Correction to: diagnostic utility of a targeted next-generation sequencing gene panel in the clinical suspicion of systemic autoinflammatory diseases: a multi-center study (vol 39, pg 911, 2019)
Tam metin
Şekil
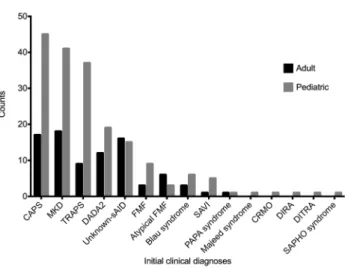
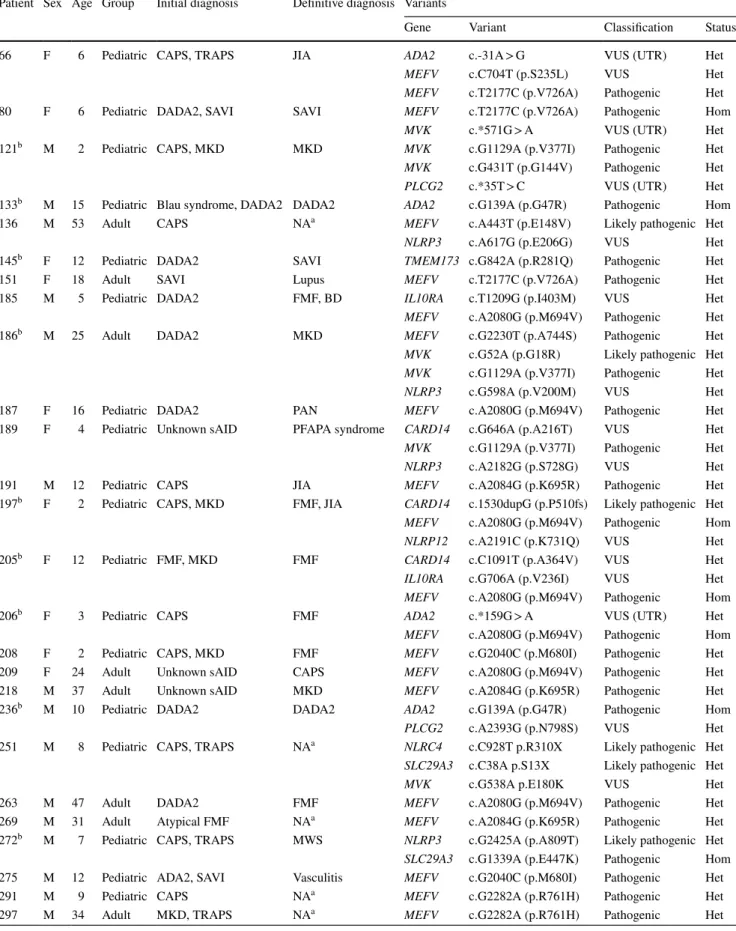
Benzer Belgeler
Tercümenin aslına tam uygun ve doğru olması, temiz ve yanlışsız bir dille yapılması sizin ve Üniversitenin şerefi demek olduğunu bir kerre daha önemle
Kaplan’a göre Türk milletinin kültürel değerleri şunlardır: Türk Dili, Türk Ede- biyatı, Türk Tarihi, Türk Mûsikîsi, Türk Plâstik Sanatları, Türk Şehirleri
Sonuç olarak akut a¤r›l› hastalarda tüm enjeksiyon fle- killerinde gruplar aras›nda fark bulunmad›¤›n›, metil prednizo- lon ilave edilen lokal anestezik ve serum
Farklı HP oranlarına sahip rasyonlarla beslenen 3 farklı grup- taki ineklerin, östrus günündeki pH değerleri arasında belirlenen farklılık istatistiki açıdan oldukça önemli
If a sequence in a quasi-metric space (X, ρ) is left ρ-statistically convergent (left K-statistical Cauchy, weakly left K-statistical Cauchy, left ρ-statistical Cauchy), then it has
It is for sure that the rapid change in the Congo from a dependent colony to a free state, did not fit in with the improvement of the perspectives of some Belgians, which
effects of biosorbent dosage, biosorption time, solution pH, initial zinc and copper ions concentration, humic acid or ionic strength or competitive effects on the biosorption
