Automatic characterization of copy number polymorphism using high throughput sequencing
Tam metin
Şekil
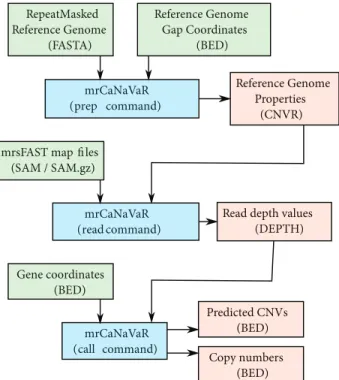
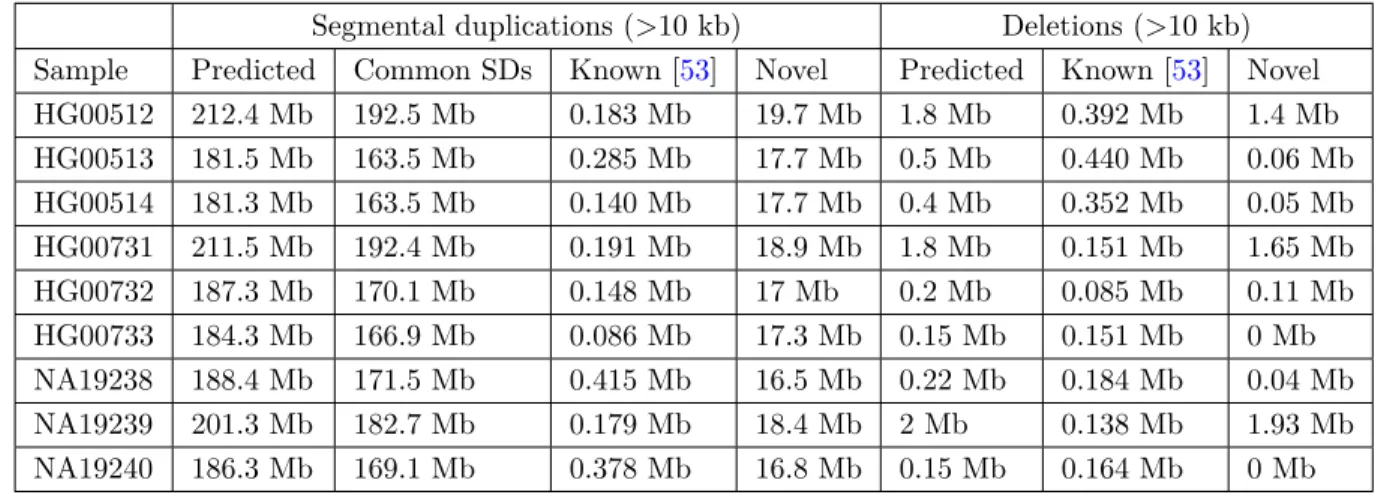
Benzer Belgeler
This thesis presents the basic design education literature in terms of digital approaches, virtual reality technologies, usability criteria and also technology acceptance model..
In order to explain the asymmetric transmission, we investigated the induced surface currents at 6.2 GHz, when the CMM is excited by x-polarized waves propagating in the
We observe that a larger rate region can be obtained by using non-uniform input distributions for both users, and motivated by this observation, we propose an encoding scheme based
In our method, the fingerprint image is first processed by a binary nonlinear subband decomposition filter bank and the resulting subimages are coded using vector quantizers
The in-situ transmittance response depicts that the photo-bleaching response is 6.2 s for 70% transmittance change under 100 mW/cm 2 illumination in short circuit configuration, and
After studying a wide range of common Internet attacks that require some sort of user interaction or decision (e.g., reflected cross-site scripting attacks, advertisement trick
Daß der Prinz sich den Thron verdient und erfolgreich Macht aus übt, ist in erster Linie für den Padischah wichtig, der ihm den Thron überläßt - aber auch für den Prinzen, dem
This measurements show that the nanocrystals efficiently transfer energy to dye molecules and consequently quench, with their lifetimes decreasing successively, and reaching
