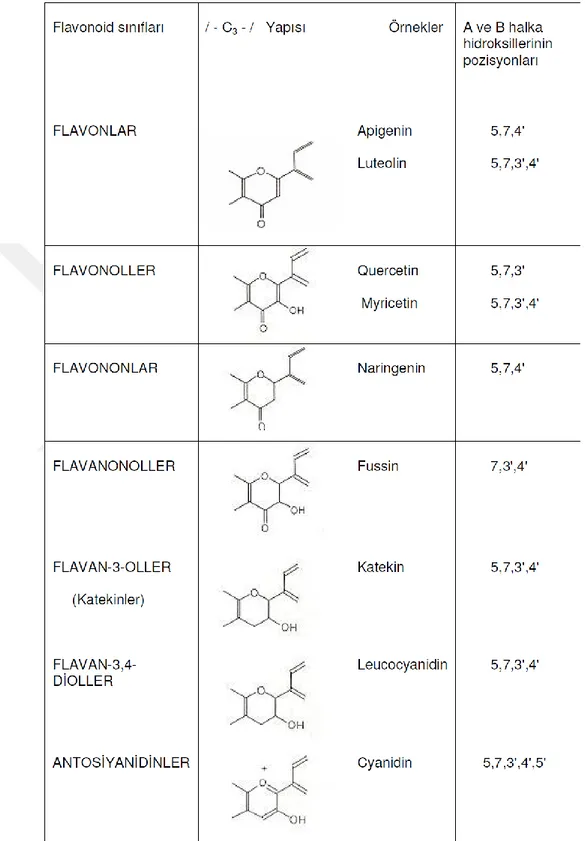
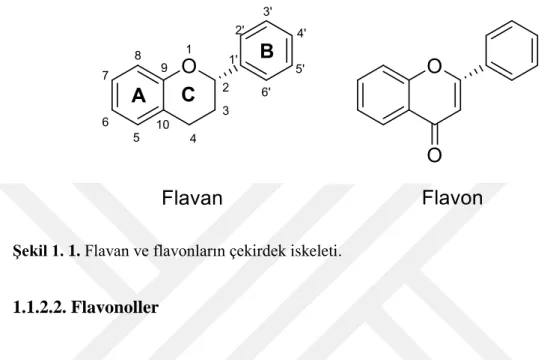
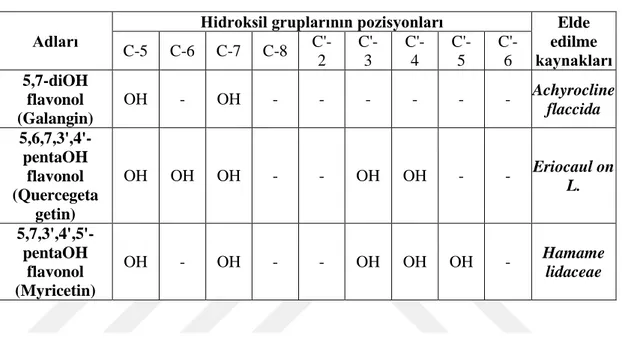
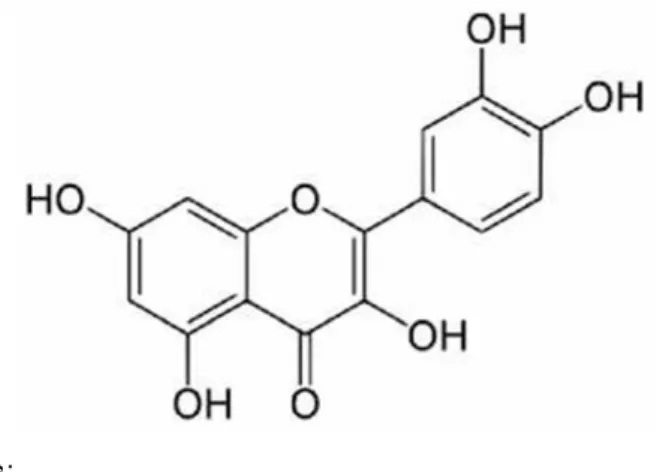
Bazı flavonoidlerin antioksidan kapasitelerinin belirlenmesi, asetilkolinesteraz ve karbonik anhidraz enzimleri üzerine etkilerinin incelenmesi
Tam metin
Şekil
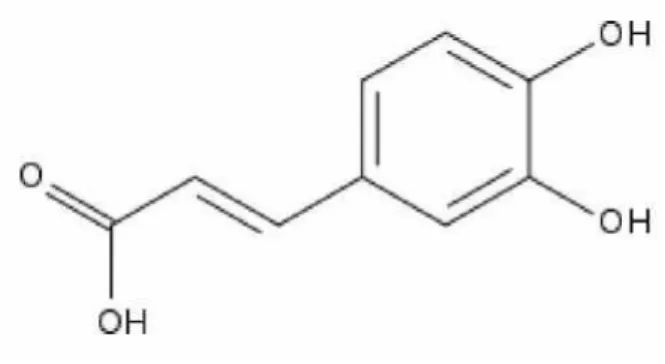




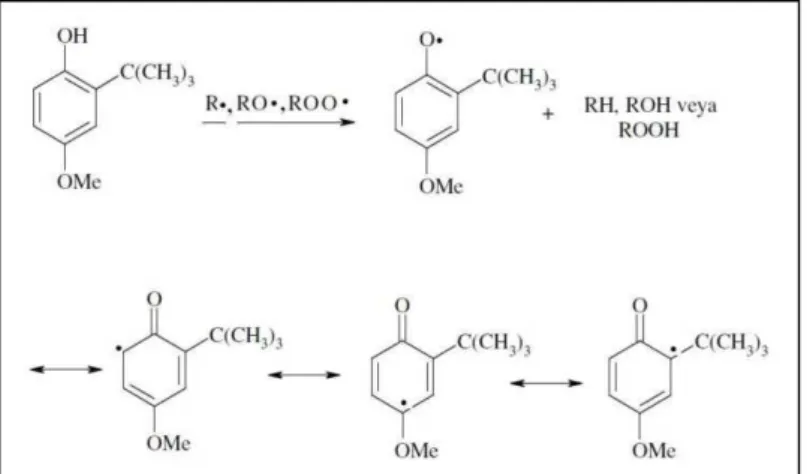
Benzer Belgeler
Fakat “bala üşüşen bu sinekler” (s. 90) gidip nişan veya kumar oyunları oynamakta çekirdek yiyip cinsel muhabbetler etmektedir. Kimse Ahmet’in yaptığı gösterinin ne
A Deniz Gezmiş, Yusuf Aslan ve Hüseyin inan ın 6 Mayıs 1972'de ^ idamları ilesonuçlanan davayı hükme bağlayan Yargıtay Da ireler Kurulu'nun üyesi emekli Hâkim Albay
An upper estimate for the Alexander polynomial of an algebraic curve is obtained, which sharpens Libgober’s estimate in terms of the local polynomials at the singular points of
Simutenkov, bu düzenlemelerin bir tarafta Avrupa Birliği Üye Devleti veya Avrupa Ekonomik Alanı vatandaşları ve diğer tarafta üye olmayan ülkelerin
1) It supports multipath routing and forwarding without looping or congestion problem due to interest/data packets flow-balance. This brings the idea of adaptive
In Section 2, we will construct a dynamic sys- tem model based on exact penalty function (Meng et al., 2004b). Moreover, we will prove that the system can converge to the
We have also performed quasi-real time SHPM imaging to study how the vortices penetrate into the BSCCO superconductor. We first degaussed the system at 110 K and then applied
sınıf Fen ve Teknoloji dersi programında öngörülen kavramların kazandırılma düzeyini saptamak için, bilgi düzeyi, kavrama düzeyi ve toplam erişi düzeyi
