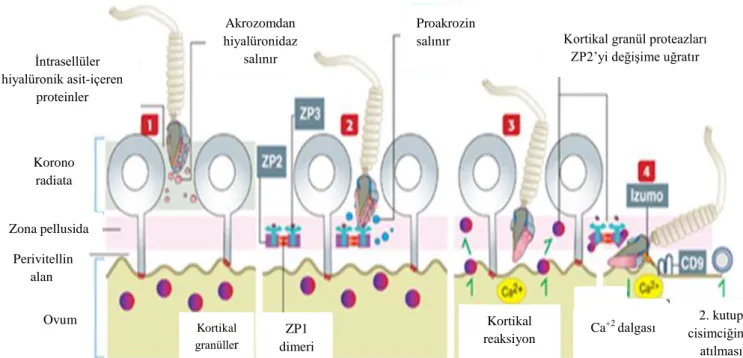
Normospermi ve astenospermi olgularında PLCζ’nın fertilizasyondaki etkisi
Tam metin
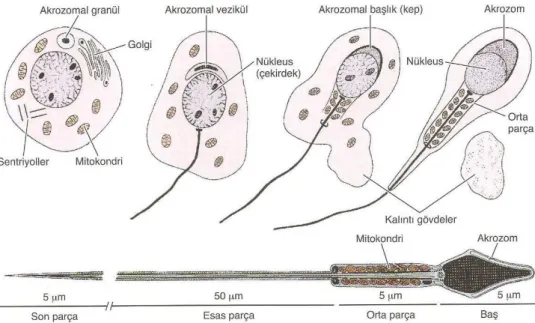
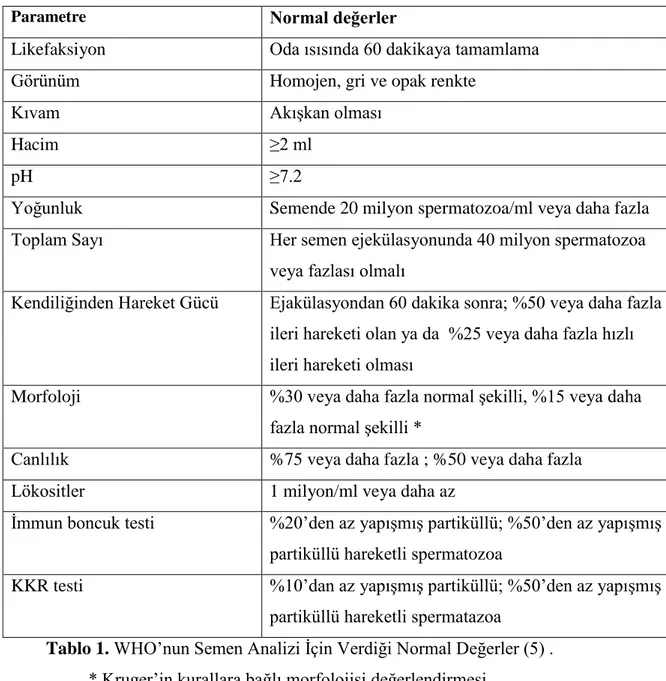
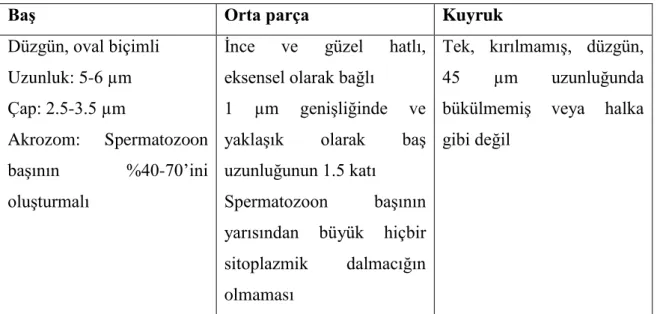
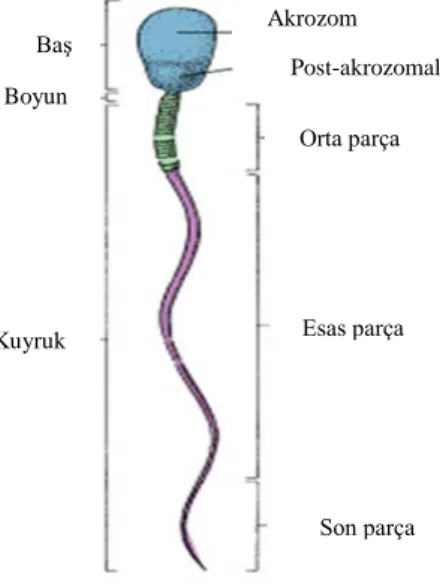
Şekil
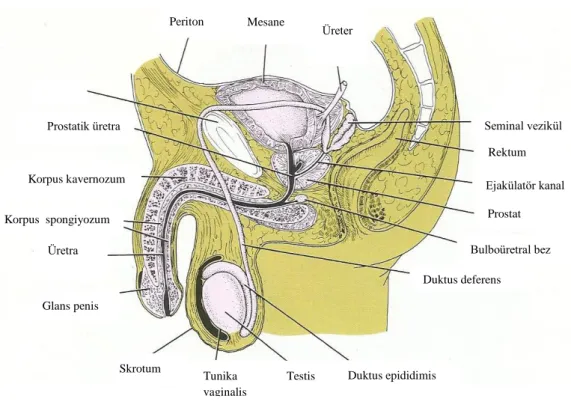
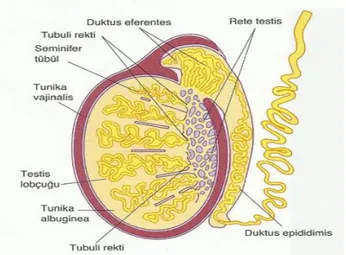
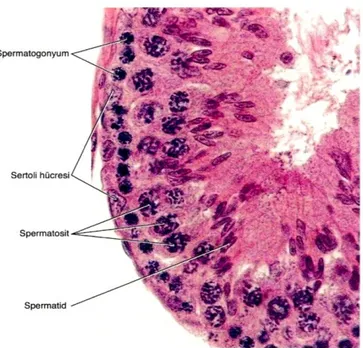
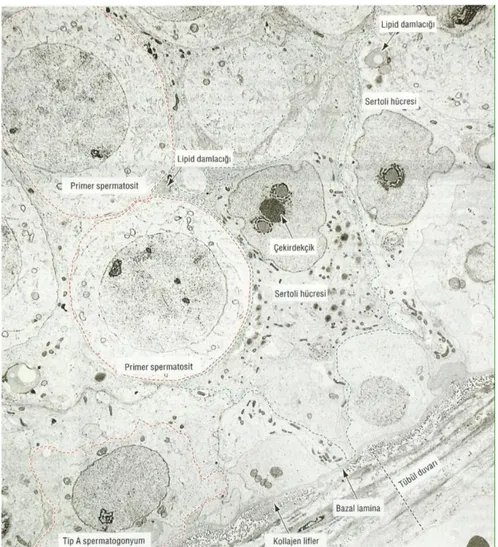

Outline
Benzer Belgeler
Ulaşılabilen literatürdeki B.henselae se- roprevalansı araştırmalarında, antijen kaplı lamların hazırlanmasında B.henselae’nın ço- ğunlukla Vero hücrelerinde, ikinci
Hawkins ve arkada§lan (11) yapt1klan bir gall§mada sagl1kl1 geng adOitlerin azot protoksite akut olarak maruz kalmalan halinde idrarlanndaki FIGLU miktannm iki kat
grubunda, eğitim görmemi ş anne leri n ve sosyoekonomik sevi yesi düş ük ailelerin çocuklarinda şehir şe beke suyu kullananlarda anti-HAY 19 G pozitifliği
2012 Sezonu Türkiye Üniversite Sporları Federasyonu Korumalı Futbol Ligi fikstür çekimine aşağıdaki Üniversiteler katılmış olup çekilen fikstüre göre gruplar ve
192 Au izotopuna ait Şekil 4.3 de verilen seviye şemasında Band-1 olarak adlandırılan ve 11 - band başlangıç spin-paritesine sahip bandda daha önce yapılan deneysel ve teorik
1 Renault Grubu, sabit parametre ve döviz kurlarına göre konsolide gelirlerdeki değişimi analiz etmek için önceki yılın yıllık ortalama döviz kurlarına başvurarak ve
Deneysel bir yaklaĢım değildir, her genetik hastalık/durum için ayrıca kanıtlanmıĢ ve tekrarlanabilen test protokolleri olan bir disiplindir Test sonuçlarını içeren
Sınıf / A Şubesi (İMAM HATİP PROGRAMI UYGULANAN ALAN) Sınıf Listesi.. Sınıf Öğretmeni: METİN KILIÇ
