T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
TÜRKĠYE'DE YETĠġTĠRĠLEN BAZI KEÇĠ IRKLARINDA BÜYÜME HORMONU GENĠ (GH-1), ĠNSÜLĠN BENZERĠ BÜYÜME FAKTÖRLERĠ (IGF-1) VE PĠTÜĠTER SPESĠFĠK TRANSKRĠPSĠYON
FAKTÖRÜ-1 (PĠT-1) GENLERĠ POLĠMORFĠZMLERĠ
MAHAMAT ALĠ AMĠNE OUCHAR DOKTORA TEZĠ
Zootekni Anabilim Dalı
Aralık - 2019 KONYA Her Hakkı Saklıdır
i
TEZ KABUL VE ONAYI
Mahamat Ali Amine OUCHAR tarafından hazırlanan “Türkiye'de YetiĢtirilen Bazı Keçi Irklarında Büyüme Hormonu geni (GH1), Ġnsülin Benzeri Büyüme faktörleri (IGF-1) ve Pitüiter Spesifik Transkripsiyon Faktörü-1 (PĠT-1) Genleri Polimorfizmleri” adlı tez çalıĢması 26/12/2019 tarihinde aĢağıdaki jüri üyeleri tarafından oy birliği / oy çokluğu ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü Zootekni Anabilim Dalı’nda DOKTORA TEZĠ olarak kabul edilmiĢtir.
Jüri Üyeleri Ġmza
BaĢkan
Prof. Dr. Fatin CEDDEN ………..
DanıĢman
Prof.Dr. Birol DAĞ ………..
Üye
Prof.Dr. Saim BOZTEPE ………..
Üye
Prof.Dr. Ercan KURAR ………..
Üye
Doç. Dr. Ġbrahim AYTEKĠN ………..
Yukarıdaki sonucu onaylarım.
Prof. Dr. Mustafa YILMAZ FBE Müdürü
Bu tez çalıĢması S.Ü. BAP Koordinotörlüğü tarafından 17201119 nolu proje ile desteklenmiĢtir.
ii
TEZ BĠLDĠRĠMĠ
Bu tezdeki bütün bilgilerin etik davranıĢ ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalıĢmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all materials and results that are not original to this work.
Mahamat Ali Amine OUCHAR Tarih: 26.12.2019
iii ÖZET DOKTORA TEZĠ
Türkiye'de YetiĢtirilen Bazı Keçi Irklarında Büyüme Hormonu Geni (GH-1), Ġnsülin Benzeri Büyüme Faktörleri (IGF-1) ve Pitüiter Spesifik Transkripsiyon Faktörü-1
(Pit-1) Genleri Polimorfizmleri
Mahamat Ali AMĠNE OUCHAR Selçuk Üniversitesi Fen Bilimleri Enstitüsü
Zootekni Anabilim Dalı DanıĢman: Prof. Dr. Birol DAĞ
2019, 63 Sayfa Jüri
Prof. Dr. Birol DAĞ Prof. Dr. Saim BOZTEPE
Prof. Dr. Ercan KURAR Prof. Dr. Fatin CEDDEN Doç. Dr. Ġbrahim AYTEKĠN
Bu araĢtırmada büyümeye ve geliĢmeye etkili olan ve kas geliĢmesinin düzenlenmesinde memelilerde aday gen olarak kabul edilen Hipofiz Spesifik Transkripsiyon Faktörü-1 (Pituitary-specific Transcription Factor; Pit-1) geninin ekzon 6, (Insulin Like Growth Factor-1; IGF-Pit-1) geninin ekzon 4 ve intron 4 ve (Growth Hormone-1; GH-1) geninin ekzon 2 ve 3 bölgelerinin polimorfizm bakımından incelenmesi amaçlanmıĢtır. Elli iki baĢ Kıl, 51 baĢ Ankara keçisi, 50 baĢ Kilis, 50 baĢ Honamlı, 50 baĢ Halep ve 50 baĢ Saanen keçisi olmak üzere 6 farklı ırktan oluĢan toplam 303 baĢ keçide GH-1 geni HaeIII, IGF-1 HaeIII ve Pit-1 PstI restriksiyon enzimleri ile kesilmiĢtir. PCR-RFLP (Polimeraz Zincir Reaksiyonu ve Restriksiyon Parça Uzunluk Polimorfizmi) kullanılarak Pit-1 geni PstI polimorfizmi (450 bç), IGF-1 geni HaeIII polimorfizmi (363 bç) ve GH-1 geni HaeIII polimorfizmi (422 bç) ile genotiplendirilmiĢtir.
Pit-1 geni PstI polimorfizmi bakımından T ve C allel frekansları Kıl, Ankara, Honamlı, Halep, Sannen ve Kilis ırklarında sırasıyla 0.9038 ve 0.0962, 0.902 ve 0.098, 0.8900 ve 0.1100, 0.9300 ve 0.0700, 0.8300 ve 0.1700 ile 0.9100 ve 0.0900 olarak tespit edilmiĢtir. Bütün ırklarda CC genotipi tespit edilmemiĢ olup, TT ve TC genotip frekansları ise sırasıyla Kıl keçilerinde 0.808 ve 0.192 (P>0.05); Ankara keçilerinde 0.804 ve 0.196 (P>0.05); Honamlı keçilerinde 0.780 ve 0.220 (P>0.05); Halep keçilerinde 0.860 ve 0.140 (P>0.05); Saanen keçilerinde 0.660 ve 0.340 (P>0.05)ve Kilis keçilerinde 0.820 ve 0.180 (P>0.05) olarak bulunmuĢtur. Genel ırklar dikkate alındığında T ve C allel frekanslarını 0.8944 ve 0.1056, TT, TC ve CC genotip frekanslarını ise 0.789, 0.211 ve 0.000 olarak tespit edilmiĢtir (P<0.05). Pit-1 geni ekzon 6 bölgesi PstI polimorfizmi bakımından ırklar bazında genel değerlendirme yapıldığında populasyonun Hardy-Weinberg dengesinde olmadığı belirlenmiĢtir (P<0.05).
IGF-1 geni HaeIII enzim kesimi sonucu iki allel (A ve B) ve üç genotip (AA, BB ve AB) belirlenmiĢtir. Kıl, Ankara, Honamlı, Halep, Saanen ve Kilis keçi ırkları için A ve B allel frekansları sırasıyla 0.3558 ve 0.6442, 0.5784 ve 0.4216, 0.2100 ve 0.7900, 0.7300 ve 0.2700, 0.7800 ve 0.2200, 0.2900 ve 0.7100 olarak bulunmuĢtur. AA, AB ve BB genotip frekansları bakımından ise sırasıyla 0.250, 0.212 ve 0.538 (P<0.05); 0.451, 0.255 ve 0.294 (P<0.05); 0.060, 0.300 ve 0.640 (P>0.05); 0.600, 0.260 ve 0.140 (P<0.05); 0.700, 0.160 ve 0.140 (P<0.05); 0.200, 0.180 ve 0.620 (P<0.05) olarak bulunmuĢtur.. Bütün ırklar dikkate alındığında A ve B allel frekansları
iv
0.4901 ve 0.5099, AA, AB ve BB genotip frekansları ise 0.376, 0.228 ve 0.396 (P<0.05) olarak tespit edilmiĢtir. ÇalıĢılan ırklarda populasyonun Hardy-Weinberg dengesinde olmadığı belirlenmiĢtir (P<0.05).
GH-1 geninin HaeIII enzimi ile kesimi sonucu iki allel (A ve B) ve üç genotip (AA, BB ve AB) belirlenmiĢtir. Kıl, Ankara, Honamlı, Halep, Saanen ve Kilis keçi ırkları için A ve B allel frekansları sırasıyla 0.4038 ve 0.5962, 0.4314 ve 0.5686, 0.4600 ve 0.5400, 0.4500 ve 0.5500, 0.3800 ve 0.6200, 0.5400 ve 0.4600 olarak bulunmuĢtur. AA, AB ve BB genotip frekansları bakımından ise sırasıyla 0.019, 0.769 ve 0.212 (P<0.05); 0.235, 0.392 ve 0.372 (P>0.05); 0.080, 0.760 ve 0.160 (P<0.05); 0.060, 0.780 ve 0.160 (P<0.05); 0.160, 0.440 ve 0.400 (P>0.05) ve 0.160, 0.760 ve 0.080 (P<0.05) olarak bulunmuĢtur. Bütün ırklar dikkate alındığında A ve B allel frekansları 0.4439 ve 0.5561, AA, AB ve BB genotip frekansları ise 0.119, 0.650 ve 0.231 (P<0.05) olarak tespit edilmiĢtir. Genel populasyonun Hardy-Weinberg dengesinde olmadığı belirlenmiĢtir (P<0.05).
v
ABSTRACT Ph.D THESIS
Genetic Polymorphism of Growth Hormone (GH-1), Insulin-like Growth Factor-1 (IGF-1) and Pituitary-Specific Transcription Factor-1 (Pit-1) Genes Polymorphism in
Some Goat Breeds Reared in Turkey
Mahamat Ali AMĠNE OUCHAR
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY THE DEGREE OF DOCTOR OF PHILOSOPHY IN
ANIMAL SCIENCE
Advisor: Prof. Dr. Birol DAĞ 2019, 63 Pages
Jury
Prof. Dr. Birol DAĞ Prof. Dr. Saim BOZTEPE
Prof. Dr. Ercan KURAR Prof. Dr. Fatin CEDDEN Doç. Dr. Ġbrahim AYTEKĠN
The purpose of this study is to determine Pituitary-specific Transcription Factor-1 (Pit-1) gene exon 6, insulin-like growth factors (IGF-1) gene exon 4 and intron 4 and growth hormone (GH-1) gene exon 2 and 3 regions in terms of polymorphism. Animal materials of the study consist of a total of 303 goats including 52 Hair, 51 Angora, 50 Kilis, 50 Honamlı, 50 Aleppo and 50 heads of Saanen breeds. GH-1 and IGF-1genes were cut with HaeIII and Pit-1 gene were cut with PstI restriction enzymes. Pit-1 gene PstI polymorphism (450 bp), IGF-1 gene HaeIII polymorphism (363 bp) and GH-1 gene HaeIII polymorphism (422 bp) were genotyped by using PCR-RFLP (Polymerase Chain Reaction and Restriction Fragment Length Polymorphism)
Pit-1 gene in terms of PstI polymorphism, T and C allele frequencies were determined as 0.9038 and 0.0962; 0.902 and 0.098; 0.8900 and 0.1100; 0.9300 and 0.0700; 0.8300 and 0.1700 and 0.9100 and 0.0900 for Hair, Angora, Kilis, Honamlı, Aleppo and Saanen breeds respectively. CC genotype was not detected in all breeds. TT and TC genotype frequencies were found to be 0.808 and 0.192 in Hair goats (P>0.05), 0.804 and 0.196 (P>0.05) in Ankara goats; 0.780 and 0.220 (P>0.05) in Honamlı goats; 0.860 and 0.140 (P>0.05) in Aleppo goats; respectively 0.660 and 0.340 (P>0.05) in Saanen goats and 0.820 and 0.180 (P>0.05) in Kilis goats respectively. When the general breeds were taken into consideration, T and C allele frequencies were found to be 0.8944 and 0.1056, and TT, TC and CC genotype frequencies were 0.789, 0.211 and 0.000 (P<0.05). In terms of PstI polymorphism in the exon 6 region of the Pit-1 gene, it was determined that the population was not in Hardy-Weinberg equilibrium (P<0.05).
IGF-1 gene HaeIII restriction enzymes results were identified two alleles (A and B) and three genotypes (AA, BB and AB). A and B allele frequencies for Hair, Angora, Honamlı, Aleppo, Saanen and Kilis breeds were found to be 0.3558 and 0.6442, 0.5784 and 0.4216, 0.2100 and 0.7900, 0.7300 and 0.2700, 0.7800 and 0.2200, 0.2900 and 0.7100, respectively. AA, AB and BB genotype frequencies were found to be 0.250, 0.212 and 0.538 (P<0.05), 0.451, 0.255 and 0.294 (P<0.05), 0.060, 0.300 and 0.640 (P>0.05), 0.600, 0.260 and 0.140 (P<0.05), 0.700, 0.160 and 0.140 (P<0.05), 0.200, 0.180 and 0.620 (P<0.05) respectively. When all breeds were taken into
vi
consideration, A and B allel frequencies were found to be 0.4901 and 0.5099, and AA, AB and BB genotype frequencies were found to be 0.376, 0.228 and 0.396 (P<0.05). It was determined that population was in not Hardy-Weinberg equilibrium (P<0.05).
GH-1 gene HaeIII restriction enzymes results were identified two alleles (A and B) and three genotypes (AA, BB and AB). A and B allel frequencies for Hair, Angora, Honamlı, Aleppo, Saanen and Kilis breeds were found to be 0.4038 and 0.5962, 0.4314 and 0.5686, 0.4600 and 0.5400, 0.4500 and 0.5500, 0.3800 and 0.6200, 0.5400 and 0.4600, respectively. AA, AB and BB genotype frequencies were found to be 00.019, 0.769 and 0.212 (P<0.05), 0.235, 0.392 and 0.32 (P>0.05), 0.080, 0.760 and 0.160 (P<0.05), 0.060, 0.780 and 0.160 (P<0.05), 0.160, 0.440 and 0.400 (P>0.05), 0.160, 0.760 and 0.080 (P<0.05), respectively. When all breeds were taken into consideration, A and B allel frequencies were found to be 0.4439 and 0.5561, and AA, AB and BB genotype frequencies were found to be 0.119, 0.650 and 0.231 (P<0.05). It was determined that the population studied was not in Hardy-Weinberg equilibrium (P<0.05).
vii ÖNSÖZ
Öncelikle bana kazandırdığı değerli bilgiler, tüm doktora eğitim süresinde maddi ve manevi olarak verdiği destek, emek ve gösterdiği hoĢgörüden dolayı sayın hocam Prof. Dr. Birol DAĞ’a teĢekkürü bir borç bilirim.
Tezimin Ģekillenmesinde değerli bilgileri ile katkıda bulunan tez izleme komite üyelerim sayın Prof. Dr. Saim BOZTEPE ve sayın Prof. Dr. Ercan KURAR’a yürekten teĢekkür ederim. AraĢtırmada Güneysınır ve Sızma'da materyal temininde yardımcı olan Konya ili Damızlık Koyun Keçi YetiĢtiricileri Birliği yönetimine ve doktora öğrenimim boyunca bana burs desteği sağlayan YurtdıĢı Türkiye ve Akraba Topluluklar BaĢkanığı'na (YTB) ve Selçuk Üniversitesi TÖMER'de bana Türkçe öğreten sayın hocam Meral DOĞRU'ya teĢekkürü bir borç bilirim.
Tez çalısmalarının gerçekleĢmesinde sağladığı katkıdan dolayı Selçuk Üniversitesi Bilimsel AraĢtırma Projeleri Koordinatörlüğü'ne, tezimde kullandığım kan örneklerinin alınmasından hazırlanmasında yardımcı olan Ankara Üniversitesi Ziraat Fakültesi Zootekni Bölümü Biyometri ve Genetik Anabilim Dalı BaĢkanı sayın Prof. Dr. Mehmet Ali YILDIZ'a ne kadar teĢekkür etsem azdır. Ayrıca tez projem kapsamındaki tüm laboratuvar çalıĢmalarımda özveri ile yardımcı olan, laboratuvar analizleri ilgili pratik kazanmamı sağlayan ve laboratuvarda karĢılaĢtığım birçok sorunun çözümünde bana yol gösteren, mesai harcayarak destek olan çok değerli hocam Doç. Dr. Ġbrahim AYTEKĠN, Özcan ġahin ve arkadaĢım Dr. Mervan BAYRAKTAR’a minnettarım. Ayrıca çalıĢma yerim Hayvancılık GeliĢtirme AraĢtırma Enstitüsü (IRED) adına yöneticilerine, diğer Çadlı öğrenci arkadaĢlarıma, Zootekni bölümünde bütün hocalarım ve BAF Premix Ģirket yöneticilerine tüm kalbimle teĢekkür ederim.
Bu uzun yolun sonuna geldim, baĢta hocalarım olmak üzere bu eğitim süresi boyunca bana yardım eden herkese teĢekkür ederim, iyi ki Türkiye'ye geldim, iyi ki bu bayrağın altında okudum, okul eğitimimin yanı sıra, en çok da misafirperverlik, yardımlaĢma, vatan ve bayrak sevgisi nedir bu ülkede öğrendim, Türkiye'ye sonsuz teĢekkür ederim.
Akademik hayatımın baĢlamasından itibaren bana sabır ve ümit verici bir tutumla her zaman desteklerini esirgemeyen babamın ruhu, annem, eĢim, oğlum, kızım ve bütün kardeĢlerim’e bu çalıĢmayı atfeder, Çad Cumhuriyeti hayvancılığına ve genetik geliĢmesine katkı sağlalayacağı düĢüncesi ile saygılar sunarım.
Mahamat Ali AMĠNE OUCHAR KONYA-2019
viii ĠÇĠNDEKĠLER TEZ BĠLDĠRĠMĠ ... ii ÖZET ... ...iii ABSTRACT ... v ÖNSÖZ ... vii ĠÇĠNDEKĠLER... viii SĠMGELER VE KISALTMALAR... x 1. GĠRĠġ ... 1 2. KAYNAK ARAġTIRMASI ... 6
2.1. DNA Polimorfizm Teknikleri ... 6
2.2. Türkiye Yerli Keçi Irkları ve Önemi ... 7
2.2.1. Kıl Keçisi ... 7
2.2.2. Ankara Keçisi (Tiftik Keçisi ... 8
2.2.3. Kilis Keçisi ... 9
2.2.4. Honamlı Keçisi ... 10
2.2.5. Saanen Keçisi ... 11
2.2.6. Halep Keçisi ... 12
2.3. Pit-1 Geni Polimorfizmi ... 12
2.4. IGF-1 Geni Polimorfizmi ... 15
2.5. GH-1 Geni Polimorfizmi ... 21 3. MATERYAL VE YÖNTEM ... 28 3.1. Materyal ... 28 3.1.1. Hayvan materyali ... 28 3.1.2. Kan örnekleri ... 28 3.2. Yöntem ... 29
3.2.1. Genomik DNA izolasyonu ... 29
ix
3.2.3. PCR ile DNA çoğaltımı... 30
3.2.4. PCR ürünlerinin restriksiyon enzimleri ile kesilmesi ... 33
3.2.5. Agaroz jel elektroforez uygulamaları ... 34
3.2.6. Elektroforez ile DNA bantlarının ayrıĢtırılması ve genotipleme ... 34
3.2.7. Ġstatistiksel analizler ... 35
4. ARAġTIRMA BULGULARI VE TARTIġMA ... 36
4.1. Moleküler genetik çalıĢmaları ... 36
4.1.1. Genomik DNA örneklerinin spektrofotometre sonuçları ... 36
4.1.2. Pit-1, IGF-1 ve GH-1 genlerindeki restriksiyon enzimlerinin kesim bölgelerine göre allel ve genotip frekansları ile χ² testi sonuçları ... 37
4.1.2.1. Pit-1 ekzon 6 bölgesi PstI (450 bç) polimorfizm sonuçları ... 37
4.1.2.2. IGF-1 ekzon 4 ve Ġntron 4 bölgesi HaeIII (363 bç) polimorfizm sonuçları ... 42
4.1.2.3. GH-1 ekzon 2 ve 3 bölgesi HaeIII (422 bç) polimorfizm sonuçları ... 48
5. SONUÇLAR VE ÖNERĠLER ... 55
KAYNAKLAR ... 57
x
SĠMGELER VE KISALTMALAR TÜĠK Türkiye Ġstatistik Kurumu
FAO Birlesmis Milletler Gıda ve Tarım Teskilatı MAS Markör Destekli Seleksiyon
QTL Kantitatif Karakter Lokusu
RFLP Restriksiyon Fragman Uzunluk Polimorfizmi SNP Tek nükleotit deĢiĢimi
SSR Basit Tekrar Dizileri
AFLP ÇoğaltılmıĢ Fragman Uzunluk Polimorfizmi RAPD Rastgele ÇogaltılmıĢ Polimorfik DNA PCR Polimeraz zincir reaksiyonu
DNA Deoksiribonükleik asit UV Ultraviyole kDa Kilodalton cM Santimorgan μl Mikrolitre ml Mililitre o C Santigrat derece M Molar mM Milimolar g Gram kg Kilogram nm Nanometre OD Optik Densite dNTPs Deoksiribonükleozid trifosfat A Adenin nükleotid T Timin nükleotid G Guanin nükleotid C Sitozin nükleotid bç Baz çifti kb Kilo baz dk Dakika sn Saniye MgCl2 Magnezyum klorür TE Tris-EDTA EDTA Ethylendinitrilotetraaceticacid TBE Tris-Borate-EDTA U Ünite μS Mikro siemens m3 Metre küp cm Santi metre χ² Kikare Testi RNA Ribonükleoik asit mRNA Haberci RNA TD Tablo değeri
BH Büyüme Hormonu
IGF-I Insulin-like Growth Factor-I, Ġnsülin Benzeri Büyüme Faktörü-I
Pit-1 Pituitary-Specific Transcription Factor-1, Hipofize Özgü Transkripsiyon Faktörü-1 POU1F1 Pituitary-specific positive transcription factor 1, Hipofiz özgül transkripsiyon faktör 1
1. GĠRĠġ
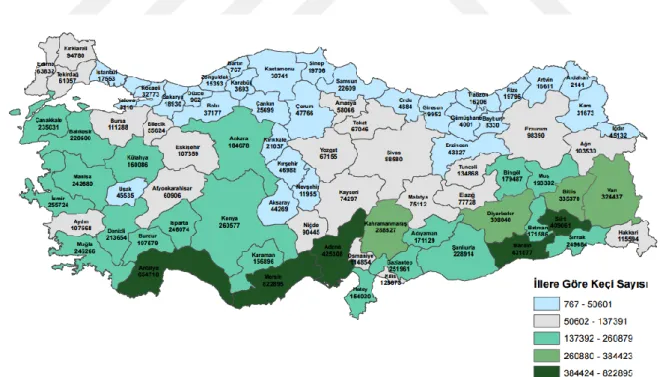
Türkiye’de keçi yetiĢtiriciliği denildiğinde akla ilk olarak kıl keçileri gelmektedir. Keçi varlığı 10 992 427 milyon baĢ olup, toplam keçi varlığının yaklaĢık %98’ini Kıl keçi ve melezleri, %2’sini ise Tiftik keçileri oluĢturmaktadır. Çiftlik hayvanları içerisinde payı ise %17.35’tır (Anonim, 2018). Koyun ve keçi yetiĢtiriciliği, genel olarak zayıf meralar, nadas, anız ve bitkisel üretime uygun olmayan alanları değerlendirerek et, süt, yapağı, kıl ve deri gibi ürünlere dönüĢtüren bir faaliyettir. Türkiye'de doğal kaynaklar, özellikle çayır meralar küçükbaĢ türlerinin yetiĢtiriciliği için daha uygundur. Kırsal nüfusun tüketim alıĢkanlıkları gibi nedenler, koyun ve keçi yetiĢtiriciliği için uygundur. (Kaymakcı ve Sönmez, 1996) KüçükbaĢ yetiĢtirmenin önemine rağmen, son yıllarda koyun ve keçi sayısında önemli bir azalma gözlenmiĢ olsa da son yıllarda bu alana yönelik teĢvik ve ıslah projeleri ile bir miktar da olsa iyileĢme sağlanmaktadır. Keçi yetiĢtiriciliği, genellikle geliĢmekte olan ve geliĢmiĢ ülkelerde gerçekleĢtirilen geleneksel bir hayvan üretim koludur. Ormanlarda ve kırsal alanlardaki yetersiz gelirli aileler için önemli bir geçim ve besin kaynağıdır. Keçi yetiĢtiriciliğinin diğer bir avantajı da et ve süt gibi ürünler üretmek için baĢka Ģekilde değerlendirilemeyen dağlık, taĢlık ve fundalık alanların kullanılmasıdır (Kaymakcı, 1997). ġekil 1.1'de Türkiye'de illere göre yetiĢtirilen keçi sayıları verilmiĢtir.
Çiftlik hayvanlarında verim artıĢı genotipin ve/veya çevrenin iyileĢtirilmesi ile mümkün olacağından, populasyonların genetik yapıları hakkında bilgi sahibi olunması damızlık seçiminde baĢarıyı artırır. Özellikle polimorfik özellikler ile üzerinde durulan özellikler arasında yüksek bir korelasyon olması, erken dönemde tespitinin yapılabilirliği ve cinsiyete bağlı olmama gibi kriterlerin varlığı seleksiyonda yetiĢtiricilere kayda değer bir katkı sağlamaktadır. Günümüze kadar çiftlik hayvanlarında birçok aday gen çalıĢılmıĢ ve bu genler ile ekonomik önemi olan verim özellikleri arasındaki iliĢkiler araĢtırılmıĢtır. Aday genler, fizyolojik veya biyokimyasal özelliklerdeki farklılıklar ve verim özellikleri arasındaki iliĢkiye dayanarak seçilmektedir (Çizelge 1.1). Bu genlerin düzenleyici veya yapısal bölgelerindeki allel farklılıklarının varlığı, verim kalitesi ve miktarında niteliğinde farklılaĢmalara yol açabilmektedir. Aslında, bu bölgelerdeki nükleotitlerin polimorfizmi, gen ekspresyonununu etkileyebilmektedir. Bununla birlikte intron sekanslarındaki bu farklılıkların, genetik markerler olarak potansiyel kullanımları olduğu da bildirilmiĢtir (Soller, 1983).
Çizelge 1.1. Keçilerde aday genlerin önemli ekonomik özelliklerde yeri (Supakorn, 2009)
Aday gen
Yer Fonksiyon Kaynak
GH 19q22 Büyüme ve süt verimi (Missohou ve ark., 2006)
GHR 20q17 Süt verimi, ana süt bileĢenlerinin içeriği ve somatik hücre sayımı
(Maj ve ark., 2007).
IGF-I 5q31 Büyüme, metabolizma ve üreme (Lan ve ark., 2007a)
LEP 3q33 Büyüme geliĢimi, yem dönüĢüm etkinliği ve süt
(Whitley ve ark., 2005)
POU1F1 1q21-q22 Büyüme, karkas, süt ve yün (Lan ve ark., 2007a)
MSTN - Kas geliĢtirme (Li ve ark., 2006)
BMP
- Embriyonik geliĢim, homeostaz, çeĢitli doku desenlerinin onarımı, hücre farklılaĢması, apoptoz ve dıĢı fertilitesi
(Fang ve ark., 2009)
FOXL2 - DiĢilerde Follikül oluĢumu ve oktivasyonu. (Uhlenhaut ve Treier, 2006)
MTNR1A - Üreme mevsimsellik (Migaud ve ark., 2002)
SRY Y Cinsiyet belirleme (Shi ve ark., 2008)
AMEL - Cinsiyet belirleme (Chen ve ark., 2007)
Casein 6q32 Süt verimi ve süt kompozisyonu (Othman ve Ahmed, 2007)
KAP 5q25 Elyaf / yün tekstili (Zhao ve ark., 2008)
MC1R - Elyaf / yün rengi (Cecchi ve ark., 2007)
MHC 23q22 BağıĢıklık sistemi (Ahmed ve Othman, 2006)
PrP 13q17 Skrapi direnci (Babar ve ark., 2009)
Moleküler markerler, çiftlik hayvanlarında DNA seviyesindeki genetik çeĢitliliği tespit etmek, gen bölgelerini ekonomik açıdan önemli karakterler Quantitave Trait Loci- Kantitatif Karakter Locusu (QTL) iliĢkisini bulmak veya herhangi bir avantajı etkileyen tek bir geni (majör gen) tespit etmek için yaygın olarak kullanılmaktadır (Ağaoğlu, 2010). Markörler ile QTL arasındaki iliĢki Marker Assisted Selection = Marker Destekli Seleksiyon (MAS) için kullanılabilir. MAS tek baĢına veya klasik yöntemleriyle birlikte, özellikle büyükbaĢ ve küçükbaĢ gibi generasyonlar arası süresi uzun olan türlerde daha önemlidir.
Aday gen yaklaĢımı, araĢtırmacılara bir özelliğin genetik tabanı hakkında geçerli bir bilgiye dayalı tahmin yapma imkânı ortaya koymaktadır. Bir aday gen, fonksiyonel bir gene yakından bağlı olan ve belirli bir özelliğin fonksiyonel fizyolojisi üzerinde biyolojik etkiye sahip olan bir gen olarak tanımlanabilir. Mevcut polimorfizmlerin seçilen aday genlerdeki etkilerini daha iyi anlamak için, bunların nicel özelliklerle olan iliĢkileri Marker Assisted Selection = Marker Destekli Seleksiyon (MAS) programlarında test edilebilmektedir (Wu ve ark., 2005). Bu amaç için son zamanlarda çok sayıda potansiyel aday gen tanımlanmıĢtır. Kantitatif karakterlerde varyasyonun genetik temelini oluĢturan genlerin bireysel etkileri bilinmediğinden, klasik Mendel genetiği metotlarını kullanarak bu genleri tek tek incelemek mümkün değildir. Gen sayısı ve bireysel özellikleri hakkında bilgi eksikliği nedeniyle, teorik çalıĢmalar bazı varsayımlara dayanmaktadır. Temel varsayıma göre, niceliksel kiĢilik, tüm bölgelerde benzer frekanslara sahip çok sayıda geni etkiler ve eklenen etkiler ve baskınlık iliĢkileri neredeyse aynıdır (Roberts ve Smith, 1982; Falconer ve Mackay, 1996; Ağaoğlu, 2010). Bireylere genlerin etkileri doğrudan gözlenemediğinden, çalıĢılan karakterlere iliĢkin genellikle kalıtım derecesi gibi parametreler tanımlanmaktadır. Bireysel anlamda ise genetik yapı veya ele alınan verim özelliği bakımından bireyin sahip olduğu, ama özellikleri bilinmeyen genlerin bütününe iliĢkin bilgiler fenotipik performans ve pedigri bilgilerinden tahminlenmeye çalıĢılmaktadır (Montaldo ve Meza-Herrera, 1998). Kantitatif karakterler boy, kilo, süt verimi, yumurta verimi, canlı ağırlık, döl verim gibi ölçülebilen ve en düĢük değerden en yüksek değere doğru değiĢen, fakat aralarındaki farklılıkların kesin sınırlarla ayrılmadığı normal dağılım eğrisi gösteren karakterlerdir (Hoeschele, 1988; Karaca ve ark., 1992). Bu kantitatif karakterler, çevresel faktörlerden yüksek derecede etkilenmektedirler (Cemal, 1996). Ayrıca, genom taramaları ve son yıllarda moleküler genetik alanındaki moleküler haritaların geliĢtirilmesi üzerine yapılan çalıĢmalarda aktif hale getirilmiĢ olan genetik markerler ile genetik korelasyon çalıĢmaları ile çeĢitli kantitatif karakter özellikleri (Quantitative Trait Loci QTL) keĢfedilmiĢtir (Milan ve ark., 2000). Genetik belirteçlerin getirilmesi, mevcut ana genetik tespit yöntemlerine önemli bir güç katmıĢtır. Bununla birlikte,
planlı melezlemelerle ve DNA analizi için geliĢmiĢ laboratuvar maliyetleri ve bu çalıĢmalar için gereken yüksek masraflı büyük deneysel gruplara olan ihtiyaç, bu çalıĢmaların yaygınlığı bakımından kısıtlayıcı olmaktadır. Çiftlik hayvanlarında verim özelliklerini etkileyen aday genlerin ortaya çıkması, doğal olarak, nicel teoriye yeni bir boyut getirmiĢtir. Bu çerçevede, insanlar, hayvanlar ve bitkilerde major genleri tanımlamak ve genotipleri ayırt etmek için son dönemlerde bazı farklı istatistiksel yöntemler ortaya konulmuĢtur (Elsen ve Le Roy, 1995). En önemli adım, günümüz genetik markör tekniklerini kullanarak MAS sistemi için faydalı genetik markörleri tanımlamaktır (Ihara ve ark., 2004).
Son zamanlarda moleküler genetik teknikler ve istatistik metotlarındaki geliĢmelere paralel olarak populasyonların genotiplerinin DNA seviyelerinde belirlenmesinde kullanılan önemli moleküler markörler : Restriksiyon Fragman Uzunluk Polimorfizmi (RFLP), Mikrosatelitler (Basit Tekrar Dizileri - SSR), ÇoğaltılmıĢ Fragman Uzunluk Polimorfizmi (AFLP) ve Rastgele ÇoğaltılmıĢ Polimorfik DNA (RAPD) ve SNP metotlarıdır. DNA seviyesinde moleküler genetik metotları kullanılarak genotiplere özgü spesifik özellikler ve ekonomik verimler üzerine etkili kantitatif özellik lokuslarının (QTL) incelenmesi söz konusu olabilmektedir. Bu yöntemlerin en önemlilerinden biri olan ve mikrosatellit genom boyunca yinelenen rastgele DNA dizileri için birçok moleküler markör türü tanımlanmıĢtır. MAS amaçları için mikrosatellit markörler incelenmiĢ ve birçok çalıĢmada tanımlanmıĢtır (Chen ve ark., 2006). Diğer önemli ve yaygın kullanım alanına sahip yöntem olan PCR-RFLP, sığırlarda genetik varyasyonu çalıĢmak için ucuz, hızlı ve yararlı bir metot olarak görülmekte ve özellikle önemli genler üzerindeki ya da yakın bölgelerindeki polimorfizm ile ilgili çalıĢmalarda kullanılmaktadır. PCR-RFLP (Polimeraz Zincir Reaksiyonu - Restriction parça uzunluğu polimorfizmi), restriksiyon endonükleazların çift iplikli DNA’yı özel tanıma bölgelerinden kesmesi esasına dayanmaktadır. Hayvanlarda genetik varyasyonu çalıĢmak için ucuz, hızlı ve kullanıĢlı bir yöntem olarak görülür ve polimorfizm çalıĢmalarında kullanılmaktadır.
PCR yöntemi uygulaması, in vitro olarak DNA replikasyon yöntemidir. Reaksiyonlar, farklı sıcaklıklarda üç olayın periyodik olarak tekrar olmasına bağlıdır. DNA fragmanları PCR ile çoğaltılabilir ve çok küçük numunelerden analiz için yeterli miktarlar elde edilebilir (ġekil 1.2).
ġekil 1.2. RFLP analiz tekniği aĢamaları (Aytekin, 2011).
Büyüme, geliĢme ve ekonomik özellikler ile iliĢkili olduğu bilinen birçok aday gen vardır. Genler içerisinde Büyüme Hormonu (BH/ GH), Ġnsülin Benzeri Büyüme faktörleri (ĠBBF/ IGF-1) ve Mipofize Özel Transkripsiyon Faktörü-1(Pit-1) yer almaktadır.
Pit-1 geni seleksiyonda et kalitesi ve verimini geliĢtirebilmek için kullanılabilir. BH'nin büyüme, dokuların büyümesi ve kas geliĢimi ile iliĢkisinin olduğu tespit edilmiĢtir. ĠGF-1 faktörleri geninin ise kas büyümesi, geliĢimi ve fonksiyonu ile iliĢkili olduğu düĢünülmektedir. Pit-1 geni ise özellikle büyüme, geliĢme ve kas büyümesi ile iliĢkilidir. ÇeĢitli ülkelerde yapılan çalıĢmalarda gerek morfolojik, gerekse çeĢitli verim ya da hastalıklarla ilgili özelliklerle iliĢkili olan çeĢitli genler ortaya çıkartılmaktadır. KüçükbaĢ hayvanlarda da et verimi ve kalitesini arttırmaya yönelik çalıĢmalarda et kalitesini doğrudan ya da dolaylı olarak ilgilendiren genler için moleküler markör arama ve geliĢtirme çalıĢmalarının son derece yararlı araĢtırmalar olduğu açıktır. Dolayısıyla, bu gibi moleküler genetik çalıĢmaların et ve özellikle etin kalitesine yönelik bazı özelliklerin ıslahı açısından önemli kazanımlar sağlayacağını da söylemek mümkündür.
Bu çalıĢmada Türkiye yerli keçi ırklarından Kıl, Ankara, Kilis, Honamlı, Halep ve Saanen keçilerinde Pit-1, IGF-1 ve GH-1 genlerindeki polimorfizmlerin PCR-RFLP yöntemi ile belirlenmesi amaçlanmıĢ olup söz konusu ırklarda ilk kez çalıĢılmıĢ olması da önem arz etmektedir.
2. KAYNAK ARAġTIRMASI
2.1. DNA Polimorfizm TeknikleriHayvanların genetik yapısını belirlemek için moleküler genetik yöntemlerin kullanımı gün geçtikçe artmaktadır. Moleküler tanımlama yöntemleri, özellikle genetik kaynakların koruma programlarına yol gösterici rol oyanmakta ve koruma faliyetlerinin etkinliği de bir anlamda test edilebilmektedir. Koruma programlarının temel amacı, mevcut genetik çeĢitliliği, gelecek nesillere en az kayıpla aktarmaktır. Bu amaçla çeĢitli moleküler genetik yöntemler geliĢtirilmiĢ ve kullanılmaktadır. Bu yöntemlerden bazıları; Rastgele ÇoğaltılmıĢ Polimorfik DNA (Random Amplified Polymorphic DNA; RAPD), ÇoğaltılmıĢ Parça Uzunluk Polimorfizmi (Amplified Fragment Length Polymorphism; AFLP), Restriksiyon Parça Uzunluk Polimorfizmi (Restriction Fragment Length Polymorphism; RFLP), Tek Eksen Konformasyon Polimorfizmi (Single Strand Conformation Polymorphism; SSCP), Mikrosatellit veya Kısa Ardarda Tekrarlar (Short Tandem Repeats, STR) veya Basit Dizi Tekrarları (Simple Sequence Repeat; SSR), Dizisi EtiketlenmiĢ Alanlar (Sequence Tag Sequence; STS) ve Tek Nükleotid Polimorfizmleri (Single Nucleotid Polymorphism; SNP) vb. Ģeklindedir. Bu belirteçler ile hem nükleer (çekirdek) genomda hem de mitokondriyal genomda var olan varyasyonun ortaya çıkarılması ile türler, ırklar ve hatta ırk içindeki bireyler arasındaki varyasyonun derecesi araĢtırılabilmektedir (Shivaji ve ark., 2003). Herhangi bir belirteç, genomik bir bölgenin tanımlayıcısıdır ve bu belirteç aracılığı ile tespit edilen alleller bu bölgenin polimorfizmini ortaya koymaktadır. Moleküler belirteçler; DNA düzeyinde polimorfizmlerin ortaya konmasında ve özellikle hayvan genetiği çalıĢmalarında yaygın bir Ģekilde kullanılmaktadır (Fatima, 2006). Bu belirteçlerden biri olan RFLP (Restriksiyon Parça Uzunluk Polimorfizmi) tekniğinin uygulanması kolay bir yöntem olsa da, çalıĢılan gen bölgesi için hangi enzimin uygun olduğunu belirlemek çok önemli bir basamaktır. Enzim, çalıĢılan alan için uygun olmadığında, çok fazla veya daha yakın bant görülebilir. Ayrıca bu durumda, büyüklükleri arasında kıyaslama ve bantların sayısını karĢılaĢtırmak ve değerlendirmek mümkün olmayabilir. Bu amaçla, incelenen organizmaya göre elde edilen PCR ürünlerini kesen kısıtlama enzimlerini ve kesim enzimleri değerlendirmek için kullanılan birçok program vardır. Böylece, seçilen organizmanın hedef gen bölgesinde kullanılacak olan kesim enziminin seçimindeki hata azaltılabilmektir. Test için pozitif ve negatif kontroller kullanılmalıdır. RFLP tekniği uygulanmasından sonra, elde edilen ürünler, agaroz jel üzerine yüklenir ve etidyum bromid ile boyanıp jel elektroforezinde yürütüldükten sonra UV ıĢığı altında görüntülenebilmektedir.
RFLP metodu, restriksiyon endonükleazların keĢfedilmesinden sonra 1960'larda geliĢtirilmiĢtir. RFLP markörleri ilk baĢarılı genetik haritalama çalıĢmalarında kullanılmıĢtır. DNA'daki mutasyonları veya polimorfizmleri saptama ve kesim enzimlerinden kaynaklanan değiĢken uzunluktaki DNA segmentlerini belirleme yöntemidir. PCR’ın ardından bazı restriksiyon enzimlerinin kullanılmasıyla mutasyon taraması yapılabilmektedir. Restriksiyon enzimleri, DNA’da özgün dizileri tanır ve o noktadaki fosfodiester bağını keserler. Bu farklı DNA profillerine ekleme, silme, nükleotid değiĢimi veya tek nükleotid polimorfizmi (SNP) neden olabilir (Young ve ark., 1992). RFLP metodu populasyonlarda, genetik çeĢitlilik, genetik kompozisyon, gen haritalama ve iliĢki analizleri gibi çalıĢmalarda kullanılmaktadır.
2.2.Türkiye Yerli Keçi Irkları ve Önemi.
Türkiye’de 2019 yılı geçici istatistiklerine göre 10 992 427 milyon baĢ keçi bulunmaktadır. Toplam keçi varlığının yaklaĢık %98’ini Kıl keçi ve melezleri, %2’sini ise Tiftik keçileri oluĢturmaktadır (Anonim, 2018). Koyun ve keçi yetiĢtiriciliğinin tarım ekonomisi ve insanın kırsal yaĢamı dıĢında genetik çeĢitlilik açısından, özel bir önemi vardır. Keçi yetiĢtiriciliği, özellikle kırsal alanlarda için yaĢayanlar hayvansal protein kaynağı olarak önemli bir yere sahiptir. Keçi; et, süt, tiftik, kıl, deri, post ya da gübre gibi verimlerinden yaralanılan evcil hayvan türlerinden biridir (Akçapınar, 1994). Az masrafla yetiĢtirilebilmesi, diğer çiftlik hayvanlarına göre bakım ve elveriĢsiz beslenme koĢullarına daha karĢı dayanıklı olması nedeniyle geliĢmekte olan ülkelerde hayvancılık sektöründe önemli bir yer tutmaktadır (ġengonca, 1989; ġimĢek, 2006). Türkiye hayvancılığı içerisinde keçi yetiĢtiriciliği önemi giderek artan bir yere sahiptir. Türkiye’de keçi ırkları Kıl Keçisi, Ankara Keçisi, Kilis Keçisi, Honamlı Keçisi, Norduz Keçisi gibi çeĢitli ırklardan oluĢmaktadır.
2.2.1.Kıl Keçisi
Türkiye yerli kıl keçileri uzun sürüden beri yetiĢtirildiği Anadolu'nun iklim; çevre ve yetiĢtirme koĢullarına dayanıklı, zayıf meralardan yararlanabilen kanaatkar hayvanlardır. Kıl keçilerinde vücut rengi beyazdan siyaha kadar değiĢebilmekle birlikte en çok rastlanan renk siyahtır. YetiĢtirildikleri bölgeye göre uygulanan bakım ve besleme yöntemleri farklılık gösterebilmektedir. Kombine, süt ve et verim yönlü olarak yetiĢtirilmektedir. Türkiye'nin bütün bölgelerinde bulunduğu gibi komĢu Arap ülkelerinde, Ġran ve Afganistanda da yetiĢtirildiği bildirilmiĢtir (Akçapınar, 1994). Genellikle boynuzlu olmakla birlikte boynuzsuzlarına da rastlanmaktadır, zayıf fakat güçlü bir vücut yapısına sahip olan kıl keçilerinin yürüme ve tırmanma yetenekleri çok iyidir. (ġekil 2.1). EriĢkin erkekler ortalama
65-90 kg, diĢiler ortalama 45-65 kg canlı ağırlığa sahiptirler. Genel olarak verimlerinin düĢük olduğu kabul edilmiĢtir. Ancak, yetersiz bakım-besleme, parazit ve hastalıklara dayanıklı zorlu iklim koĢullarına uyabilen bir ırktır. Kıl keçisinde süt verimi 80- 150 litre, laktasyon süresi 180-220 gündür. Doğumda keçi baĢına oğlak sayısı 1'dir. Kıl keçi eti yetiĢtirildikleri bölgelerde sevilerek tüketilir. Dünya piyasalarındaki keçi eti talebi ve fiyatı göz önünde bulundurulduğunda önemli bir ihraç ürünü ve gelir kaynağı olabileceği anlaĢılan keçi eti ve et verimi üzerinde önemle durulması gerektiği görülür. Türkiye'de keçi varlığının yaklaĢık %98’ini Kıl keçi ve melezleri oluĢturmaktadır (Anonim, 2018).
ġekil 2.1. Konya Doğu sınırdaki Kıl keçilerine ait fotoğraflar.
2.2.2. Ankara Keçisi (Tiftik)
Ankara Keçisi adını Ankara ilinden alan ve dünyada Angora olarak bilinen önemli bir yerli keçi ırkıdır. BaĢlıca verimi tiftiktir ve tiftik dünyada Mohair olarak bilinen önemli bir dokuma maddesidir. Bütün dünyada Ankara Keçisi olarak tanınan bu ırk 1838 yılına kadar sadece Anadolu'da yetiĢtirilmiĢ, bu tarihten sonra öncelikle Güney Afrika olmak üzere dünyanın çeĢitli ülkelerinde yetiĢtirilmeye baĢlanmıĢtır. Türkiye'de Ankara keçisi yetiĢtiriciliği özellikle Orta Anadolu'da yoğunlaĢmıĢtır. Bunun dıĢında, Siirt, Mardin, Bitlis, Sivas, Burdur, Isparta illerinde de yetiĢtiriciliği yapılmaktadır. Ankara keçisi küçük ve zayıf yapılıĢlı bir ırktır. Keçiler 28-35, tekeler 35-45 kg canlı ağırlıktadır. Temel vücut rengi beyaz olan Ankara keçilerinin, krem, sarı, gri, kahverengi, siyah ve alacalı renklerine de rastlanmaktadır (Anonim, 2009). Ankara keçilerinde esas verim tiftiktir. Tiftik verimi yaĢ, cinsiyet, beslenme ve bireyin özelliklerine bağlı olarak değiĢmek üzere diĢilerde 1.5-3 erkeklerde 3-6 kg arasındadır (ġekil 2.2). Ankara keçisi yetiĢtiriciliğinin bu durumu yetiĢtiricilerin gelirlerinin arttırılması için tiftik dıĢındaki verimlerininde üzerinde önemle durulması gerektiğini göstermiĢtir. Ankara keçilerinin ıslahında tiftik verim ve özellikleri yanında süt ve et verim özelliklerinin de üzerinde durularak daha üretken bir tipin oluĢturulmasına çalıĢılmalıdır
(Anonim, 2009). Ankara keçisi etinin kokusuz oluĢu sevilerek tüketilmesi ve tüketiminin yaygınlaĢması bakımından avantaj olarak kabul edilmektedir (Ağaoğlu, 2010). Oğlak ve çepiçlerinin etleri gevrek ve lezzetlidir. Buna karĢılık ikiz doğum oranı düĢük olan ve nisbeten geç geliĢen bir ırktır (Soysal ve ark., 2004).
ġekil 2.2. Ankara Keçilerine (Tiftik Keçisi) ait fotoğraflar
2.2.3. Kilis Keçisi
Kilis Keçisi Kilis, Gaziantep, Adıyaman ve Hatay illerinde yayılma alanı göstermektedir. bu ırk Damascus (ġam) keçileri ile kıl keçilerinin melezlenmesi ile ortaya çıkmıĢtır (Anonim, 2009). Genellikle küçük gruplar halinde bağ, bahçe çevresinde aile iĢletmelerinde yetiĢtirilir. Vücut uzun siyah kıllarla örtülüdür. Gri, kahverengi ve alacalara da rastlanır. Kulaklar tipik olarak uzun, geniĢ ve sarkıktır (ġekil 2.3). Gögüs ve sağrı bölgeleri iyi görünüme sahiptirler. Canlı ağırlık erkeklerde 50-60 kg, diĢilerde 35-45 kg'dir. 210-260 günlük laktasyon sürelerinde 200-300 kg süt verirler (Ġriadam, 2004). Ġyi bakım ve besleme koĢullarında süt verimleri 400-500 kg'a kadar çıkabilir. Doğuran keçi baĢına ortalama 1.5 oğlak verimine sahiptir. Sayıları 100 bin dolayında olan Kilis keçileri Türkiye yerli keçilerin içerisinde süt verimi en yüksek olan ırktır (Ertuğrul, 2007). Boynuzlu ve boynuzsuz olanları mevcuttur. Erkekleri kalın, kuvvetli ve uzun, diĢileri kısa, ince ve geriye kıvrık boynuzludur. Meme çok iyi geliĢmiĢ, sarkık meme tipindedir. Kilis keçilerinin et verimi tatminkardır. Kilis keçisi dayanıklı yerli bir keçi ırkıdır. Kilis keçisi yetiĢtiriliğine yönelik projeler, Kilis keçilerinin banka kredisine dahil edilmesi sonucu hızlanmıĢtır. Sütçü Kilis keçilerinin Güney Doğu Anadolu ve Doğu Akdeniz Bölgesinde yaygın olarak yetiĢtiriciliği yapılmaktadır.
ġekil 2.3. Kilis Keçilerine ait fotoğraflar (Anonim, 2009).
2.2.4. Honamlı Keçisi
Akdeniz Bölgesi, Toros Dağları etekleri, Antalya, Isparta ve Konya yörelerinde yetiĢtirilen ve dıĢbükey bir burun yapısı ve kısa üst dudak yapısı ile dikkat çeken bir ırktır (Elmaz ve ark., 2012). Renk çoğunlukla siyahtır. Alın ve ayaklarda beyaz lekelere rastlanmaktadır. Keçilerde canlı ağırlık erkek 97 ve diĢi 72 kg, cidago yüksekliği erkeği 91 ve diĢiği 85 cm, Vücut uzunluğu erkeği 93 ve diĢiği 84 cm, laktasyon süresi 270 gün ve pazarlanabilir süt verimi 175 litredir. Ayrıca kuyruk yapıları da Kıl keçilerinden daha uzun ve püskül görünümüne sahiptir. Deri koyu renklidir. Erkek ve diĢiler genellikle boynuzludur (Anonim, 2009). Alt çene üst çeneden uzundur. Gözleri belirgin bir Ģekilde iri ve canlıdır. Kulaklar küçük ve kalındır. Burun belirgin bir Ģekilde dıĢ bükeydir. Uysal ve insana çok yakın bir ırktır (ġekil 2.4). GeniĢ ölçüde, Akdeniz bölgesinde yörükler tarafından, bölgelerinde bulunan fundalıklar, çalı formundaki bitkiler, orman içi meralar, anızlar ve nadasa bırakılmıĢ alanlardaki otlakrlarda neredeyse masrafsız bir Ģekilde yetiĢtirilir (Ağaoğlu, 2010). Yılın hemen tamamında merada barındırılabilen bir ırktır. Bununla birlikte ağız yapılarından dolayı zayıf meralardan yeteri kadar yararlanamaz. Zengin mera veya ek yemleme olanağına sahip iĢletmelerde baĢarıyla yetiĢtirilmektedir. Göçebe yörükler keçi kılından yapılan çadırlarda yaĢarlar. Son yıllarda bilimsel olarak tanımlanmaya baĢlanan Honamlı keçileri yok olma tehdid altındadır. Honamlı keçisinin tanımlanması ve korunması ile ilgili projeler yürütülmektedir (Anonim, 2009).
ġekil 2.4. Honamlı Keçilerine ait fotoğraflar (Anonim, 2015).
2.2.5. Saanen Keçisi
Saanen Keçisi 1959 yılı baĢlarında Türkiye’ye getirilmiĢ ve halen saf ya da melez olarak yetiĢtirilmektedir (ġekil 2.5). Marmara ve Ege sahillerinde yaygın olarak bulunmaktadır. Saanen ırkının en önemli özelliği farklı iklim koĢullarına uyum gösterme yeteneği sayesinde, götürüldüğü her yere çok çabuk adapte olabilmesidir. Saanen keçileri yemleme ve mera koĢullarına karĢı çok duyarlıdır. Yüksek verim yeteneği ancak iyi bakım ve besleme koĢullarında ortaya çıkar (Özcan, 1989); (Özder, 2006). Saf Saanen keçileri, bölgede bulunan Maltız (malta keçisi: ada keçisi) ve kıl keçilerinin melezlenmesi ile oluĢturulmuĢtur. Türk Saanen keçisi süt beyazı renkten krem beyaza kadar değiĢen tonlarda beyaz rengin hâkim olduğu bir genotiptir. Kıl uzunluğu oldukça kısa ve parlaktır. Deri esnek ve yumuĢak olup sütçü bir yapıyı temsil eden niteliktedir. Vücut narin, diĢilik özelliklerini gösterir tipte, boyun ince ve uzun, baĢ narin ve göğüs kafesi geniĢ ve sağlam yapıdadır. Sağrı eğimi çok düĢük olmayıp hafif eğimdedir (Anonim, 2014).
Türk Saanen keçilerinin süt verimleri ortalama 500 kg/laktasyon düzeyinde olup, bakım Ģartlarının iyileĢtirilmesi ile aynı sürüde 800 kg’a kadar süt üreten hayvanlara rastlanabilmektedir. Süt keçileri 200-300 gün sağılabilmektedir (Özder, 2006; Haris, 1996). Ayrıca bu ırk yerli keçi ırklarına göre daha az hareketli olup entansif yetiĢtirme koĢullarına daha uyumludur. Süt ve döl verimi yüksek bir ırktır (Kaymakcı ve ark., 2006).
2.2.6. Halep Keçisi
Halep Keçisi, Suriye, Arap ülkeleri ve Türkiye'de yetiĢtirilir. Damascus yada ġam keçisi olarak bilinen kahverengi veya kırmızı renkte sütçü bir varyetesinin yanında Filistin keçisi olarak bilinen baĢka bir varyetesi vadır (ġekil 2.6). Gaziantep, Kilis, ġanlıurfa, Adıyaman ve Hatay bölgesinde görülmektedir. Kombine, süt ve et verim yönlü bir ırktır. Halep keçileri doğum baĢına 2-3 yavru verir. Canlı ağırlık diĢilerde 50-60 kg ve Erkeklerde 90-130 kg arasında değiĢmektedir.
ġekil 2.6. Halep Keçisi (Anonim, 2015).
2.3. Pit-1 geni polimorfizmi
Hipofiz transkripsiyon faktörü-1 geni (POU1F1; Pit-1) büyüme ve geliĢmenin düzenlenmesinde memelilerde aday gen olarak kabul edilmektedir. Pit-1 geni POU familyası (Herr ve ark., 1988) POU domain class 1 transcription factor 1) olarak isimlendirilmekte ve memelilerde büyüme hormonu (GH), prolaktin (PRL) ve tirotropinin (TSHβ) ekspresyonundan sorumlu olan bir gendir (Haugen ve ark., 1993; Renaville ve ark., 1997; Kopp ve Jameson, 1998; Miyai ve ark., 2005). Keçi kromozomlarında 1q21- 22’de lokalize olup, POU ve homeo-alan bölgeleri içeren 6 ekzondan meydana gelmektedir (Woollard ve ark., 2000). Bu genin bağlanma yeri (ġekil 2.7), ekzon 1'deki transkripsiyonun varsayılan baĢlangıcından 350 kb aĢağıdadır (Howard ve ark., 2009). Bu gende meydana gelen bazı varyosyonlar memelilerde GH, PRL ve TSH hormonlarının düzenini bozmakta ve ayrıca hipofizer geliĢmede anormalliklere (hipoplazi) de neden olabildiği bildirilmektedir (Renaville ve ark., 1997).
ġekil 2.7. Keçi POU1F1 geninin yapısı ve ekzon 6 ve Pst1 enzim bölgesi için SNP'lerin konumu Ģekil 2.7'de verilmiĢtir (Lan ve ark., 2007a).
Lan ve ark. (2007b)’nın Ġç Moğolistanda, yaptıkları çalıĢmada, toplam 801 baĢ, Beyaz KeĢmir (452 baĢ), Xinong saanen (74 baĢ), Laoshan (80 baĢ), Guanzhong (62 baĢ), Guizhou Siyah (21 baĢ), Matou (22 baĢ), Banjiao (25 baĢ), Guizhou Beyaz (31 baĢ) ve Leizhou keçisi (34 baĢ), keçi ırklarında POU1F1 genindeki: TCT (241Ser)>TCG (241Ser) sessiz bir alleli tespit etmek için bir DdeI PCR-RFLP yöntemini kullanmıĢlardır. D1 allelinin frekansları 0.600 ila 1.000 arasında değiĢtiğini bildirmiĢlerdir. Süt veriminde DdeI polimorfizminin üretim özellikleri ile anlamlı iliĢkileri bulunmuĢtur (P<0.05). Farklı genotipler arasında doğum tipi ve bir yaĢ ağırlığı bakımndan gözlemle farkılıklar önemli bulunmuĢtur (P<0.05). D1D1 genetipli bireyler D1D2 üstün süt verim performansı, üreme özelliği ve ağırlık özelliği için potansiyel aday gen olarak önerilmektedir. ÇalıĢmada D1D1 genotipli keçiler seleksiyonda daha iyi performans için yetiĢtiricilere moleküler bir markör olarak tavsiye edilmiĢtir.
Lan ve ark. (2009)’nın yaptıkları çalıĢmada, Ġç Moğolistan'da 847 baĢ Beyaz KeĢmir keçisinde, genotipleri belirlemek için PCR-RFLP yöntemi ile polimorfizmi araĢtırmıĢdır. PstI restiriksiyon enzimi ile yaptıkları çalıĢmada, T ve C allel frekanslarını sırası ile 0.959 ve 0.041 olarak bulmuĢlar ve populasyonun Hardy-Weinberg dengesinde (P>0.05) olduğunu bildirmiĢlerdir. Ayrıca, POU1F1 geninin PstI polimorfizmi ve keĢmir verimleri arasındaki iliĢkiler önemli bulunmuĢtur (P<0.05).
Alakilli ve ark. (2012) yaptıkları çalıĢmada; Pit-1 geninin, ekzon 6, polimorfizmini Restriksiyon Fragman Uzunluk Polimorfizmi (RFLP) yöntemi ile araĢtırmıĢlardır. Pst1 kesim enzim ile kesme iĢini yapmıĢlardır. Sonuç olarak Barki, Zaribi, Ardi ve Masri ırklarında T allel frekanslarının 0.250 ve 0.840 arasında değiĢtiğini bulmuĢlardır.
Sharma ve ark. (2013), yaptıkları çalıĢmalarda; toplam 50 baĢ Barbari ırk keçilerde PCR-RFLP yöntemi ile Pit-1 gen polimorfizmi çalıĢmıĢlardır. PstI restiriksiyon enzimi ile yaptıkları çalıĢmada, PstI kesim bölgelerine dayanan iki allel tespit etmiĢler ve TT
genotiplerinin frekanslarını 1.0, TC ve CC genotiplerinin frekanslarını 0 bulmuĢlardır. T ve C allel frekanslarını sırası ile 1.0 ve 0 olarak bildirmiĢlerdir. Sonuç olarak, Barbari keçi ırkının Pit-1 geni bakımından monomorfik olduğunu ifade etmiĢlerdir.
Dagong ve ark. (2016), 72 baĢ, Marica, Kacang ve Peranakan Ettawa ırkı keçilerde PCR-RFLP yöntemi ile Pit-1 gen polimorfizmi çalıĢmıĢlardır. PstI restiriksiyon enzimi ile yaptıkları çalıĢmada, PstI kesim bölgelerine dayanan iki allel tespit etmiĢ, T ve C allel frekanslarını sırası ile 0.76 ve 0.24 olarak bildirmiĢlerdir. Sonuç olarak, bulunan varyasyonun, Endonezya yerli keçileri Pit-1 polimorfizmlerinin doğal büyüme ve karkas kalitesine olan iliĢkisini bulmak için gelecekteki araĢtırmalarda kullanılabileceğini ifade etmiĢlerdir.
Li ve ark. (2016) Çinde yetiĢtirilen yerli 6 ırktan oluĢan toplam 709 baĢ keçi üzerinde Nanjiang kaĢmir keçi (247 baĢ), Sincan keçi (175 baĢ), Shaanbei Beyaz keĢmir keçi (150 baĢ), Boer keçi (84 baĢ), Haimen keçi (33 baĢ) ve Xuhui keçi (20 baĢ) Pit-1 geni polimorfizmini çalıĢmıĢlardır. Bu çalıĢma POU1F1-DdeI lokusundaki keçi polimorfizmlerini ve 15 yerli Çin keçi ırkında allel ve genotip frekanslarını PCR-RFLP yöntemi ile araĢtırmıĢlardır. DdeI enzim kesimi sonucunda D1 ve D2 allel frekanslarını olaral, 0.790 ve 0.210 D1D1, D1D2 ve D2D2 genotip frekanslarını ise 0.550, 0,371 ve 0,079 olarak bulmuĢtur. Bu lokustaki genotip ve allel frekansların dağılımları Ki kare testine dayanan populasyonlar arasında anlamlı olarak farklı bulunmuĢtur (P<0.001). POU1F1 geninin, moleküler genetik karakterini ırk faktörünün önemli derecede etkilediği belirlenmiĢtir.
Sarma (2019) çalıĢmasında Assam Hill keçisinde FecB ve POU1F1 geninin polimorfizmini araĢtırmıĢtır. “Keçi ĠyileĢtirme Üzerine AICRP” altında Batabari, Nahira ve Tetelia gibi üç biriminde muhafaza edilen tek ve çoklu doğum tipine sahip 80 rastgele seçilen Assam Hill keçisine ait kan örnekleri kullanılmıĢtır. 190 bp'lik bir FecB geni fragmanı ve 450bp'lik bir POU1F1 geni fragmanı, kullanılarak çoğaltılmıĢtır. AvaII enzimi kullanılarak FecB'nin RFLP analizi, tüm numuneler için kesilmemiĢ 190 bp ürünü ortaya çıkarmıĢtır. POU1F1 geni DdeI ile enzimi kesilmiĢtir. Monomorfizmi açığa çıkaran tüm numuneler için agaroz jel elektroforezinde 102 bp, 118 bp ve 200 bp'lik üç fragman üretmiĢtir. POU1F1 geninin RFLP analizi de, kesilmemiĢ 450 bp'lik ürünle sonuçlanan PstI kullanılarak yapılmıĢtır. Sekans analizinde, FecB geninde AvaII için hiçbir sınırlama bölgesi bulunmamakla birlikte POU1F1 geninde DdeI için dört kısıtlama bölgesi bulunmuĢtur. Numunelerin sekans analizi, tüm numuneler arasında doğum tipi bakılmaksızın her iki gen için % 99-100 homegenlik göstermiĢtir. Pit-1 geni ile bazı çalıĢmaların özeti aĢağıda Çizelge 2.2’de özetlenmiĢtir.
Çizelge 2.2. Hipofiz Spesifik Transkripsiyon Faktörü-1 (Pit-1) geni ile ilgili bazı çalıĢmalar Kaynak ve PCR ürünü Irk N Ekzo n bölge
Genotip frekansları Allel
frekansları Verimle iliskisi
TT TC CC T C Lan ve ark. (2009) 450 bç ĠMWC 847 Ekzon 6 0.917 0.083 0.00 0.959 0.041
T ve C allelleri için allelik frekanslar sırasıyla 0.959 ve 0.041'dir ve genotipik frekanslar Hardy-Weinberg dengesindedir (P>0.05). Ayrıca POU1F1 geninin
PstI polimorfizmi ile keçi
kaĢmir verimi arasında iliĢkiler bulunmuĢtur (*P<0.05). Alakilli ve ark. (2012), 450 bç Barki Zaribi Ardi Masri 20 20 15 15 Ekzon 6 - - - 0.840 0.351 0.512 0.250 0.160 0.649 0.488 0.650 # Sharma ve ark. (2013) 450 bç Barbari 50 Ekzon 6 0.917 0.083 0.00 - - # Dagong ve ark. (2016), 450 bç Kacang PE Marica 21 31 20 Ekzon 6 0.43 0.36 0.90 0.57 0.64 0.10 0 0 0 0.71 0.67 0.95 0.29 0.33 0.05 #
# herhangi bir verimle iliĢkilendirilmemiĢtir.
2.4. IGF-1 geni polimorfizmi
Ġnsülin Benzeri Büyüme Faktörleri (IGF-1 ve IGF-2) ile insülin canlılarda büyüme ve geliĢmenin düzenlenmesinde kritik bir role sahiptir (Reinecke ve Collet, 1998). Dolayısıyla IGF genindeki polimorfizmler araĢtırıcıları bu genin aday gen olarak kullanılması için farklı çalıĢmalara yöneltmiĢtir. IGF'ler, IGF-I ve IGF-II olarak iki önemli forma sahiptir. IGF-II intrauterin büyüme üzerinde etkili iken IGF-I postnatal büyüme üzerinde daha etkilidir. IGF bağlayıcı proteinler (IGFBP) altı yüksek afiniteli protein ailesinin bir üyesidir. Bağlanma proteinleri, hücre zarında, insülin alıcısında üç farklı reseptöre sahiptir. IGF'ler kemik ve kas üzerindeki otokrin ve parakrin mekanizmalarından etkilenmiĢtir. GeliĢme, büyüme, üreme ve yaĢlanmada önemli bir rol oynamaktadır (Miller ve Gore, 2001; Lan ve ark., 2007c; Li ve ark., 2009). IGF-I geni, kromozom 5 üzerinde bulunan tek bir gen tarafından kodlanır (Schibler ve ark., 1998). Bu gen üç lider ekzon (1 w, 1 ve 2) ve dört çeĢit olgun mRNA
kodllar (Mikawa ve ark., 1995). IGF-I memeli büyümesinde, laktasyonda ve metabolizmada kilit rol oynar (Zhang ve ark., 2008). Kas hücreleri için özel proteinler sentezletir ve bu Ģekilde kasın miyofibril alanını arttırarak büyümeyi sağlar. Kemik formasyonu açısından da önemli bir gen olup iskelet geliĢimini doğrudan veya dolaylı mekanizmalarla etkilemektedir. POUIF1 (ayrıca Pit-1 veya GHF-1 olarak adlandırılır) esas olarak memelilerde hipofizde eksprese edilir. Büyüme hormonu (BH), prolaktin (PRL) ve tiroid stimüle edici hormon β (TSH-β) sentezinde rol oynar. Bu gen üzerinde mutasyon muhtemelen GH, PRL ve TSH eksikliği ile sonuçlanabilir (Cohen ve ark., 1996; Li ve Crenshaw, 1990 ). Keçiler için Ġnsülin Benzeri Büyüme Faktörleri (IGF) geninin bütün sekansına GenBank veri tabanında, 80190 bç ve NC_030812 eriĢim numarası ile ulaĢılmaktadır Ģekil 2.8'de ĠGF-1 geni; (Anonymous, 2017-a).
ġekil 2.8. Keçi IGF-1 geni, 5. kromozom genetik haritası (Schibler ve ark., 1998)
Wu-Jun ve ark. (2010)’nın yaptıkları çalıĢmada, 530 baĢ Çin'den iki Xinjiang yerli ırk keçilerinde, ekzon 4, intron 4, PCR-RFLP yöntemi ile IGF-1-P1 gen polimorfizmi çalıĢmıĢlardır. HaeIII restiriksiyon enzimi ile yaptıkları çalıĢmada Xinjiang ve Nanjiang keĢmir keçilerinde AA genotipleri için frekansları sırası ile 0.487 ve 0.277 olarak; BB genotipleri için ise 0.274 ve 0.486 olarak bulmuĢlardır. AB genotiplerin frekansları ise sırası ile 0.239 ve 0.236 Ģeklindedir. IGF-1 geninin polimorfizmlerinin KeĢmir verimi, lif çapı ve canlı ağırlık bakımından önemsiz olduğu bildirilmektedir (P>0.05).
Deng ve ark. (2010) Çin’de yetiĢtirilen iki sütçü keçi ırkından toplam 708 baĢ hayvanda insülin Benzeri Büyüme Faktörü-1 (IGF-1) gen polimorfizmlerini ve bu polimorfizmlerin süt verimi ve canlı ağırlık ile iliĢkisini araĢtırmıĢlardır. IGF-I geni 5’-flanking bölgesinde ve 4. intronda sırasıyla guaninden adenine (g.1617 G-A) ve guaninden sitozine (g. 5752 G-C) iki
yeni mutasyon bildirmiĢlerdir. g.1617 G>A mutasyonunun süt verimi ve canlı ağırlık ile önemli bir iliĢkisinin bulunmadığı bildirilmiĢtir (P>0.05). Ayrıca, CG genotipli Xinong Saanen sütçü keçilerinin vücut ölçülerinin daha yüksek olduğunu bildirmiĢlerdir (P<0.05). GG genotipli Guanzhong keçilerinin göğüs çevrelerinin daha fazla olduğu tespit edilmiĢtir (P<0.05). CC genotipli Xinong Saanen sütçü keçilerinin birinci ve ikinci laktasyon süresince süt verimlerinin daha yüksek olduğu bildirilmiĢtir (P<0.05). Bu nedenle, g. 5752 G>C mutasyonunun Çin sütçü keçi yetiĢtiriciliği ve genetiği için genetik belirteç olarak kullanılabileceği bildirilmiĢtir.
Qiong ve ark. (2011), Çin’de yetiĢtirilen üç yerli keçi ırkında, toplamda 776 baĢ, Ġnsülin Benzeri Büyüme Faktörü-1 (IGF-1) geninin 4. ekzonunda bulunan, mutasyonların KeĢmir verim özellikleri ve canlı ağırlık arasındaki iliĢkiyi incelemiĢlerdir. Xijiang, Bogeda KeĢmir ve Nanjiang KeĢmir keçi ırkları için sırasıyla AA genotipleri için frekansları sırası ile 0.414, 0.591 ve 0.319; AB genotipleri için frekansları sırası ile 0.000, 0.126 ve 0.029; BB genotipleri için frekansları sırası ile 0.586, 0.241 ve 0.597; AC genotipleri için frekansları sırası ile 0.000, 0.042 ve 0.055 olarak bulunmuĢtur. ÇalıĢma da ile 4. ekzonda yeni bir SNP’nin (c. 1617 G>A, c. 1620 C>T) varlığı ortaya çıkarılmıĢ ve bunların sessiz mutasyon olduğunun tespit edildiği bildirilmiĢtir. Nanjiang KeĢmir keçi populasyonunda; AA genotipindeki keçilerde mutasyonun AB genotipine göre daha düĢük olduğu bildirilmiĢtir (P<0.05). AC genotipindeki bireylerin canlı ağırlığının BB genotipinden daha fazla ve bunun önemli olduğu bildirilmiĢtir (P<0.05).
Alakilli ve ark. (2012)’nın yaptıkları çalıĢmada Ġnsülin Benzeri Büyüme Faktörleri (IGF-1) geninin 4. ekzondaki polimorfizm ile keçilerde büyüme özellikleriyle iliĢkisi araĢtırılmıĢtır. 4 Mısır ve Suudi keçi ırklarında (Barki, Zaribi, Ardi ve Masri) PCR-RFLP yöntemini kullanmıĢlar ve BH, IGF-I, POUIF1, MSTN ve BMP15 genlerinin genotip ve allel frekanslarını tespit etmiĢlerdir. Ġnsülin Benzeri Büyüme Faktörü-1 (IGF-1) geni için A ve B allel frekanslarını Barki keçi ırkında 0.731 ve 0.269, Zaribi keçi ırkında 0.432 ve 0.568, Ardi keçi ırkında 0.615 ve 0.385, Masri keçi ırkında 0.473 ve 0.527 olarak tespit edilmiĢlerdir.
Liu ve ark. (2012), 316-bp'lik bir parçası Ġnsülin Benzeri Büyüme Faktörleri-3 (IGFBP-3)gene 2. Ekzon, üzerindeki polimorfizmi PCR-RFLP yöntemi ile araĢtırılmıĢtır. ÇalıĢmada HaeIII kesim enzim kullanarak 3 genotip (AA, AB ve BB) gözlemlenmiĢtir. Genotiplerin frekansları sırayla; 0.58, 0.33 ve 0.09. A ve B allel frekanslarını ise 0.75 ve 0.25 olarak bildirmiĢlerdir. Nükleotit sekanslaması ekzon 2 bölgesinde C>G polimorfizme neden olan R158G değiĢikliği göstermiĢtir. En küçük kareler analizi genotiplerin hayvanların keĢmir
ağırlığı (P<0.001), keĢmir lifi uzunluğu (P<0.001) ve üst kaba kıl uzunluğu (P<0.05) üzerinde önemli bir etkisinin olduğunu göstermiĢtir. AB ve BB genotipli hayvanların, AA genotipine sahip hayvanlara göre daha yüksek keĢmir ağırlığı, keĢmir elyaf uzunluğu ve kıl uzunluğuna sahip oldukları gözlenmiĢtir. Bu sonuçlar, IGFBP-3 genindeki polimorfizmlerin keĢmir keçilerinde keĢmir ağırlığı için potansiyel bir moleküler markör olabileceğini göstermiĢtir.
Sharma ve ark. (2013), yaptıkları çalıĢmada, Hindistan'da yetiĢtirilen keçilerin üç büyük boy ırk (Jamunapari, Beetal ve Jakhrana), üç orta boy ırk (Sirohi, Barbari, ve Osmanabadi) ve üç küçük cüsseli (Black Bengal, Changthangi, ve Gaddi) olmak üzere farklı ırklara ait toplam da 80 baĢ keçi kullanmıĢlardır. ÇalıĢmalarında Ġnsülin Benzeri Büyüme Faktörü-1 (IGF-1) geni SNP’lerini araĢtırmıĢlardır. KarĢılaĢtırmalı dizi analizi sonucu toplamda 18 SNP tespit edilmiĢtir. Gözlenen ortalama heterozigotluk Sirohi, Beetal, Osmanabadi ve Gaddi keçi ırklarında maksimum (0.074), sıyah Bengal keçi ırkında minimum (0.019) bulunmuĢtur. Barbari ve Gaddi'nin daha farklı olduğu tespit edilmiĢ (0.0123), Changthangi ve Jamunapari, Nei'nin genetik mesafesine bağlı olarak en az farklılaĢmıĢlardır (0.00110).
Zeytünlü (2015) çalıĢmasında, Honamlı (n=150) ve Kıl (n=150) keçi ırklarına ait toplam da 300 baĢ keçi kullanmıştır. Ġnsülin Benzeri Büyüme Faktörü-1 (IGF-1) geni HaeIII (g. 5752 G-C) polimorfizminin belirlenmesi, polimeraz zincir reaksiyonu ve restriksiyon parça uzunluk polimorfizmi PCR-RFLP yöntemi ile araĢtırmıĢlardır. Kıl keçisi ırkında 2 allel (G ve C), 3 genotip (GG, GC ve CC) ve Honamlı keçisi ırkında 2 allel (G ve C), 2 genotip (GG, GC) tespit edilmiĢtir. IGF-1 geni için GG genotipi Honamlı ve Kıl keçi ırklarının en yaygın genotipi olarak tespit edilmiĢtir. Honamlı ve Kıl Keçisi ırklarının Hardy-Weinberg dengesinde olduğu gözlenmiĢtir (P>0.05).
Sharma ve ark. (2013), yaptıkları çalıĢmada, Hindistan'da yetiĢtirilen keçilerin üç büyük cüsseli (Jamunapari, Beetal ve Jakhrana), üç orta cüsseli (Sirohi, Barbari, ve Osmanabadi) ve enzeri Büyüme Faktörü-1 (IGF-1) geni HaeIII (g. 5752 G-C) polimorfizminin belirlenmesi, PCR-RFLP yöntemi ile araĢtırılmıĢtır. IGF-I geni HaeIII polimorfizmi yönünden belirlenen genotipler incelendiğinde; Kıl keçisi ırkında 2 allel (G ve C), 3 genotip (GG, GC ve CC) ve Honamlı keçisi ırkında 2 allel (G ve C), 2 genotip (GG, GC) tespit edilmiĢtir. Genotip frekansları ile doğru orantılı olarak G allelinin her iki ırkta da C alleline göre daha yüksek frekansta olduğu belirlenmiĢtir. IGF-I geni için GG genotipi Honamlı ve Kıl keçi ırklarının en yaygın genotipi olarak tespit edilmiĢtir. CC genotipi Honamlı keçi ırkında tespit edilmemiĢtir. Gözlenen heterozigot değerleri Kıl ve Honamlı keçi ırklarında sırasıyla 0.22 ve 0.18;
beklenen heterozigotluk değerleri ise sırasıyla 0.20 ve 0.16 olarak belirlenmiĢtir. Honamlı ve Kıl Keçisi ırklarının Hardy-Weinberg dengesinde olduğu gözlenmiĢtir (P>0.05).
Naicy ve ark. (2017) çalıĢmasında, Güney Hindistan'da yetiĢtirilen toplam 277 baĢ iki yerli keçi ırklarında (Attappady Sıyah ve Malabari), Ġnsülin Benzeri Büyüme Faktörü-1 (IGF-1) gen polimorfizmini tespit eden PCR-SSCP yöntemini kullanmıĢlardır. Ġncelenen lokusta AA ve AG olmak özere iki genotip gözlenmiĢtir. IGF-1, 19 fosforilasyon bölgesi ile 49 amino asit uzunluğunda sinyal peptidi içeren salgılayıcı bir yol proteinidir. Keçi IGF-1 amino asit sekansı %83–99 oranıda diğer türlere benzemekle birlikte ruminantlarla en yüksek özdeĢliğe sahiptir. IGF-1'in nispi ekspresyonunun uterus ve karaciğerde en yüksek, bunları ovidukt ve kasların izlediğini bildirmiĢlerdir (P<0.05).
Shareef ve ark. (2018)'nın çalıĢmasında, Beetal ırkı keçilerden toplam 60 baĢ, 30 yüksek ve 30 düĢük canlı ağırlıktaki hayvanlardan, büyüme hormonunun (BH) ve Ġnsülin Benzeri Büyüme Faktörleri (IGF-1) genleri ile büyüme ile ilgili özelliklerle olan iliĢkilerin PCR-RFLP ile incelmiĢlerdir. IGF-1 geninin polimorfik olduğu bulunan ve test edilen hayvanlarda mutant (B) ve yabani (A) allellerinin frekansları sırasıyla 0.35 ve 0.65 olan üç genotip (AA, AB ve BB) tespit edilmiĢtir. ÇalıĢmada homozigot genotipin (BB), vücut ağırlığının daha yüksek (47.13±0.78kg) olduğunu bildirmiĢlerdir (P<0.05). Ardından sırasıyla 40.45±0.31 kg ve 38.18±0.37 kg vücut ağırlığı ile heterozigot (BB) ve yabani tip (AA) genotip takip etmiĢtir. En yüksek cidago yüksekliği ve vücut uzunluğu (32.25±0.33, 31.93±0.27 inç) BB genotipinde bunu AB (31.93±0.27, 31.16±0.35) ve AA (31.62±0.68, 30.87±0.86) takip edilmiĢtir. Ayrıca, maksimum gögüs çevresi de BB (33.65±0.11) genotipinde gözlenmiĢtir. GH geninin bilinen tüm nokta mutasyonları, test edilen hayvanlarda monomorfik bulunmuĢtur. Sonuçlar, IGF-1 geninde BB genotipine sahip homozigot hayvanların, bu ırk içindeki mutant (B) allelinin büyüme özellikleri üzerindeki ilave etkilerini düĢündüren büyüme özellikleri için en yüksek değerlere sahip olduğunu göstermiĢtir.
Çizelge 2.3a. Ġnsülin Benzeri Büyüme Faktörleri-1 (IGF-1) geni ile ilgili bazı çalıĢmalar Kaynak ve
PCR ürünü Irk N
Ekzon
bölge Genotip frekansları
Allel
frekansları Verimle iliskisi
AA AB BB A B Wu-Jun ve ark. (2010), 363 bç Xinjiang KeĢmir 220 310 Ekzon 4 Ġntron 4 0.277 0.236 0.486 0.395 0.604
KeĢmir keçisinde keĢmir verimi, elyaf çapı, vücut ağırlığı ile iliĢkilendirildi. Bununla birlikte, üç genotip arasındaki KeĢmir üretim özelliklerine iliĢkili değildi (P>0.05). Qiong ve ark. (2011), 233 bç Xinjiang Bogeda KeĢmir Nanjiang KeĢmir 207 279 290 Ekzon 4 Ġntron 4 0.414 0.591 0.319 0.000 0.126 0.029 0.586 0.241 0.597 0.414 0.675 0.361 0.586 0.304 0.611
Nanjing KeĢmir keçi populasyonunda, AA genotipli bireylerin keĢmir inceliği, AB genotiplerden daha düĢüktür (P<0.05). AC genotip bireylerin vücut ağırlığı, BB genotiplerden daha yüksek bulunmuĢtur (P<0.05). Alakilli ve ark. (2012) 363 bç Barki Zaribi Ardi Masri 20 20 15 15 Ekzon 3 ve Ekzon 4 - - - 0.731 0.432 0.615 0.473 0.568 0.385 0.527 0.269 # Liu ve ark. (2012), 363 bç Mongolia n cashmere 414 Ekzon 2 0.58 0.33 0.09 0.75 0.25
Genotiplerin keĢmir lif çapına etkisi istatistiksel olarak anlamlı değildir (P>0.05). AB ve BB genotipli hayvanlarında, AA genotipine sahip olanlara kıyasla daha yüksek keĢmir ağırlığı, KeĢmir lif uzunluğu ve kıl uzunluğu görülmüĢtür. Sharma ve ark. (2013) 363 bç Hindistan yetiĢtirile n keçi 80 Ekzon 4 ve Ġntron4 - - - - - # Shareef ve ark. (2018) 363 bç Beetal keçi 60 Ekzon 4 ve intron 4 0.39 0.47 0.09 0.65 0.35 IGF-1 geninde BB genotipine sahip homozigot taĢıyıcı hayvanların, bu ırk içindeki mutant (B) alelinin büyüme özellikleri üzerindeki ilave etkilerini düĢündüren büyüme özellikleri için en yüksek değerlere sahip olduğunu göstermiĢlerdir.
2.5. GH-1 Geni Polimorfizmi
Büyüme Hormonu (GH-1) keçilerde, hipofiz ön lobundaki eozinofilik hücrelerden salgılanan, protein yapısında bir hormondur. Hipofiz bezi çıkartılmıĢ olan hayvanlarda büyümenin durduğunun ve dıĢardan BH verilmesi ile büyümenin devam ettiğinin tespit edilmesi ile büyüme hormonunun büyüme üzerinde esas görevi olduğu belirlenmiĢtir. Postnatal büyüme dıĢında hipofiz bezinden salınan büyüme hormonunun, vücutta kemik, kas, yağ dokusu üzerinde etki ederek; glikoz, protein, lipit metabolizmaları, azot ve mineral dengesi üzerinde de görevlere sahip olduğu belirlenmiĢtir. Büyüme hormonu sentez ve salınımı yaĢ, cinsiyet, fizyolojik kondisyona göre değiĢtiği bilinmektedir (Møller ve Jørgensen, 2009). Bu hormonların yüksek seviyelerine sahip hayvanların geliĢmiĢ büyüme performansı sergiledikleri iyi bilinmektedir. GH-1 büyüme, metabolizma, laktasyon ve üremesi gibi farklı biyolojik iĢlemlerde hayati bir rol oynar (Ge ve ark., 2003). GH büyüme ve metabolizmanın ana düzenleyicisi olması yanında insülin benzeri büyüme faktörü (ĠGF-1) dahil olarak üzere bu çok genin ekspresyonunu module ederek büyüme hızını vücut kompozisyonu, sağlığı, süt verimini ve yaĢlanmayı da etkilemektedir. Keçiler için GH geninin bütün sekansına GenBank veri tabanında NC_030826 eriĢim numarası ile ve 1634 bç ulaĢılmaktadır (Anonymous, 2017-b). GH geni çoğu memelide 2.6 ila 3.0 kbp'dir ve keçilerde 19. kromozom üzerinde (19q22) bulunan, 5 ekzon ve 4 introndan meydana gelmektedir (Wickramaratne ve ark., 2009; Dettori ve ark., 2013). AĢağıda BH geni sekansı ve mevcut çalıĢmada çoğaltılacak gen bölgesi verilmiĢtir (ġekil 2.9).
ġekil 2.9. Keçi büyüme hormon (GH) geni yapısı (Wickramaratne ve ark., 2009)
Malveiro ve ark. (2001), çalıĢmaların, 108 baĢ Algarvia keçide, büyüme hormon (GH) geni polimorfizmi ile süt verim özellikleri arasındaki iliĢkiyi SSCP metodu ile araĢtırmıĢlardır. ÇalıĢmada GH-1 geninin 5 ekzon bölgesi incelen 1 ve 2. ekzon her biri için iki, ekzon 3 için dört, ekzon 4 için altı ve 5. ekzon için beĢ yapısal model bulmuĢlardır. Bu sonuĢlardan elde edilen bu modeller ile süt, süt yağ ve süt protein verimi ile yağ ve protein içeriği karĢılaĢtırılmıĢtır. 4. ekzon FF modeli ve 5. ekzon AA modeli ile süt verimi arasında pozitif bir iliĢki bulunmuĢtur (P<0.05).