.26
Botstein, D., White, R.L., Skolnick, M., & Davis, R.W. (1980). Construction of a genetic linkage map in man using restriction fragment length polymorphisms. The American Journal of Human Genetics, 32, 314–331. Bourguiba, H., Audergon, J.M., Krichen, L., Trifi-Farah, N.,
Mamouni, A., Trabelsi, S., & Khadari, B. (2012). Genetic diversity and differentiation of grafted and seed propa-gated apricot (Prunus armeniaca L.) in the Maghreb region. Scientia Horticulture, 142, 7–13. doi:10.1016/j. scienta.2012.04.024
Bourguiba, H., Khadari, B., Krichen, L., Trifi-Farah, N., Mamouni, A., Trabelsi, S., & Audergon, J.M. (2013). Genetic relationships between local North African apri-cot (Prunus armeniaca L.) germplasm and recently introduced varieties. Sciences Hort, 152, 61–69. doi:10.1016/j.scienta.2013.01.012
Burgos, L., Ledbetter, C.A., Perez-Tornero, O., Ortin-Parraga, F., & Egea, J. (1997). Inheritance of sexual incompatibility in apricot. Plant Breeding, 116, 383– 386. doi:10.1111/pbr.1997.116.issue-4
Cipriani, G., Lot, G., Huang, W.G., Marrazzo, M.T., & Peterlunger, E. (1999). AC/GT and AG/CT microsatel-lite repeats in peach [Prunus persica (L) Batsch]: Isolation, characterisation and cross species amplifica-tion in Prunus. Theoretical and Applied Genetics, 99, 65– 72. doi:10.1007/s001220051209
De Nettancourt, D. (1977). Incompatibility in Angiosperms. New York: Monographs on theoretical and applied genetics, Springer-Verlag, Berlin/Heidelberg.
Dirlewanger, E., Cosson, P., Tavaud, M., Aranzana, J., & Poizat, C. (2002). Development of microsatellite markers in peach [Prunus persica (L.) Batsch] and their use in genetic diversity analysis in peach and sweet cherry
(Prunus avium L.). Theoretical and Applied Genetics, 105, 127–138. doi:10.1007/s00122-002-0867-7
Doyle, J.J., & Doyle, J.L. (1987). A rapid isolation proce-dure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11–15.
Faostat. (2013). Retrieved February 4, 2015, from http:// apps.fao.org.
Gogorcena, Y., & Parfitt, D.E. (1994). Evaluation of RAPD marker consistency for detection of polymorphism in apricot. Scientia Horticulture, 59, 163–167. doi:10.1016/ 0304-4238(94)90083-3
Gu, C., Wu, J., Du, Y.H., Yang, Y.N., & Zhang, S.L. (2013). Two different Prunus SFB alleles have the same function in the self-incompatibility reaction. Plant Molecular Biology Reporter, 31, 425–434. doi:10.1007/s11105-012-0518-3
Gulcan, R., Misirli, A., Saglam, H., Yorgancioglu, U., Erkan, S., Gumus, M., . . . Demir, T. (2006). Properties of Turkish apricot land races. Acta Horticulture, 701, 191–198. doi:10.17660/ActaHortic.2006.701.28
Gupta, P.K., Balyan, H.S., Sharma, P.C., & Ramesh, B. (1996). Microsatellites in plants: A new class of molecu-lar markers. Current Science, 70, 45–54.
Gürcan, K., Öcal, N., Yılmaz, K.U., Ullah, S., Erdoğan, A., & Zengin, Y. (2015). Evaluation of Turkish apricot germplasm using SSR markers: Genetic diversity assess-ment and search for Plum pox virus resistance alleles. Scientia Horticulturae, 193, 155–164. doi:10.1016/j. scienta.2015.07.012
Halasz, J., Hegedus, A., Szikriszt, B., Ercişli, S., Orhan, E., & Unlu, H.M. (2013). The S-genotyping of wild-grown apricots reveals only self-incompatible accessions in the Erzincan region of Turkey. Turkish Journal of Biology, 37, 733–740. doi:10.3906/biy-1306-27
Halasz, J., Pedryc, A., Ercisli, S., Yılmaz, K.U., & Hegedus, A. (2010). S-genotyping supports the genetic relation-ships between Turkish and Hungarian apricot
germ-plasm. Journal of The American Society for
Horticultural Science, 135, 410–417.
Halasz, J., Pedryc, A., & Hegedus, A. (2007). Origin and dissemination of the pollen-part mutated SC-haplotype that confers self-compatibility in apricot (Prunus arme-niaca). New Phytology, 176, 793–803. doi: 10.1111/j.1469-8137.2007.02220.x
Hormaza, J.I. (2002). Molecular characterization and simi-larity relationships among apricot genotypes using sim-ple sequence repeats. Theoretical and Applied Genetics, 104, 321–328. doi:10.1007/s001220100684
Hurtado, M.A., Badenes, M.L., Llacer, G., Westman, A., Beck, E., & Abbott, G.A. (2001). Contribution to apricot genetic analysis with RFLP, RAPD and AFLP markers. Acta Horticulture, 546, 417–420. doi:10.17660/ ActaHortic.2001.546.54
Jaccard, P. (1908). Nouvelles recherches sur la distribution florale. Bulletin de la Societe Vaudoise des Sciences Naturelles, 44, 223–270.
Jiang, X., Wang, D.J., Feng, J.R., Zhang, D.H., Jiang, J.Q., & Liu, Y.X. (2010). Identification of self-incompatibility genotype of apricot (Unpublished). NCBI Gene Bank. Jie, Q., Shupeng, G., Jixiang, Z., Manru, G., & Huairui, S.
(2005). Identification of self-incompatibility genotypes of apricot (Prunus armeniaca L.) by S-allele-specific PCR analysis. Biotechnology Letters, 27, 1205–1209. doi:10.1007/s10529-005-0019-7
Kafkas, S., & Perl-Treves, R. (2001). Morphological and molecular phylogeny of Pistacia species in Turkey.
Theoretical and Applied Genetics, 102, 908–915. doi:10.1007/s001220000526
Kostina, K.F. (1969). The use of varietal resources of apri-cots for breeding. Trud Nikit Botanic Sada, 40, 45–63. Kostina, L.D. (1970). Self-fertility studies in apricot. Trud
Nikit Botanic Sada, 15, 7–17.
Krichen, L., Bourguiba, H., Audergon, J.M., & Trifi-Farah, N. (2010). Comparative analysis of genetic diversity in Tunisian apricot germplasm using AFLP and SSR mar-kers. Scientia Horticulture, 127, 54–63. doi:10.1016/j. scienta.2010.09.012
Li, H., Wang, K., Sun, X., Zhang, C., & Fang, J. (2012). Development of EST-derived SSR markers and their applications in apple genetic diversity analysis. Jiangsu Journal of Agricultural Science, 28, 158–165.
Li, M., Zhao, Z., & Miao, X. (2014). Genetic diversity and relationships of apricot cultivars in North China revealed by ISSR and SRAP markers. Scientia
Horticulture, 173, 20–28. doi:10.1016/j.
scienta.2014.04.030
Liu, K., & Muse, S.V. (2005). PowerMarker: Integrated analysis environment for genetic marker data. Bioinformatics, 21, 2128–2129. doi: 10.1093/bioinfor-matics/bti282
McClure, B.A., Ebert, P.R., Anderson, M.A., Simpson, R.J., Sakiyama, F., & Clarke, A.E. (1989). Style self-incompat-ibility gene products of Nicotiana alata are ribonu-cleases. Nature, 342, 955–957. doi:10.1038/342955a0
Mehlenbacher, S.A., Cociu, V., & Hough, L.F. (1990). Apricots, genetic resources of temperate fruit and nut crops. International Society of Horticultural Science, 3, 65–107.
Messina, R., Lain, O., Marrazzo, M.T., Cipriani, G., & Testolin, R. (2004). New set of microsatellite loci isolated in apricot. Molecular Ecology Notes, 4, 432–434. doi:10.1111/men.2004.4.issue-3
Milatovic, D., & Nikolic, D. (2007). Analysis of self-(in) compatibility in apricot cultivars using fluorescence microscopy. The Journal of Horticultural Science and
Biotechnology, 82, 170–174. doi:10.1080/
14620316.2007.11512215
MirAli, N.P., & Nabulsi, I. (2003). Genetic diversity of almonds (Prunus dulcis) using RAPD technique. Scientia Horticulturae, 98, 461–471. doi: 10.1016/S0304-4238(02)00200-5
Misirli, A., Saglam, H., Gulcan, R., Olmez, H.A., & Sahin, M. (2006). Investigation on fertilization biology of important dried apricot cultivars. Acta Horticulture, 701, 159–162. doi:10.17660/ActaHortic.2006.701.23
Mnejja, M., Garcia-Mas, J., Howad, W., & Arús, P. (2005). Development and transportability across Prunus species of 42 polymorphic almond microsatellites. Molecular Ecolology Notes, 5, 531–535. doi: 10.1111/j.1471-8286.2005.00977.x
Paydas, S., Eti, S., Gülcan, R., Derin, K., & Yilmaz, K.U. (2006). In vitro investigations on pollen quality, produc-tion and self-incompatibility of some apricots varieties in Malatya, Turkey. Acta Horticulture, 701, 75–80. doi:10.17660/ActaHortic.2006.701.8
Peakall, R., & Smouse, P.E. (2006). GENALEX 6: Genetic analysis in excel. Population genetic software for teach-ing and research. Molecular Ecology, 6, 288–295. doi:10.1111/j.1471-8286.2005.01155.x
Pedryc, A., Ruthner, S., Herman, R., Krska, B., Hegedus, A., & Halasz, J. (2009). Genetic diversity of apricot revealed by a set of SSR markers from linkage group
G1. Scientia Horticulture, 12, 19–26. doi:10.1016/j. scienta.2009.01.014
Rajia, R., Jannatizadehb, A., Fattahia, R., & Esfahlanica, M. A. (2014). Investigation of variability of apricot (Prunus armeniaca L.) using morphological traits and microsa-tellite markers. Scientia Horticulture, 176, 225–231. doi:10.1016/j.scienta.2014.06.033
Raz, A., Stern, R.A., Bercovich, D., & Goldway, M. (2009). SFB-based S-haplotyping of apricot (Prunus armeniaca) with DHPLC. Plant Breeding, 128, 707–711.
Rehder, A. (1967). Manual of cultivated trees and shrubs. NewYork: Macmillan.
Rholf, J.F. (2004). NTSYS-pc.: 2.11 numerical taxonomy and multivariate analysis system. New York: Stony Brook. Romero, C., Vilanova, S., Burgos, L., Martinez-Calvo, J.,
Vicente, M., Llacer, G., & Badanes, M.L. (2004). Analysis of the S locus structure in Prunus armeniaca L. identification of S-haplotype S-RNase and F-box genes. Plant Molecular Biology, 56, 145–157. doi:10.1007/s11103-004-2651-3
Sa´nchez-Pe´rez, R., Ruiz, D., Dicenta, F., Egea, J., & Martı ´nez-Go´mez, P. (2005). Application of simple sequence repeat (SSR) markers in apricot breeding: Molecular characterization, protection, and genetic relationships. Scientia Horticulture, 103, 305–315. doi:10.1016/j. scienta.2004.06.009
Sambrook, J., & Russell, D. (2001). Molecular cloning, a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Scheulke, M. (2000). An economic method for the fluor-escent labeling of PCR fragments. Nature Biotechnology, 18, 233–234. doi:10.1038/72708
Sosinski, B., Gannavarapu, M., Hager, L.D., Beck, L.E., & King, G.J. (2000). Characterization of microsatellite mar-kers in peach (Prunus persica L. Batsch). Theoretical and Applied Genetics, 101, 421–424. doi:10.1007/ s001220051499
Tao, R., Yamane, H., Sugiura, A., Murayama, H., Sassa, H., & Mori, H. (1999). Molecular typing of S-alleles through identification, characterization and cDNA cloning for S-RNases in sweet cherry. Journal of the American Society for Horticultural Science, 124, 224–233.
Tautz, D. (1989). Hypervariability of simple sequences as a general source for polymorphic DNA. Nucleic Acids Research, 17, 6463–6471. doi:10.1093/nar/17.16.6463
Testolin, R., Marrazo, T., Cipriani, G., Quarta, R., Verde, I., Dettori, T. Pancaldi, M., Sansavini, S. (2000). Microsatellite DNA in peach (Prunus persica L Batsch)
and it use in fingerprinting and testing the genetic origin of cultivars. Genome, 43, 512–520.
Topcu, H., Coban, N., & Kafkas, S. (2016). Novel micro-satellite markers in Pistacia vera L. and their transfer-ability across the genus. Pistacia Scientia Horticulturae, 198, 91–97. doi:10.1016/j.scienta.2015.11.012
Vilanova, S., Badenes, M.L., Burgos, L., Martinez-Calvo, J., Llacer, G., & Romero, C. (2006). Self-compatibility of two apricot selections is associated with two pollen-part mutations of different nature. Plant Physiology, 142, 629–641. doi:10.1104/pp.106.083865
Vilanova, S., Romero, C., Llacer, G., & Badenes, M.L. (2005). Identification of self-incompatibility aleles in apricot by PCR and sequence analysis. Journal of the American Society for Horticultural Science, 130, 893–898. Yilmaz, K.U. (2008). Phenological, morphological and pomological characteristics of some local apricot genotypes and determination of their genetic relationship and self-incompatibility by molecular markers (Phd Thesis),, Adana: Cukurova University.
Yilmaz, K.U., Kafkas, S., & Kargı Paydas, S. (2013). Determination of self-(in)compatibility in Turkish apri-cot genotypes. Fruit Science, 1, 34–40.
Yilmaz, K.U., Paydas Kargi, S., & Kafkas, S. (2012). Morphological diversity of the Turkish apricot (Prunus armeniaca L.) germplasm in the Irano-Caucasian eco-geographical group. Turkish Journal of Agriculture and Forestry, 36, 688–694.
Yuan, Z., Chen, X., He, T., Feng, J., Feng, T., & Zhang, C. (2007). Population genetic structure in apricot (Prunus armeniaca L.) cultivars revealed by fluorescent-AFLP markers in Southern Xinjiang, China. Journal of Genetics and Genomics, 34, 1037–1047. doi:10.1016/ S1673-8527(07)60117-9
Zaloglu, S., Kafkas, S., Dogan, Y., & Guney, M. (2015). Development and characterization of SSR markers from pistachio (Pistacia vera L.) and their transferability to eight Pistacia species. Scientia Horticulture, 189, 94– 103. doi:10.1016/j.scienta.2015.04.006
Zhang, L.J., Chen, X.S., Chen, X.L., Zhang, C.Y., Liu, X.L., Ci, Z.J., . . . Liu, C.Q. (2008). Identication of self-incom-patibility (S) genotypes of Chinese apricot cultivars. Euphytica, 160, 241–248. doi:10.1007/s10681-007-9544-x
Zhebentyayeva, T.N., Reighard, G.L., Gorina, V.M., & Abbott, A.G. (2003). Microsatellite (SSR) analysis for assessment of genetic variability in apricot. Theoretical and Applied Genetics, 106, 435–444.