T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
ARPA GENOTĠPLERĠNDE TUZ TOLERANSININ FĠZYOLOJĠK ANALĠZLERLE BELĠRLENMESĠ
Zeynep Gül YEĞĠN YÜKSEK LĠSANS TEZĠ Tarla Bitkileri Anabilim Dalı
Ocak-2012 KONYA
iv ÖZET
YÜKSEK LĠSANS TEZĠ
ARPA GENOTĠPLERĠNDE TUZ TOLERANSININ FĠZYOLOJĠK ANALĠZLERLE BELĠRLENMESĠ
Zeynep Gül YEĞĠN
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Tarla Bitkileri Anabilim Dalı
DanıĢman: Yrd. Doç. Dr. Mustafa YORGANCILAR 2012, 74 Sayfa
Jüri
Yrd. Doç. Dr. Mustafa YORGANCILAR Prof. Dr. Süleyman SOYLU Yrd. Doç. Dr. Semiha ERĠġEN
Bu araĢtırmada bazı arpa genotiplerinin tuza toleransları araĢtırılmıĢtır. ÇalıĢmada S.Ü. Ziraat Fakültesi Tarla Bitkileri Bölümünde ıslah çalıĢmaları sonucu geliĢtirilen Soylu-10 ve Soylu-11 ile Konya ilinde yaygın olarak yetiĢtirilen Anadolu, Konevi, Larende ve Tokak çeĢitleri olmak üzere 6 arpa genotipi kullanılmıĢtır. Tohumlar perlit içeren saksılara ekilmiĢ ve çıkıĢtan sonra farklı NaCl konsantrasyonları (0, 25, 50 ve 75 mM) içeren half hoagland çözeltisi ile sulanmıĢtır. Onuncu hafta sonunda bitkilerin morfolojik ve fizyolojik parametreleri incelenmiĢtir. AraĢtırma sonuçlarına göre; bağıl su içeriği (BSĠ), klorofil değeri (KD), stoma iletkenliği (SĠ), Potasyum (K) içeriği ve köklerde kalsiyum (Ca) içeriği bakımından kontrol grubu bütün genotiplerde daha yüksek değerler vermiĢtir. Prolin miktarındaki artıĢ; arpa bitkisinin tuz stresine olan tepkisi ve stres koĢullarının verdiği zararı en aza indirmek amacıyla oluĢturduğu savunma mekanizmasının harekete geçmesi olarak nitelendirilmiĢtir. Na ve Cl değerleri bütün genotiplerde tuz konsantrasyonları arttıkça bitki ve köklerde artıĢ olmuĢtur. AraĢtırmada bitki ve kök uzunluğu, bitki ve kök kuru ağırlığı, bitki ve kök yaĢ ağırlığı sonuçları tuz stresini belirlemede önemli bir kriter olarak belirlenmiĢtir. Fotosentetik verim üzerine yapılan araĢtırmada tuz stresinin altı genotipe etkisi olmadığı belirlenmiĢtir. Genotipler tuz stresi altında farklı dayanıklılık mekanizmaları geliĢtirmiĢlerdir. Bazı genotipler yüksek tuz uygulamasından az etkilenirken bazıları daha etkilenmiĢtirler.
Sonuç olarak yapılan çalıĢmada çeĢit adayı olan arpa hatlarının tuza tepkileri belirlenerek, tolerans sınırları ortaya konmaya çalıĢılmıĢtır. Yapılan çalıĢmalar sonucunda 10. ve Soylu-11 tuza hassas olduğu belirlenmiĢtir. Bu iki genotipin ekim alanlarındaki tuz oranları düĢük değil ise bunların ekiminde ekonomik karlılık görülmemektedir. Tokak 157/37 çeĢidi de sonuçlara göre hassas bir çeĢit olduğu belirlenmiĢtir. Larende, Anadolu 98 ve Konevi çeĢitleri de tuzlu alanlarda kullanılabilecek en uygun çeĢitlerdir.
v ABSTRACT MS THESIS
DETERMINATION OF SALT TOLERANCE OF BARLEY GENOYTYPES WITH PHYSIOLOGICAL ANALYSIS
Zeynep Gül YEĞĠN
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY
THE DEGREE OF MASTER OF SCIENCE / DOCTOR OF PHILOSOPHY IN MECHANICAL ENGINEERING
Advisor: Asst. Prof. Dr. Mustafa YORGANCILAR 2012, 74 Pages
Jury
Asst. Prof. Dr. Mustafa YORGANCILAR Prof. Dr. Süleyman SOYLU
Asst. Prof. Dr.Semiha ERĠġEN
In this search the tolerances to the salt of different barley genotypes are investigated. In the search total 6 different genotypes are used. These genotypes are Soylu 10 and Soylu 11, Anadolu, Konevi, Larende and Tokak varieties. The seeds are planted in pots with perlite. Once shoot the plants are irrigated half Hoagland solution with different NaCl concentrations. (0, 25, 50 and 75 mM). At the stage of 10th week end the physiologic parameters of the plants are observed. The results of the research the control group have shown higher values in relative leaf water (RWC) stomatal conductance, chlorophyll amount, potassium content and calcium content in roots. The augmentation of proline quantity is characterized as defense system of the plant to inhibit the damage effects of salt stress. When the Na and Cl values are increasing in roots and shoots in all genotypes when the salt concentrations increases. The plants roots and shoots lengths, as well the dry and wet weights of roots and shoots are observed as the important factors to determine the salt stress. The observation shoved that the salt stress has no effect on photosynthetic yield. Genotypes have developed different mechanisms of resistance under salt stress. As some genotypes are more affected than others to salt stress.
The aim of this experiment is to determine the behavior different genotypes of barley in salt and find their tolerance limits. All the facts are showing us that the 10th and 11th Lines are sensible to salt. Those 2 genotypes are economically not profitable if the salt level in the land where they are cultivated is high. Tokak 157/37 genotype is determined as salt sensible. Larende, Anadolu 98 and Konevi genotypes are the best cultivable ones in salty areas.
vi ÖNSÖZ
Arpa bitkisinde yürütülecek çalıĢmalara örnek teĢkil edecek bu çalıĢmamda; bilgi ve tecrübeleriyle bana yol gösteren danıĢman hocam Sayın Yrd. Doç. Dr. Mustafa YORGANCILAR’a, çalıĢmamın her aĢamasında desteğini esirgemeyen danıĢman hocam kadar emeği geçen hocam Sayın Prof. Dr. Süleyman SOYLU’ya, fizyolojik çalıĢmalarımda yol gösteren Yrd. Doç. Dr. Mehmet HAMURCU’ya, laboratuar çalıĢmalarımda yardımcı olan hocam Dr. Emine ATALAY’a ve tecrübeleriyle bana destek olan Yrd. Doç. Dr. Semiha ERĠġEN hocama, 11201029 No’lu Proje ile tezimi maddi olarak destekleyen Selçuk Üniversitesi Bilimsel AraĢtırma Projeleri Koordinatörlüğü’ne, ayrıca çalıĢmalarım süresince beni yalnız bırakmayan, maddi ve manevi destekleriyle bana güç veren aileme sonsuz teĢekkürlerimi sunarım.
Zeynep Gül YEĞĠN KONYA-2012
vii ĠÇĠNDEKĠLER ÖZET ... iv ABSTRACT ... v ÖNSÖZ ... vi ĠÇĠNDEKĠLER ... vii 1. GĠRĠġ ... 1 2. KAYNAK ARAġTIRMASI ... 5 3. MATERYAL VE METOD ... 20 3.1. Materyal ... 20 3.2. Metot ... 20 3.2.1 Denemelerin kurulması ... 20 3.2.1.1. Saksıların hazırlanması ... 20 3.2.1.2. Tohumların hazırlanması ... 21
3.2.1.3. Besin çözeltisinin hazırlanması ... 21
3.2.1.4. Ekim ve kültür Ģartları ... 21
3.2.2. Gözlem ve ölçümler ... 21
3.2.2.1. Büyüme parametrelerinin ölçülmesi ... 21
3.2.2.1.1. Uzunluk (cm) ... 22
3.2.2.1.2. YaĢ ağırlık (g) ... 22
3.2.2.1.3. Kuru ağırlık (g) ... 23
3.2.2.2. Fizyolojik özellikler ... 23
3.2.2.2.1. Bağıl su içeriği (RWC) (%) ... 23
3.2.2.2.2. Klorofil değeri (spad) ... 24
3.2.2.2.3. Stoma iletkenliği (mmol/m2sn) ... 24
3.2.2.2.4. Fotosentetik verim (klorofil flüoresansı) (Fv/Fm) ... 25
3.2.2.2.5. Prolin analizi (µmol/g) ... 26
3.2.2.3. Element analizi (Cl, K, Ca ve Na) ... 26
3.2.2.3.1. Klor analizi ... 26
3.2.2.3.2. Sodyum (Na), Potasyum (K) ve Kalsiyum (Ca) Analizi ... 26
3.2.2. Ġstatistiki analiz ve değerlendirmeler ... 27
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 28
4.1. Büyüme Parametreleri ... 28
4.1.1. Bitki ve kök uzunluğu ... 28
4.1.2. Bitki ve kök yaĢ ağırlık ... 32
4.1.3. Bitki ve kök kuru ağırlık ... 35
4.2. Fizyolojik Analizler ... 39
4.2.1. Bağıl su içeriği ... 39
4.2.2. Klorofil değeri ... 41
4.2.3. Stoma iletkenliği ... 43
viii
4.2.5. Prolin analizi ... 46
4.3. Element Analizleri (Cl, Na, K ve Ca) ... 49
4.3.1. Klor (Cl) ... 49 4.3.2. Sodyum (Na) ... 50 4.3.3. Potasyum (K) ... 54 4.3.4. Kalsiyum (Ca) ... 58 5. SONUÇLAR VE ÖNERĠLER ... 62 5.1 Sonuçlar ... 62 5.2 Öneriler ... 63 KAYNAKLAR ... 66 ÖZGEÇMĠġ ... 75
1. GĠRĠġ
Arpa, hububat üretimimizin %27.7’sini oluĢturmaktadır. Türkiye’nin tüm bölgelerinde yetiĢtirilmekle birlikte, özellikle Orta Anadolu (Konya, Ankara, EskiĢehir ve KırĢehir) ve Güneydoğu Anadolu Bölgesinde üretimi daha fazladır. Ülkemizde arpa ekim alanı 3.5 milyon ha, üretim ise 7 milyon tondur. Geleneksel bir tarım ürünü olan arpanın ekim alanı, üretim ve verimi yıllar itibari ile sürekli artıĢ göstermektedir. Bu nedenle arpanın kuraklık ve tuzluluk stresine karĢı toleransının aydınlatılması çok önemlidir.
Tuzluluk; özellikle kurak ve yarı kurak iklim bölgelerinde yıkanarak yeraltı suyuna karıĢan çözünebilir tuzların yüksek taban suyuyla birlikte kapillarite yoluyla toprak yüzeyine çıkması ve buharlaĢma sonucu suyun topraktan ayrılarak tuzun toprak yüzeyinde ve yüzeye yakın bölümünde birikmesi olayıdır (Ergene,1982; Kwiatowsky, 1998; Kara, 2002).
Dünyanın yaklaĢık %7’si, kültüre alınmıĢ alanların %20’si ve sulama yapılabilen alanların neredeyse yarısı tuzluluktan etkilenmektedir (Zhu, 2001). Ülkemizde ise tuzlu topraklar toplam 1.5 milyon ha alanı kapsamaktadır (TÜĐK 2004). Yüksek tuz konsantrasyonlarında tüm bitki türleri belirgin büyüme inhibisyonları göstermesine karĢın, büyüme ve üretkenlik açısından türler ve çeĢitler arasında belirgin farklılıklar bulunmaktadır (Munns, 2002). Tuza tolerans için agronomik seleksiyon parametreleri arasında verim, canlılığın sürdürülmesi, bitki boyu (Noble ve Rogers, 1992), nispi büyüme oranı (Cramer ve ark., 1990), nispi büyüme inhibisyonu (He ve Cramer, 1992; Bağcı ve ark., 2003), yaprak yüzey alanı (Franco, 1993), hücresel zarar (Munns, 1993) ve Na+/K+ oranı (Joshi ve ark., 1979; Kholová ve ark., 2010) gösterilmektedir. Bununla birlikte, bir bitki türü hücre, doku veya tüm bitki seviyesinde tuza toleransın ayırt edici indikatörlerine sahipse, seleksiyonun çok daha elveriĢli ve uygulanabilir olduğu ileri sürülmüĢtür (Munns, 2002; Ashraf ve Harris, 2004).
Dünya toplam alanının (135 milyon km2) 4 milyon km2’si tuzluluk sorunu yaĢamaktadır. Türkiye’de ise toplam 778 bin km2’lik (78 milyon ha) alanının 8 bin km2’si (800 bin ha) yani %1’lik kısmı tuzluluk sorunu yaĢamaktadır (FAO, 2005). Türkiye’deki tuzlu veya sodyumlu arazi miktarı 1.270.000 hektardır. Bunlar bütün ülke yüz ölçümünün % 1.62’sini kapsar. Sözü edilen bu alanlar, genellikle sulamaya elveriĢli
çukur ovalardan, kapalı havzalardan veya derin aluviyal topraklardan oluĢmaktadır (BeĢer, 2003).
Tuzluluk, doğal yolla oluĢabildiği gibi sulama uygulamalarının yanlıĢ yapılması sonucu insan kaynaklı da ortaya çıkabilmektedir. Ekonomik yönden önemli ürünlerin yetiĢmesi amacıyla yağıĢlar yetersiz geldiğinde sulama yoluna gidilmektedir. Ancak bu uygulama topraklarda çeĢitli tuz iyonlarının birikimine neden olmaktadır. Sulama suyundaki çözünmüĢ madde konsantrasyonu yüksekse, biriken tuzlar drenaj sistemi ile yıkanmazsa tuzluluk oranında artıĢ görülebilmektedir. Özellikle doğal drenaj koĢullarının kötü olduğu kurak bölgelerde buna daha çok rastlanmaktadır. Yapılan çalıĢmalarda dünyada sulanan alanların % 20’sinin tuzluluktan etkilendiği rapor edilmiĢtir (Pitman ve Lauchli, 2002).
Toprakların tuzlulaĢmasında, bilinçsiz sulama yanında, drenaj olanaklarının yetersizliği ve yüksek taban suyunun da rolü çok büyüktür. Özellikle, sulama sonucu toprakların tuzlu ve alkali hale dönüĢmesi, sulu tarımın uygulandığı bölgelerde güncel bir sorundur. Drenaj Ģebekelerinin yetersizliği ve sulama sonucu yükselen taban suyu, kurak bölgelerde tuzluluğun baĢlıca nedenidir. Bitki kök bölgesinde fazla miktarda eriyebilir tuzların birikmesi, bilindiği gibi, toprakta tuzluluk sorununun ortaya çıkmasına neden olmaktadır. Böyle bir toprakta, kültür bitkilerinin çimlenme, büyüme ve ürün verimleri, mevcut tuzların cinsi ve miktarlarına bağlı olarak azalmakta ve hatta tamamen durmaktadır (Richards, 1954; Dizdar, 1978).
Yarı kurak iklim koĢullarında sulama yapılan alanlarda önemli bir sorun olan tuzluluğun potansiyel etkisi, sadece ürün verimi üzerine değil, aynı zamanda arazilerin tuzlulaĢması, toprağın ve suyun bozulması ve yer altı sularına tuzun karıĢarak kalitelerinin bozulmasına neden olmaktadır (Feng ve ark., 2003). Aynı zamanda tuzluluğun neden olduğu arazi bozunması sonucu gıda üretimi olumsuz bir Ģekilde etkilenmektedir. Kurak ve yarı kurak alanlarda biriken tuzlu taban sularının uzaklaĢtırma Ģansı olmadığında ciddi bir problem oluĢturmakta ve farklı kullanımlar için ihtiyaç duyulan kaliteli suya olan talebi artırmaktadır (Sharma ve ark., 1993 ve 1994).
Toprak tuzluluğu dünyadaki tarımsal üretimi sınırlayan temel çevresel streslerden biridir (Lauchli ve Grattan, 2007). Toprakta biriken tuzlar toprağın fiziksel ve kimyasal özelliklerini bozmakta ve bitki geliĢimini de olumsuz yönde etkilemektedir. Toprakta bulunan tuzlar su potansiyelinde azalmaya neden olup topraktan mineral
alınımını sınırlamaktadırlar. YetiĢtirilen bitkinin veriminde görülen azalmalar, toprak çözeltisinin konsantrasyonuna bağlı olduğu kadar, bitkinin tuza dayanıklılığı ile de ilgilidir (Ekmekçi, 2005). Bitkilerin tuza dayanımlarının incelenmesi özellikle toprak tuzluğunun belirli bir düzeyin altına düĢürülmediği alanlarda ekonomik düzeyde ürün verebilecek bitkilerin seçilerek yetiĢtirilmesi amacıyla önemlidir (Kotuby ve ark., 1997).
YetiĢtirme ortamının aĢırı tuzlu olması fizyolojik birçok olumsuz etkiyi de beraberinde getirir. Bu olumsuz etkiler arasında membran disfonksiyonu, ozmotik uyumsuzluk ve su alımında dengesizlik, genel metabolik süreçte aksamalar, besin dengesizliği, enzim aktivasyon bozukluğu ve bitkide genel geliĢim bozukluğu olarak sıralanabilir. Tuz stresi altındaki bitkilere dıĢarıdan uygulanan Ca, K veya fosfor (P) içeren bileĢiklerin, bitkinin yaprak ve köklerinde Na ile rekabete girerek onun alınımını azalttığı ve bitki bünyesinde Ca, K ve P iyonlarının strese karĢı koyabilecek yeterli düzeylere ulaĢmasıyla birlikte bitkinin strese karĢı koyabilme kapasitesinin de arttığı bazı çalıĢmalarla bildirilmiĢtir. (Yakıt ve Tuna, 2006).
Arpa (Hordeum spp), çoğunlukla fizyolojik ve genetik çalıĢmalar için kullanılan model bir bitkidir (Koornneef ve ark., 1997). Kısa yaĢam döngüsüyle tek yıllık olan arpa 7 çift kromozom içerir ve diploiddir. Buğday, mısır ve pirinçten sonra tarımı yapılan dünyanın en önemli dördüncü tahıl ürünüdür. Taksonomik yönden Poaceae familyasına ait olan arpanın dünyadaki yıllık üretimi 140 milyon tonun üzerindedir (Li ve ark. 2007). Yarı kozmopolit olup 20 kadar tür içermektedir. H. vulgare L. ülkemizde geniĢ oranda kültürü yapılan tek yıllık bitkidir.
Arpa, insanlar için besin kaynağı oluĢtururken hayvanlar içinde yem olarak kullanılmaktadır. Bunun yanı sıra bira ve viskinin de hammaddesidir. Diğer ürünlerden daha geniĢ ekolojik dağılıma sahip olan arpa fazla soğuk ve fazla sıcak olmayan, nispi nemi yüksek olan yerlerde iyi geliĢir. Buğdayın yetiĢemediği topraklarda yetiĢebilme özelliği vardır. Kuraklığa ve tuzluluğa olan toleransı nedeniyle arpa dünyanın her köĢesinde kurak olan bölgelerde yetiĢmektedir. Sıcaklığı 0ºC’nin altına düĢmeyen ve 18-20ºC’nin üzerine çıkmayan, nispi nemi %70 - 80 olan yerler arpa için çok uygundur.
Tuzluluk problemi, kurak ve yarı kurak bölgelerde yağıĢın az, buharlaĢmanın fazla olduğu alanlarda doğal olarak ortaya çıkmakta ve bu koĢullara ilaveten, sulamaya açılan bölgelerde ise aĢırı sulama ile taban suyundaki tuzların üst katmanlara çıkıĢı ile oluĢmaktadır. Bu da tarım alanlarında tarımı yapılan çoğu kültür bitkisi yetiĢtiriciliğinde sorunlara neden olmaktadır. Bu sebeple oluĢacak tarımsal kayıpları önlemek için bu
alanların ıslah edilmesi ya da tuza toleranslı bitkilerin belirlenerek tuzluluk problemi olan alanlarda bu bitkilerin tercih edilmesi gerekliliğini ortaya koymaktadır. Bu nedenlerle tuzlu alanlarda yetiĢtirilebilecek arpa genotiplerinin belirlenmesi amacıyla yapılan araĢtırmada NaCl tuzluluğuna en toleranslı ve en hassas arpa çeĢitleri belirlenmiĢ ve çeĢitler arasındaki farklılıklar ortaya konmuĢtur. ÇalıĢmada S.Ü. Ziraat Fakültesi Tarla Bitkileri Bölümünde ıslah çalıĢmaları sonucu geliĢtirilen ve çeĢit adayı olan arpa hatları (Soylu-10 ve Soylu-11) ile Konya ilinde yaygın olarak yetiĢtirilen bazı arpa çeĢitleri (Anadolu, Konevi, Larende ve Tokak) farklı NaCl konsantrasyonlarına (0, 25, 50 ve 75 mM ) maruz bırakılmıĢ ve fizyolojik değiĢimler incelenmiĢtir.
2. KAYNAK ARAġTIRMASI
Rains (1972)’a göre, yüksek tuz konsantrasyonu koĢullarında bulunan bitkiler, iyon toksisitesinin yanı sıra osmotik strese girmekte ve su noksanlığından kaynaklanan fizyolojik bozukluklar yaĢamaktadır. Artan iyon alımı ile osmotik stresin giderilmesi ve böylece hücre turgorunda azalma olmadan bitkinin geliĢmesini sürdürebilmesine “osmotik uyum” adı verilmektedir.
Çevresel streslere maruz kalan birçok bitki prolin biriktirir. Prolin birikimi bitkinin çeĢidine, stresin süresine ve Ģiddetine bağlı olarak değiĢmektedir. Yapılan çalıĢmalarda da stres koĢullarında farklı arpa genotipleri arasında prolin biriktirme hızında ve miktarında büyük farklılıklar olduğu belirlenmiĢtir (Singh ve ark., 1973a).
Bates ve ark. (1973)’na göre su stresindeki bitkilerde diğer aminoasitlere göre nispeten daha hızlı artan prolinin, çok etkili stres karakterlerinden biri olduğunu ve kuraklık ölçüsünün tek parametresi olarak kullanıldığını açıklamıĢlardır. DeğiĢik su stresi, bitkilerdeki sayısız metabolik düzensizlikleri teĢvik etmektedir.
Hellebust (1976)’un bildirdiğine göre, osmotik uyum, bitkilerin K ve Na gibi bazı inorganik iyonlar ya da gliserol, sukroz, prolin, betain gibi bazı organik iyonları biriktirebilme yeteneği olarak tanımlanmaktadır.
Tuzlu koĢullarda yetiĢtirilen bitkilerin iyon dengesinin bozulmasına paralel olarak mineral madde konsantrasyonlarında da önemli sayılabilecek oranlarda değiĢimler olmaktadır. Tuz stresinden etkilenmeyen ya da göreceli olarak daha az etkilenen bitkilerin dokularında Na ve Cl iyonları daha az, prolin miktarı ise daha fazladır (Flowers ve ark., 1977).
Pek çok araĢtırıcı tuzluluğun bitki büyüme ve geliĢmesinde meydana getirdiği olumsuz etkiyi azaltmaya ya da tamamen ortadan kaldırmaya yönelik çalıĢmalar yapmıĢlar ve bu amaçla çimlenme ve büyümeyi olumlu yönde etkilediği bilinen büyüme hormonlarını kullanmıĢlardır (Bozcuk, 1978; Kabar, 1987).
Wyne Jones ve ark. (1978)’e göre; prolinin osmoregülasyon yoluyla etkili olduğunu ileri sürerken; Schobert ve ark. (1978) proteinlerin stabilizasyonuna etki ettiğini; Ahmad ve ark. (1982) ile Paleg ve ark. (1981) ise inaktivasyonuna ve tuza karĢı hücre zarlarını ve stoplazmik enzimleri koruduğunu bildirmektedirler. Bu görüĢlere paralel olarak Handson ve ark. (1982) stres toleransının ölçülmesinde prolin birikiminin bir parametre olabileceği görüĢünü savunmaktadırlar.
Tuz stresinde bitkilerde aĢırı miktarlarda biriken Na, potasyumun alınımını (Siegel ve ark., 1980), Cl- ise özellikle NO3 alımını engelleyerek (Kirkby ve Knight 1987; GüneĢ ve ark., 1994; Ġnal ve ark., 1995) bitkilerin iyon dengesinde bozulmalara neden olabilmektedir. Tuzlu topraklarda artan osmotik potansiyelden dolayı bitkilerin suyu yeteri kadar kullanamaması ya da ortamda aĢırı miktarlarda bulunan Na ve Cl’nin neden olduğu toksik etkiden dolayı üründe azalma olmaktadır (Flowers ve ark.,1981, Lewitt, 1980).
Tuz stresinde Na alımı diğer mineral maddelerin alımı ile rekabete girerek beslenme noksanlığına yol açmaktadır. Bitkiler osmotik stres elimine edilerek tuz stresine maruz bırakıldıklarında büyümede meydana gelen azalmalar ve bu azalmaların K uygulamaları ile iyileĢtirilmesi, NaCl’ün bitkilerde K noksanlığına yol açtığına iĢaret etmektedir (Levitt, 1980).
Prolinin koruyucu rolünü bazı bitkilerde glisinbetain yada sorbitol gibi baĢka bileĢikler üstlenebilir. Bu nedenle tüm bitkilerde prolin birikimi gözlenmeyebilir. Prolinin uyuma iliĢkin rolü büyümenin devamından çok canlılığın sürdürülmesi ile iliĢkilidir. Prolin birikiminin tuza toleranslılarda tuza duyarlılardan daha fazla olduğu yolunda yeterli kanıt yoktur Aynı araĢtırmacı tuzluluğun çimlenmeyi azalttığı, bitki geliĢimini ve verimi düĢürdüğünü belirtmiĢtir (Greenway ve Munns, 1980).
Tuzluluk, bitkilerin klorofil içeriğinde ve fotosistem-II aktivitesinde azalmaya neden olmaktadır (Smillie ve ark., 1982). Hücresel inorganik iyon miktarı ve hücresel bileĢim ile prolin birikimi arasında bir iliĢki bulunmaktadır. Bununla ilgili olarak Na+ yada K+ miktarı bir eĢik konsantrasyonuna ulaĢıncaya kadar yapraklarda prolin birikiminin baĢlamadığı, bununla birlikte prolin birikimini baĢlatacak eĢik değerinin türden türe farklılık gösterdiği bilinmektedir (Buhl ve ark., 1983; Coughlan ve ark., 1980).
Çimlenme devresinden sonraki geliĢme devrelerinde tuza dayanıklı olan bitkiler çimlenme esnasında toprak tuzluluğuna karĢı oldukça duyarlıdırlar (Tekinel ve Çevik, 1983). Tuzlu alanlara uygun çeĢit ve tür seçiminde bu özellik dikkate alınmalıdır.
Bitki dokularında prolin birikiminin kuraklık, tuzluluk ve düĢük sıcaklık gibi çevresel streslerle teĢvik edildiği ve tuz stresine cevap olarak hem halofitlerde hem de halofit olmayan bitkilerde prolin biriktirildiği bildirilmiĢtir (Buhl ve Stewart, 1983). Buna ilaveten yüksek seviyedeki prolinin, kuraklık ve tuzluluk koĢulları altında osmoregülasyonu, proteinlerin stabilizasyonunu, enzimlerin ısı denatürasyonun
önlenmesini ve stresten sonraki enerji ve azotun korunmasını sağladığı belirtilmiĢtir (Aloni ve Rosenshtein, 1984).
Bulh ve Stewart (1983), prolin birikimi miktarının, tuzluluk derecesiyle yakından ilgili olduğunu ileri sürmüĢlerdir. Aynı araĢtırıcılar arpa ile yaptıkları çalıĢma sonucunda, tuz stresinin prolin birikimine neden olduğunu bulmuĢlardır.
Bitki tarafından alınan aĢırı miktardaki tuzun hücre fonksiyonlarını bozması, hücre ve organel zarlarında meydana gelen tahribatlar nedeniyle fotosentez, solunum vb. iĢlevlerin sekteye uğraması tuz zararının baĢka bir sonucudur (Leopold ve Willing, 1984).
Seemann ve Crithley (1985) ile Aranda ve Stvertsen (1996), yüksek tuz konsantrasyonlarında iyon birikimi ve stomaların açılıp kapanmasındaki düzensizlik nedeniyle toplam korofil miktarında azalmalar olduğunu ve bunun sonucu olarak fotosentez etkinliğinin azalarak bitkinin geliĢmesinde gerilemeler ortaya çıktığını açıklamaktadırlar.
Tuza toleransın belirlenmesinde bitki doku ve organellerinde iyon (Na+
, K+ ve Cl-) birikimi, bitkide taĢınımı ve dağılımı ile bu iyonların birbirine olan oranları (K/Na) (Hasegawa ve ark., 1986), bitkilerin organik madde biriktirme ve sentezleme yetenekleri ile hücre düzeyinde meydana gelen oksidatif stresten kaynaklanan zararlanmalar üzerinde durulmaktadır.
Tuzluluk, sıcaklık, kuraklık, UV-radyasyon ve yaralanma gibi biyotik ve çevresel streslere maruz kalan bitkilerde H2O2, söz konusu streslere karĢı tepki gösteren önemli bir moleküldür. Bir çok araĢtırıcı (Federico ve Angelini, 1986; Svalheim ve Robertsen, 1993; Laurenzi ve ark., 1999) bitki hücrelerinde lignin biyosentezinin ve hücre duvarı sertleĢmesinin H2O2’e bağlı olduğunu tutarlı bulgularla rapor etmiĢlerdir.
Potasyum konsantrasyonunun bitki bünyesinde artması, bitkinin tuza dayanıklılığını arttırmaktadır (Hsiao ve Lauchli, 1986). Ayrıca bitkinin sahip olduğu yüksek K+/Na+ oranının tuza dayanıklılıkla doğru orantılı olduğu bilinmektedir (Gorham, 1990; Ashraf ve ark., 1997; Sherif ve ark,. 1998).
Munns ve ark. (1986) ile Neumann ve ark. (1988), tuzun etkisi ile bitki büyümesinin azalması, zararlanan ya da ölen yaprakların oranının, yeni geliĢen yapraklardan fazla olması nedeniyle azalan fotosentez alanıyla da açıklamaktadırlar.
Tuzluluk, çoğunlukla yapraklarda erken yaĢlanmaya neden olmaktadır (Sahu ve Mishra, 1987; Yeo ve ark., 1991). Tuz stresinin neden olduğu yapraklardaki erken yaĢlanma ile lipid peroksidasyonu ürünü olan malondialdehit (MDA) arasındaki bir bağlantıdan bahsedilmektedir. MDA birikimi, iyon sızması (relative leakage ratio=RLR) ile paralellik göstermektedir.
Tuz stresi, yapraklarda erken yaĢlanmaya neden olduğu gibi (Sahu ve ark., 1987; Yeo ve ark., 1991). Chen ve ark. (1989)’ göre genellikle protein veya klorofil konsantrasyonundaki azalma ve hücre zarı geçirgenliğindeki artıĢla (Dhindsa ve ark., 1981) ifade edilmektedir.
Neuman ve ark. (1988), fasulye fidelerinin düĢük düzeylerde (50-100 mM) tuzluluğa maruz bırakıldığında, üç günlük süre içerisinde ıĢıklı koĢullarda primer yapraklarda hücre geniĢlemesinin azaldığını bildirmekte ve bunu hücre büyüme parametrelerinden hücre duvarı büyümesi ve turgora bağlamaktadırlar. Tuzluluk 72 saat sonra hücre büyümesinde herhangi bir olumsuz etki meydana getirmemiĢ, kontrol bitkilerinde oransal olarak küçük artıĢlar görülmüĢtür. Diğer yandan 50 mM NaCl 24 saat içinde toplam yaprak turgorunu önemli ölçüde azaltmıĢ, yaprak osmotik potansiyelinde adaptif azalmaya paralel olarak ksilem basıncı, yapraktaki apoplastik madde potansiyelini düĢürmüĢ ve bu düĢüĢ de sonuçta yaprağın su potansiyelini düĢürmüĢtür. Bu bulgularla, fasulye fidelerinde orta tuzluluk seviyelerinde baĢlangıçta yaprak büyüme oranının, hücre duvarı büyümesindeki azalmadan çok turgordaki düĢüĢten kaynaklandığı ve uzun dönem tuzluluk koĢullarında (10 gün) turgordaki azalmaya ters olarak hücre duvarı büyümesinin sağlandığını ifade etmiĢlerdir.
Tuzlu koĢulların çimlenme ve büyüme üzerinde meydana getirdiği olumsuz etkinin poliaminin grubu maddelerde de ortadan kaldırılabileceğini literatürde belirtmiĢlerdir (Prakash ve Prathapasenan, 1988a; Prakash ve Prathapasenan, 1988b; Krishnamurthy, 1991; Mishra ve Sharma, 1994).
Çevresel faktörler ve fizyolojik etkilerin eĢlik ettiği tuza tolerans özelliğinin esas kaynağı kalıtsal unsurlardır. Tuza tolerans bakımından bitkiler arasında önemli farklılıklar bulunmaktadır. Familya, cins ve türler arasında farklılıklar bulunduğu gibi, aynı türe ait genotipler arasında da tuza tolerans yönünden farklılıkların bulunduğu bilinmektedir (Shalata ve Tal, 1988).
Son yıllarda yapılan stres fizyolojisi çalıĢmalarına konu olan prolin; su, tuz, ıĢık, kirlilik, patojen enfeksiyonları ve sıcaklık streslerinin bir indikatörü olarak kabul edilmektedir (Amberger ve ark.,1998; Axbazhagan ve ark., 1988).
Tuz stresi hücresel düzeydeki mevcut suyun osmotik olarak bağlanmasına ve bitkilerin fizyolojik olarak karıĢıklığa girmesine neden olur (Cheeseman, 1988). Su yetersizliği süperoksit, hidrojen peroksit, hidroksil radikal (Halliwell ve Gutteridge, 1985) ve singlet oksijen (Elstner, 1987) gibi reaktif oksijen türlerinin sitoplazmada ve suya dismutasyona uğratılır (Imlay ve Linn, 1988). Bu sitosolik aktive edilmiĢ oksijen türleri lipidler (Fridovich, 1986; Wise ve Naylor, 1987), protein ve nükleik asitlerde (Imlay ve Linn, 1988) oksidatif hasarlanma yoluyla normal metabolizmayı ciddi bir biçimde bozabilir. Ġnternal O2 konsantrasyonları fotosentez sırasında yüksek olduğundan (Steiger ve ark., 1977) kloroplastlar genellikle aktive edilmiĢ oksijen türlerini üretme eğilimindedir (Asada ve Takahashi, 1987). Süperoksit üretildiğinde enzimatik ya da enzimatik olmayan proseslerle hızlı bir Ģekilde H2O2 ve suya dismutasyona uğratılır (Imlay ve Linn, 1988).
Bitkiler reaktif oksijen türlerine çeĢitli seviyelerde antioksidanlarla karĢı koyarlar. Metaloenzim süperoksit dismutaz süperoksidi H2O2’ ye dönüĢtürür. Katalaz ve çeĢitli tipteki peroksidazlar H2O2’nin bozulmasını katalizler (Chang ve ark., 1984), katalaz kloroplastlarda olmadığından dolayı bu organelde hidrojen peroksit askorbata spesifik peroksidazlarla askorbat-glutatyon döngüsü boyunca detoksifikasyona uğratılabilir (Chen ve Asada, 1989; Asada, 1992). Askorbat ve glutatyonun her ikisinin de kloroplastlarda milimolar düzeylerde olduğu rapor edilmiĢtir (Halliwell, 1982). Askorbat okside α-tokoferolün α-kromoksil radikalinin bir indirgeyicisi gibi iĢlev yaparak ya da süperoksit radikali ile direkt reaksiyona girme yoluyla da okside edebilir (Foyer ve ark., 1991). Tilakoid membranlar α-tokoferol bakımından zengindir. Bu antioksidan sadece süperoksitle reaksiyona girmez, bunun yanında hidroksil, peroksil ve alkoksil radikallerinin temizlenmesine de yardımcı olur (Halliwell, 1987).
Mangal ve Lal (1990) ile Awang ve ark. (1993), hormon dengesinde ortaya çıkan değiĢikliklerin tohum çimlenmesi üzerinde de etkide bulunduğunu, azalan sitokinin sentezlenmesinin sonucu olarak çimlenme oranında azalma oluĢtuğu iddia edilmektedir. Tuzlu koĢullarda çimlenmenin engellenmesi ve çimlenme yüzdesinin düĢmesi, beklenen bir tepkidir (Demir ve Demir, 1992).
Sharma ve ark. (1990), klor ve sülfat tuzluluğunun dominant olduğu değiĢik saksılarda yetiĢtirdikleri iki farklı nohut çeĢidinin her ikisinde de klorofil, karbonhidrat, niĢasta ve protein miktarının azaldığını, aksine amino asit ve serbest prolin oranının artan tuzluluk ile beraber arttığını tespit ettiklerini bildirmekte ve klorun sülfattan daha fazla olumsuz etkide bulunduğunu ve denenen çeĢitlerin tuzluluğa farklı reaksiyon gösterdiğini belirtmektedirler.
Tuz stresi atında bulunan bitkilerde serbest oksijen radikallerine karĢı bitkiyi koruyan enzim aktivitelerinin dayanıklı genotiplerde daha yüksek olduğu, yapılan araĢtırmalarla belirlenmiĢtir (Cakmak ve Marschner, 1992; Polle ve ark., 1992; Gossett ve ark., 1994a; Gosset ve ark., 1996). Süper oksit dismütaz (SOD), askorbat peroksidaz (APX), glutatyon redüktaz (GR), katalaz (CAT) gibi enzimler serbest oksijen radikallerinin yok edilmesinde en etkin antioksidatif enzimler olarak bilinmektedirler (Cakmak ve Marschner, 1992; Cakmak, 1994; Gossett ve ark., 1994a).
Tuz stresinin yaprak yaĢlanması üzerindeki özel etkisi, toksik iyonların (Na+ ve Cl-) birikmesi veya K+ ve Ca+2 iyonlarını tüketmesi biçiminde ortaya çıkmaktadır (Yeo ve ark., 1993). NaCl, bitki hücre duvarının sertleĢmesine (Neumann, 1993; Neumann ve ark., 1994; Nabil ve Coudret, 1995) ve plazma membranının su taĢıma yeteneğinin düĢmesine neden olur (Azaizeh ve ark., 1992; Cramer, 1992). Potasyumun fotosentez, enzimatik aktivite, turgor potansiyeli, hücre uzaması, toprak üstü ve toprak altı organlarının büyümesi, stoma hareketliliği, transpirasyon ve protein sentezinde önemli etkileri vardır (Tisdale ve ark., 1993; Marschner, 1995).
Çimlenmenin azalmasının ana nedeni olarak, yüksek tuz yoğunluğundan ötürü su alımının engellendiğini açıklayanlar olduğu gibi (Mansour, 1994), tuzun toksik etkisinden ortaya çıktığını belirtenler de vardır (Leopold ve Willing, 1984; Khajeh-Hosseini ve ark., 2003). Tuzluluk problemi olan topraklarda Cl- ve SO4- tuzları dominant olarak bulunmakta ve doğadaki tuzluluk stresi genelde Na+ tuzları ve NaCl tarafından meydana getirilmektedir (Özdemir ve Engin, 1994; Özcan ve ark., 2000).
Ayrıca bitkilerin tuza karĢı gösterdikleri tepki, geliĢme durumlarına göre farklılık gösterdiği gibi, bitki familyalarının ve hatta tür içindeki çeĢitlerin de tuzluluğa farklı reaksiyon gösterdiği bilinmektedir. Tuzlu koĢullarda çimlenme ve fide geliĢimi dönemi, bitkinin toplam yaĢam döngüsü içerisinde en kritik dönemdir (Katerji ve ark., 1994; Wang ve Shannon, 1999; Almansouri ve ark., 2001).
Marschner (1995)’a göre, bir bitkinin tuza dayanım gösterebilmesi için osmotik uyum mekanizmalarından bir veya birkaçı olması gerekmektedir. Tuz stresi ile karĢılaĢan bitkilerde; dıĢarıdan tuz iyonlarının bünyeye alınması veya bünye tarafından çözünebilir organik maddelerin sentezlenmesi ve bunların hücre içinde biriktirilmesi yoluyla osmotik uyum sağlanabilmektedir.
Lutts ve ark. (1996)’nın çeltik bitkisinde yaptıkları bir araĢtırmada tuza dayanıklı çeĢitte MDA miktarı en düĢük değerleri verdiği halde, tuza duyarlı çeĢitte en yüksek MDA ölçümleri yapılmıĢtır. Tuz stresi altında tolerant ve duyarlı genotipler arasındaki çok çeĢitli fizyolojik ve biyokimyasal değiĢimlerin incelenmesi sayesinde, geniĢ bir gen havuzunda tuz stresine tolerans bakımından yapılacak bir tarama (screening) çalıĢmasında kullanılabilecek etkin parametrelerin ortaya konması mümkün olabilecektir.
Kültür bitkilerinde tuz ve su stresi koĢullarında stresin ilk adımlarından itibaren oldukça yüksek değerlere çıkan parametrelerden birisi prolindir. Bir aminoasit olan prolin, oldukça uzun zamandan bu yana araĢtırılmaktadır ve stres altındaki bitkilerde hücre ve dokuların en azından ilk zamanlarda zarar görmesini önlemek amacıyla bitki tarafından içsel olarak salgılanan bir organik bileĢiktir (Hare ve Cress 1997). Stres koĢulları sona erdiğinde kendiliğinden ve geriye bir hasar bırakmaksızın normal değerlere dönmektedir.
Tuz stresinde yetiĢtirilen buğday (GüneĢ ve ark., 1997) ve mısır (Taban ve ark., 1999) çeĢitlerinden tuza dayanıklı olan çeĢitlerin sodyum ve klor konsantrasyonlarının düĢük, potasyum ve prolin konsantrasyonlarının ise daha yüksek olduğunu saptamıĢlardır. NaCl uygulamasına bağlı olarak mısır bitkisinin Na ve Cl konsantrasyonları artarken, K konsantrasyonu azalmıĢtır (Katkat ve ark., 1999).
Bitkilerin tuza dayanıklılıklarının kök sistemleriyle aĢırı Na ve K alımını engellemelerine bağlı olduğu bildirilmiĢtir (GüneĢ ve ark., 1997). Yine yapılan baĢka araĢtırmada aĢırı miktarda Na ve Cl absorbsiyonunun, iyon dengesinde K aleyhine meydana getirdiği bozulmanın sebep olduğunu söylemek mümkündür. Na genellikle K alımını engellemekte, Cl ise özellikle NO3 üzerine olumsuz etki yaparak bitkilerde iyon dengesinde bozulmalara sebep olmaktadır (Alparslan ve ark., 1998).
Pamuk tohumlarının çimlenme ve çıkıĢı, aĢırı tuz nedeniyle hem azalmakta hem de gecikmektedir Genellikle tuzluluk pamuğun vejetatif geliĢimini azaltır (Qadir ve Shams, 1997). DüĢük tuz yoğunlukları ise büyümeyi arttırabilir (Pessarakli ve Tucker,
1985; Gorham, 1996). Prolinin normal biyokimyasal reaksiyonları engellemediği ve osmotik stres sırasında osmo-protektant (osmo-koruyucu) olarak rol oynadığı düĢünülmektedir (Yoshiba ve ark., 1997).
Strese bağlı olarak fotosentez II’ de meydana gelen değiĢikler nedeniyle fotosentez II’ nin aktivitesi azalmaktadır. Tuzlu koĢullarda bitkilerin klorofil miktarında azalma meydana geldiğini ve fotosentez II’nin fotosentez I’den daha olumsuz etkilendiğini rapor etmiĢlerdir. Tuz stresi altında büyütülen mısır fidelerinde toplam klorofil, klorofil-a ve klorofil-b içeriği önemli ölçüde azalmaktadır Aynı araĢtırmada yeĢermenin ilk 12 saatinde artıĢ gösteren klorofil a/b oranı 48 saat içinde önemli ölçüde düĢüĢ göstermiĢtir. Tuz stresi altında fotosistem-I in fotosistem-II ye göre daha stabil kaldığı göstermiĢlerdir. AraĢtırıcılar tuzun olası zararlarını ıĢık toplama kompleksinin ve tilakoid membranların azalmasına ve fotosistemler arasındaki koordinasyonun bozulmasına bağlamıĢlardır. (Ganieva ve ark., 1997)
Prolinin stres koĢullarında arttığı, serbest O2 radikallerinin detoksifikasyonuna katıldığı ve stres koĢullarına dayanıklılıkta önemli rol oynayan koruyucu özelliğe sahip azot içerikli bir bileĢik olduğu bilinmektedir (Bohnert ve Sheveleva, 1998).
Tuzun, hücre geniĢlemesinin daha yetersiz olmasına neden olabileceği (Volkmar ve ark., 1998) ve DNA sentezine zarar verebileceği de belirtilmiĢtir (Waisel, 1972). Tuzlu ortamlarda yetiĢen bitkilerde, su stresi, aĢırı Na ve Cl alımı ile iliĢkili iyon toksitesi ve besin maddeleri alımı ve yeĢil organlara taĢınmasında stres nedeniyle oluĢan dengesizlik ile hücre içi mineral bileĢiminin, özellikle K ve kısmen Ca dengesinin bozulması sonucu büyüme sınırlanmaktadır (Karanlık ve ark., 1999).
Tuzlu Ģartlarda yetiĢtirilen buğdayda tuza dayanıklılık kriteri olarak bitkinin Na+ ve K+ içeriği kullanılmaktadır (Ahsan ve Khalid, 1999). Tuzluluğun, epidermis hücre sayısı (Martıns ve Castro, 1999), epidermis hücre geniĢliği (Curtıs ve Lauchlı, 1987), stoma sayısı (Flowers ve ark., 1986), stoma değeri (Bray ve Reid, 2002), yaprak kalınlığı (Yeo ve ark., 1991), iletim demetleri arası mesafe (Kılıç ve ark., 2007), stoma geniĢliği ve uzunluğu (ÇavuĢoğlu ve ark., 2007) üzerinde engelleyici etki yaptığı uzun zamandan beri bilinmektedir.
Na miktarındaki artıĢ, genellikle osmotik regülasyonu ve besin dengesini bozarak spesifik iyon toksitesine neden olmaktadır. Bu durum, yetiĢtirme ortamının iyonik dengesinden ziyade net Na oranına bağlı olup, bitkilerin tuza toleransını belirlemektedir. Artan Na içeriği genellikle K miktarında azalmaya neden olmakta, bu
durum Na ile K arasındaki antagonizm nedeniyle desteklenmektedir. Benzer Ģekilde, dıĢ ortamda bulunan yüksek seviyedeki Na oranları, toprak çözeltisindeki Ca’un aktivitesinde büyük oranda azalmaya neden olmakta, kök hücrelerinin plazmalemmalarından Ca’u çıkartarak onun yerine geçebilmektedir (Alam,1999; Ghoulam ve ark., 2002; Çiçek ve Çakırlar, 2002).
Osmotik stres ve iyon toksisitesi tuz stresinden kaynaklanan problemlerdir ve kimyasal aktivitede meydana gelen azalma, hücrelerde turgor kaybına neden olur. Hücre büyümesi, hücre duvarları gerilimi için turgora ihtiyaç duyar ve turgorun olmayıĢı hücrenin yaĢamını sürdürmesi için tehlike oluĢturur (Proft ve Serrano, 1999).
Kültürü yapılan bitkiler arasında tuza karĢı tolerans bakımından farklılıklar bulunmaktadır. Bunun yanısıra aynı tür içerisinde çeĢitler arasında tuza tolerans düzeyindeki farklılıklar birçok çalıĢma ile belirlenmiĢtir. Domates bitkisinde çeĢitlerin tuza farklı dayanım gösterdiklerini belirleyen Alian ve ark. (2000), Fireball adındaki bir çeĢidin tuzlu koĢullarda bünyesine fazla miktarda Na+ iyonu aldığı halde bundan olumsuz yönde etkilenmediğini; buna karĢılık aynı oranda Na+ iyonunu bünyesine alan Patio adlı domates çeĢidinin tuz toksisitesi belirtilerini gösterdiğini kaydetmektedir.
Tuz stresi altındaki bitkilerde stomalar kapatılmakta, yaprak alanları da küçültülerek transpirasyon azaltılmaya çalıĢılmaktadır. Böylece bitki, su kaybını en aza indirmek ve topraktan su ile birlikte yüksek miktardaki tuzu almayı engellemeye gayret etmektedir. Yaprak alanındaki azalmanın yanında birim alandaki CO2 fiksasyonu da azalmaktadır. Bütün bunlara, yükselen respirasyon eĢlik eder. YaĢamak için yoğun enerji sarfeden bitki, daha az fotosentez yaparak harcadıklarını yerine koyamadığı için geliĢme ve büyüme geriler. Tuz stresi altında net CO2 fiksasyonunun azalması; su noksanlığı, stomaların kapanıĢı, apoplastta tuzun birikmesi ve mezofil hücrelerinin turgoru kaybetmesi veya tuz iyonlarının doğrudan toksisitesi nedeniyledir (Karanlık, 2000).
Mer ve ark. (2000), tuzun toksik etkisinin ilk önce yaĢlı yapraklarda görülmeye baĢladığını ve toksisitenin bu yaprakların uçlarından baĢlayıp yaprak ayasına ve sapına doğru ilerleyen kloroz Ģeklinde kendini gösterdiğini, daha sonra bu kısımların nekroze olduğunu belirtmektedir. Tuzlu koĢullarda büyüyen bitkilerin büyüme hızı düĢük olup bodur bir yapı sergilemektedirler, yaprakları ise çoğunlukla küçük ve rengi de koyudur. Tuz stresinde hücre büyümesi ve bölünmesinde yavaĢlamanın, sitokinin miktarının
azalması sonucu ortaya çıktığı ileri sürülmektedir. Tuz stresi bitkilerde Na birikimini arttırmakta, K alımını ise azaltmaktadır (Qian ve ark., 2000).
Reid ve Smith (2000)’nin yaptığı çalıĢmada 150 mM NaCl uygulamasının buğday (Triticum aestivum L.) bitkisinin büyümesini Ģiddetli bir Ģekilde azalttığını, geliĢme ortamında Ca konsantrasyonunun 2.34 mM üzerine çıkarılması ile büyümenin normale döndüğünü belirtmiĢlerdir. Ayrıca Ca tuz stresi altında K alım ve transportunu artırma yoluyla Na akümülasyonunu azaltır.
Zhu (2001), tuzluluğun stomaların kapanması dolayısıyla transpirasyonun ve kloroplastlara net CO2 diffüzyonunun azalmasına yol açarak fotosentezin olumsuz Ģekilde etkilendiğini rapor etmiĢtir. Bu durumun da beraberinde oksidatif zararlanmayı getirdiğini belirtmiĢtir.
Din ve ark. (2001) K+/Na+ oranının tuzluluğa bağlı olarak düĢtüğünü, tuza maruz bırakılan bitkilere yapraktan ve topraktan uygulanan potasyumun ise K+/Na+ oranını yükselttiğini bildirmektedirler. Helal ve ark. (1975) ile Shaaban ve ark. (2004)’da benzer sonuçları bildirmiĢlerdir. Tuzlu alanlarda toprağa uygulanan potasyum, arpada tuzun negatif etkisini azot alınımını arttırarak azaltmaktadır (Helal ve ark., 1975).
Karanlık (2001) ile YaĢar (2003)’a göre tuz stresi altındaki bitkilerde stomalar kapatılmakta, yaprak alanları da küçülerek transpirasyon azaltılmaya çalıĢılmaktadır. Böylece bitki, su kaybını en aza indirmek ve topraktan su ile birlikte yüksek miktardaki tuzu almayı engellemeye gayret etmektedir. Yaprak alanındaki azalmanın yanında birim alandaki CO2 fiksasyonu da azalmaktadır. Bütün bunlara, yükselen respirasyon eĢlik eder. YaĢamak için yoğun enerji sarf eden bitki, daha az fotosentez yaparak harcadıklarını yerine koyamadığı için geliĢme ve büyüme geriler. Tuz stresi altında net CO2 fiksasyonunun azalması; su noksanlığı, stomaların kapanıĢı, apoplastta tuzun birikmesi ve mezofil hücrelerinin turgoru kaybetmesi veya tuz iyonlarının doğrudan toksisitesi sebebidir.
Fischer (2001), yaprakların klorofil içeriklerinin onların fotosentetik kapasitelerini yansıttığını,yaprağın yeĢilliğini (klorofil içeriği) ve azot kapsamını tespit etmede SPAD metre kullanımının pahalı olmayan, hızlı ve yaprak yeĢilliğine zarar vermeyen bir yöntem olduğunu, en uygun okuma zamanının ise klorofilin en iyi düzeyde olduğu çiçeklenme sonrasında olduğunu ve Yadava (1986), SPAD değerleri ile okuma anında yaprakların içerdiği klorofil miktarları arasında linear bir iliĢkinin
bulunduğunu bildirmiĢtir. Bitkilerde fotosentetik organlar sıcaklığa hassas olup; genellikle yüksek sıcaklık semptomlarının görünür hale gelmesinden önce oldukça fazla zarar görmektedirler (Crafts-Brandner ve Salvucci, 2002; Camejo ve ark., 2005).
Tuzluluk, uzun yıllardan beri üzerinde çalıĢılan ve çözüm aranılan, bitkilerin geliĢimleri ile üretimlerinde verim ve kaliteyi düĢüren en önemli çevresel etkendir. YetiĢtirme ortamının osmotik basıncının yükselmesi, suyun ortamda bulunmasına rağmen alınamamasına sebep olmaktadır (Katerji ve ark., 2001; Romero ve ark., 2001).
Yi Hui-Ian (2001)’de yapmıĢ olduğu; “Tuz stresi altında çimlendirilen Hordeum vulgare’de hücre bölünmesi ve kromozom davranıĢları” adlı çalıĢmasında tuz çözeltisinin kardeĢ kromatit değiĢimi ve kromozom davranıĢları üzerine etkilerini araĢtırmıĢ ve mikronükleus, kromozomlarda fragment oluĢumu, anafazda köprü oluĢumu ve düzensiz anafaz gibi kromozom anormalliklerinin olduğunu tespit etmiĢtir. Aynı araĢtırıcı NaCl’ün, konsantrasyon ve uygulama süresine bağlı olarak mitotik değeri etkilediğini de vurgulamıĢtır.
Ortam tuzluluğun artması ile bitkilerin almıĢ oldukları kalsiyum miktarındaki azalma varılan diğer bir sonuçtur. Biber bitkisinde yapılan bir çalıĢmada, ortamdaki tuz miktarının artmasıyla bitkinin almıĢ olduğu kalsiyum miktarında düĢme, verimde azalma olduğu bildirilmektedir (AktaĢ, 2002).
Matysik ve ark. (2002), prolinin genellikle stres koĢullarında birikimi gerçekleĢen, bitkinin dayanım yeteneğini sağlaması bakımından bir indikatör görevini yapan (Bian ve ark., 1988; Sairam ve ark., 2002), suda çözünebilir bir aminoasit olduğunu belirtmiĢlerdir. Osmolit olarak görev yapmasının yanında, hücrelerin stabilizasyonu, sitosolik pH’nın ayarlanması ve hidroksil radikallerinin düzenlenmesinde etkili bir organik bileĢik olduğunu ifade etmiĢlerdir.
Topraktaki tuzlar, suyun osmotik basıncını yükselterek tohumlar tarafından alınmasını engellemekte veya Na+ ve Cl- iyonlarının toksik etkisinden dolayı çimlenmeyi olumsuz etkilemektedir (Öz ve Karasu, 2002; Sadeghian ve Yavari, 2004; Essa, 2002).
Tuzluluğun, tohum çimlenmesini engellediği (Kabar ve Baltepe, 1987; Gulzar ve Khan, 2002), kök ve gövde uzamasını baskıladığı (Dash ve Panda, 2001; Ashraf ve ark., 2002), taze ağırlık ve su içeriğini azalttığı (El-Mashad ve Kamel, 2001) birçok araĢtırıcı tarafından ortaya konmuĢtur. Na etkisine maruz bırakılmıĢ kültür bitkilerine dıĢarıdan
takviye olarak verilen 5-20 mM katyonik elementlerinin bitkide strese toleransı arttırdığını bildirmektedir (Cramer 2002).
Tuz stresi altında kalan mısır bitkisinde genel geliĢme bozuklukları ve diğer parametrelerde etkilenme sıklıkla bildirilmektedir. Çiçek ve Çakırlar (2002) çalıĢmalarında tuz stresine maruz bırakılan mısır bitkisinde, bitki boyu, nispi su içeriği ile toplam yaĢ ve kuru ağırlıklarda azalma saptarken, prolin, Na ve Na/K oranlarında artma rapor etmiĢlerdir.
Irshad ve ark (2002), tarafından yürütülen çalıĢmalar sonucunda tuz stresi altındaki bitkilerde köklerin su alma yeteneklerinde önemli azalmalar meydana geldiğinden, kök geliĢimi ve gövde uzaması gibi faaliyetlerde gerileme görülür. Stres altındaki bitkilerin gövde çapları azaldığı gibi boyları da kontrole göre küçük kalmaktadır. Aynı Ģekilde yaprak alanı ve generatif evreye geçiĢte çiçeklenme ve meyve verimi de olumsuz etkilenir. Tuz stresinin yukarıda sayılan sonuçları uzun dönemde ortaya çıkan arazlardır.
Stres altındaki bitkilerin sürgün ve köklerinde kuru madde ve yaĢ ağırlıklarında önemli ölçüde azalmalar olduğu birçok bitkide baĢka araĢtırıcılar tarafından rapor edilmiĢtir (Irshad ve ark., 2002; Gholulam ve ark., 2002; DaĢgan ve ark., 2002). Tuz, tohumun çimlenmesinde önemli rol oynayan bazı enzimlerin aktivitelerini engelleyebilir Tuz stresi pamukta yaprak alanını, kök ve toprak üstü kısımlarının geliĢimini düĢürür (Saghir ve ark., 2002).
Kafi ve ark. (2003), farklı NaCl deriĢimleri (0, 100, 200 ve 300 mM) etkisinde bıraktıkları tuza tolerant ve tuza hassas buğday çeĢitlerinde prolin ve karbonhidrat miktarlarını araĢtırmıĢlardır. Prolin ve karbonhidrat miktarlarının buğday çeĢitleri ve bitkilerin olgunlaĢmıĢ tohum, yapraklar kökler arasında dikkate değer oranlarda farklılıklar gösterdiğini bulmuĢlardır. 200 mM’a kadar tuz uygulaması bütün buğday çeĢitlerinde toplam karbonhidrat miktarlarında artıĢa neden olduğunu saptamıĢlardır.
Yüksek tuz konsantrasyonlarında iyon birikimi ve stomaların açılıp kapanmasındaki düzensizlikler nedeniyle toplam klorofil miktarında azalmalar meydana gelmekte, bunun sonucu olarak fotosentez etkinliği azalarak bitkinin geliĢiminde olumsuzluklar çıkmaktadır (YaĢar, 2003)
Tuzların hücre duvarı ve membranların fonksiyonunu etkilediği NaCl ile yapılan deneylerden bilinmektedir (Tobe ve ark., 2004). Tuzların, hücre duvarı ve hücre membranı üzerindeki bu etkilerinin, sitosolün su potansiyelini, hücresel geniĢlemesini
ve dolayısıyla tohumun çimlenmesini etkileyebileceği belirtilmektedir (Tobe ve ark., 2004).
Ashraf ve ark. (2004), Pakistan’da 2001-2002 yetiĢme sezonunda, 25 buğday genotopi ve iki tuz dozu(0, 150 mol/m3) kullanarak bu genotiplerin tuzluluğa etkilerini incelemiĢlerdir. Tuz konsantrasyonunun kuru sap ağırlığının kontrole göre %26.4 - %69.6 oranında azalttığını bildirmiĢlerdir. Kullandıkları genotiplerden Na(20) TPP, Penjamo 62 ve Inia 66’nın en yüksek verimi verdiklerini açıklamıĢlardır. Adıyaman (2005), ġahin 91 arpa ve Ceylan 95 makarnalık buğday çeĢitlerinin farklı tuz dozlarına ( 0, 2, 4, 6, 8, 10, 12, 14, 16 dS/m ) tepkilerini belirlemek amacıyla bir çalıĢma yürütülmüĢtür. Çimlenme ve geliĢme dönemlerinde farklı tuz dozlarının etkisinde kalan arpa fidelerinin yapılan korelasyon analizi sonucunda tuz dozlarının, yeĢil aksam kuru ağırlığı ve kök kuru ağırlığı üzerine etkisinin P≤0.01 seviyesinde önemli olduğu ve tuz dozu uygulamaları ile parametreler arasında negatif bir iliĢki olduğu belirlenmiĢtir. Tuz dozu artıĢı bitki geliĢimini olumsuz yönde etkileyerek ağırlık kayıplarına neden olmuĢtur.
Azevedo Neto ve ark. (2004) tarafından mısır bitkisi kullanılarak yapılan bir çalıĢmada, tuz stresi ile iliĢkili olarak yaprak ve köklerin Na içeriği arttıkça potasyum (K) içeriğinin düĢtüğü, yaprak su potansiyeli ve transpirasyon yeteneğinin özellikle tuza hassas çeĢitte bozulduğu bildirilmiĢtir.
Jaradat ve ark. (2004) , dört ( 0, 0.85, 10 20 dS/m) farklı tuz konsantrasyonu kullanarak tuzluluğun, arpanın çimlenme ve fide geliĢimine etkilerini incelemek için yaptıkları çalıĢmada, artan tuz konsantrasyonunun fidelerin yaĢ ve kuru ağırlığını azalttığını bildirmiĢlerdir.
Leonova ve ark. (2005), fide uzunluğu temelinde 12 arpa çeĢidini, Bağcı ve ark. (2003) ise kuru ağırlık temelinde 8 arpa çeĢidini tuzluluk toleransı açısından sınıflandırmıĢlardır. Leonova ve ark. (2005), tuza toleranslı ve hassas ikiĢer çeĢitte, Bağcı ve ark. (2003) ise tüm çeĢitlerde Na+ ve K+ analizleri yaparak arpa çeĢitlerinin tuza toleranslarındaki karakteristikleri belirlemiĢlerdir.
Tuzlu Ģartlarda yetiĢtirilen bitkilerde verim kaybını önlemek ve bozulan osmo regülasyonu yeniden sağlamak temel hedef olmalıdır. Tuz stresine maruz kalmıĢ bitkilerde verimdeki azalıĢın diğer bir nedeni de iyon dengesindeki bozulmadır. Bitki dokularında element bağlama yerlerinde Na ile özellikle K ve Ca gibi diğer katyonik elementlerin rekabete girmesinden dolayı Na/K ve Na/Ca dengesi hızla bozulmakta ve
bu elementlerin alınımı ve taĢınımı azalmaktadır. Bir çeĢit K noksanlığı yaratan bu durum, bitkide osmo regülasyonu bozmakta, enzimlerin aktivasyonunu engellemekte ve metabolizma olumsuz etkilenmektedir. Bu gibi durumlarda dıĢarıdan potasyum takviyesi acil ve gerekli bir koĢuldur. Bu koĢul yerine getirildiğinde bitkinin stresten etkilenme derecesi azalmaktadır (Tuna ve Kaya, 2005).
Demiral (2007), Arabidopsis thaliana’nın yabani tipi Columbia ve brassinosteroid sentezleyemeyen det2 mutantında, 24-epibrassinolid (24-epiBL)’in tuz stresi altında fotosentetik verim değerleri üzerindeki etkisi araĢtırılmıĢtır. Arabidopsis thaliana BR-yoksun mutantı det2 ve yabani ekotipi Columbia’nın fotosentetik veriminde (Fv/Fm) meydana gelen degisimler incelenmiĢtir. 4 gün boyunca 200 mM NaCl uygulaması det2’nin fotosentetik veriminde % 3, Columbia’nınkinde ise % 2 azalmaya neden olmuĢtur. Dıstan 24-epiBL uygulaması det2’nin fotosentetik verimini kontrol grubuna göre % 4 artırmıs olmasına ragmen, Columbia’nın fotosentetik veriminde önemli bir degisim meydana getirmemiĢtir. 24-epiBL uygulaması det2’nin fotosentetik veriminde tuz stresinin neden olduğu azalmayı önleyerek fotosenetik verimi % 4 oranında arttırırken, Columbia’da oluĢan azalmayı iyileĢtirememiĢtir.
Karakullukçu ve ark. (2008) tarafından yapılan bir çalıĢmada bazı nohut çeĢitlerinin toprak üstü yaĢ ve kuru ağırlık ortalamaları incelendiğinde, yaĢ ağırlık bakımından çeĢitler, kuru ağırlık bakımından ise çeĢitler X tuz interaksiyonu önemlidir. Bu çeĢitlerin tuza karĢı tepkilerinin farklı olduğunu göstermektedir. Hem yaĢ hem de kuru ağırlık ortalamalarında tuz uygulamasında daha düĢük değerler elde edilmiĢtir. Bazı nohut (Cicer arietinum L.) çeĢitlerinden elde edilen sonuçlara göre Na içeriği bakımından bitkinin toprak üstü aksamı ve kökte tuz uygulamasında daha yüksek sonuçlar alınmıĢtır.
Seçkin (2010)’ nun yaptığı çalıĢması da bizim çalıĢmamızı desteklemiĢtir. Deniz arpası (Hordeum marinum) ve kültür arpası olan (Hordeum vulgare)’deki tuz stresinin uyardığı oksidatif hasara karĢı tolerans mekanizmaları karĢılaĢtırmalı olarak araĢtırıldı. 7 gün boyunca 0, 150 ve 300 mM NaCl stresine maruz kalan H. marinum ve H. vulgare cv. Tokak 157/37 türlerindeki stoma iletkenliği değiĢimi tespit edildi. Uygulanan tuz konsantrasyonunun artmasına bağlı olarak her iki türde de stoma iletkenliği azalmıĢtır. 150 mM NaCl, H. marinum ve H. vulgare’de sırasıyla % 27 ve % 18 azalmaya neden olmuĢtur. 300 mM NaCl ise H. vulgare’de % 59 H. marinum’da ise % 39’luk bir azalmayı sonuçlandırmıĢtır.
Yıldız ve Terzi, (2011) ’nin yaptığı çalıĢmada farklı NaCl konsantrasyonlarına (0, 100, 150 ve 200 mM) maruz bırakılan on iki arpa (Hordeum vulgare L.) çeĢidi erken fide evresinde tuza toleransları bakımından sınıflandırılmıĢtır. Arpa çeĢitlerinin kök ve gövde kuru ağırlıkları üzerine çeĢit ve NaCl tuzluluğu ana faktör etkileri ve bu faktörlere ait etkileĢimler (çesit×NaCl) önemli düzeyde bulunmuĢtur. Arpa çeĢitlerinin kök ve gövde kuru ağırlıkları, NaCl konsantrasyonundaki (100, 150 ve 200 mM) artıĢa bağlı olarak önemli düzeyde (P<0.001) farklılıklar göstermiĢtir. Sonuç olarak, Avcı-2002 tuza en toleranslı ve Tokak 157/37 tuza en hassas arpa çeĢidi olarak belirlenmiĢtir. Na+/K+ oranının tuza tolerans açısından etkili bir kriter olarak kullanılıp kullanılmayacağını değerlendirmek için Avcı-2002 ve Tokak 157/37 arpa çeĢitleri, kontrol veya artan NaCl konsantrasyonlarına (100, 200 ve 300 mM) 6 gün için maruz bırakılmıĢtır. Her iki arpa çeĢidi, köküne göre gövdesinde daha fazla Na+ ve K+ biriktirmiĢtir. Her iki arpa çeĢidinin kök ve gövde dokusunda NaCl konsantrasyonunun artmasıyla Na+ içeriği önemli düzeyde artarken, K+ içeriği önemli düzeyde azalmıĢtır (P<0.05).
3. MATERYAL VE METOT 3.1. Materyal
AraĢtırmada materyal olarak çeĢit adayı olan, S.Ü. Ziraat Fakültesi Öğretim Üyelerinden Prof. Dr. Bayram Sade ve Prof. Dr. Süleyman SOYLU tarafından geliĢtirilen 2 arpa hattı ve 4 tescilli arpa çeĢidi Konevi, Larende, Anadolu 98 (tuza dayanıklı) ve Tokak 157/37 (tuza hassas)] olmak üzere toplam 6 genotip kullanılmıĢtır (Çizelge 3.1). Bu tezde Anadolu 98 bundan sonra kısaca Anadolu, Tokak 157/37 ise kısaca Tokak olarak adlandırılacaklardır.
Bu araĢtırma, Selçuk Üniversitesi Alaaddin Keykubat Kampüsü’nde bulunan Ziraat Fakültesi Tarla Bitkileri Biyoteknoloji Laboratuarı ile Fizyoloji Laboratuvarlarında yürütülmüĢtür.
Çizelge 3.1. AraĢtırmada kullanılan arpa genotiplerinin genel özellikleri
GENOTĠPLER ÖZELLĠKLER
Anadolu 98 Ġki sıralı, beyaz taneli ve maltlık kalitesi yüksektir. KıĢa, kurağa ve tuza
dayanıklıdır.
Konevi Ġki sıralı ve yatmaya meyillidir. Kurağa ve soğuğa orta dayanıklıdır. Larende Ġki sıralı, orta boylu ve yatmaya dayanıklıdır.
Tokak 157/37 Ġki sıralı ve tuza hassastır. Maltlık kalitesi iyi olup, adaptasyon kabiliyeti çok
geniĢtir.
Soylu-10 Ġki sıralı, orta boylu ve kurağa dayanıklıdır.
Soylu-11 Ġki sıralı, beyaz taneli ve orta boyludur. KıĢa ve kuraklığa dayanıklıdır.
3.2. Metot
3.2.1. Denemelerin kurulması 3.2.1.1. Saksıların hazırlanması
Tohumların ekimi için kullanılacak saksılar önce çeĢme suyundan sonra saf sudan geçirilmiĢtir. Saf sudan geçirilen saksıların içlerine saksıların tamamını dolduracak kadar yıkanmıĢ perlit konulmuĢtur. AraĢtırmada altı arpa genotipi, 4 muamele ve 3 tekerrür olmak üzere (6*4*3) toplam 72 adet 2 kg’lık beyaz saksılar kullanılmıĢtır.
3.2.1.2.Tohumların hazırlanması
Tohumlar ekilmeden önce %5’lik çamaĢır suyu ile 5 dk sterilize edilmiĢtir. Her saksı için 12 adet tohum belirlenmiĢtir.
3.2.1.3. Besin çözeltisinin hazırlanması
Besin çözeltisi olarak Half-Hoagland çözeltisi tuz (NaCl) bakımından modifiye edilerek hazırlanmıĢtır. Ele alınan uygulamalar aĢağıdaki gibi hazırlanmıĢtır;
(1) Half hoaglant çözeltisi (Kontrol) (2) Half hoaglant çözeltisi + 25 mM NaCl (3) Half hoaglant çözeltisi + 50 mM NaCl (4) Half hoaglant çözeltisi + 75 mM NaCl
Hazırlanan besin cözeltisi, tohumlar çıkıĢ yaptıktan sonra 2 günde bir olmak üzere (100 ml) saksılara uygulandı.
3.2.1.4. Ekim ve kültür Ģartları
Sterilize edilen tohumlar her saksıya 12 adet olmak üzere eĢit aralıklı ekilmiĢtir. ÇıkıĢtan sonra tüm serilerde sulama besin çözeltisi ile iki günde bir ve eĢit miktarda (100 ml) olmak üzere, kontrol grubuna sadece Half Hoagland besin çözeltisi, diğer serilere ise 25, 50 ve 75 mM NaCl içeren Half Hoagland çözeltisi filizlenmeden sonra 10 hafta süre ile uygulanmıĢtır.
Bitkiler iklim odasında kontrollü Ģartlarda yetiĢtirilmiĢtir. Gerekli ısı, nem, ıĢık ve sterilizasyon kontrolleri yapılmıĢtır. Ġklim odasında, bitkilerin tohum çimlenmesi ve çimlenme sonrası büyüme ve geliĢme süresince 24±2 ºC, 16/8 fotoperiyot, %65 nem, 10000 LuX/Gün ıĢık intensitesi olacak Ģekilde ayarlanmıĢtır. Tüm denemeler “Tesadüf Parselleri Deneme Desenine ’’ göre 3 tekerrürlü olarak planlanmıĢtır.
3.2.2. Gözlem ve ölçümler
3.2.2.1. Büyüme parametrelerinin ölçülmesi
Onuncu hafta sonunda bitkinin GS-(25-30) döneminde büyüme parametrelerinin ölçümleri yapılmıĢtır.
3.2.2.1.1. Uzunluk (cm)
Her saksıdan tesadüfi beĢ arpa bitkisinin toprak üstü ve toprak altı aksamı ölçülerek kök ve bitki uzunlukları cm cinsinden tespit edilmiĢtir.
ġekil 3.1. Bitkilerin hasat sonrası bitki uzunluğu ölçümü
3.2.2.1.2. YaĢ ağırlık (g)
Her saksıdan tesadüfi beĢ bitki örneği hasat sonrası kök ve bitki kısımları birbirinden ayırıp ayrı ayrı hassas terazide tartılarak gram cinsinden belirlenmiĢtir.
3.2.2.1.3. Kuru ağırlık (g)
YaĢ ağırlığı tespit edilen bitki örnekleri kök ve bitki kısımları ayrı olmak üzere etüvde 70°C’ de 48 saat kurutularak kuru ağırlıkları gram cinsinden belirlenmiĢtir.
3.2.2.2. Fizyolojik özellikler
3.2.2.2.1. Bağıl su içeriği (RWC) (%)
Onuncu hafta sonunda seçilen bitkilerden yaprakların orta kısımlarından 5 adet yaprak örneği alınarak (5 ayrı bitki örneğinden) yaĢ ağırlıkları ölçülmüĢtür. Bu yapraklar 6 saat boyunca saf su içinde petri kaplarında bekletilerek turgor haline gelmeleri sağlanmıĢ ve turgor ağırlıkları ölçülmüĢtür. 70 0
C’ de 72 saat etüvde kurutulduktan sonra kuru ağırlıklar saptanmıĢtır. Her bir gruba ait yaprak örneklerinin nispi su içeriği aĢağıdaki formüle göre % olarak hesaplanmıĢtır (Barr ve Weatherley 1962);
Bağıl Su Ġçeriği (%) = [(YA – KA) / (TA – KA)]X100 YA=YaĢ Ağırlık
KA=Kuru Ağırlık TA=Turgor Ağırlığı
3.2.2.2.2. Klorofil değeri (spad)
Bitki örneklerinin klorofil içerikleri 5 ayrı bitkide 3 tekerrürlü olarak Spadmetre (Spad–502) cihazı ile ölçülmüĢtür.
Bu sistem yaprakların sahip olduğu yeĢil rengin ölçülmesi yolu ile dolaylı olarak klorofil miktarı ve azot içeriğinin belirlenmesi esasına dayanmaktadır (Fischer 2001).
ġekil 3.4. AraĢtırmada çeĢitlerin klorofil miktarı özelliğinin tespit çalıĢmasından görüntüler
3.2.2.2.3. Stoma iletkenliği (mmol/m2sn)
Stoma iletkenliği her saksıdan 5 bitkide 3 tekerrürlü olarak porometre cihazı ile ölçülmüĢtür.
3.2.2.2.4. Fotosentetik verim (klorofil flüoresansı) (Fv/Fm)
Ölçümler, 5 ayrı bitkinin her birinde ve 4 tekerrürlü olarak kapalı havada, hasat öncesi yapılmıĢtır. Ölçümlerden önce yaprakların orta kısımlarının üst yüzeyleri 30 dk boyunca kapatılmıĢtır. Bitki verim analiz cihazı “Plant Efficiency Analyser” (PEA) (Hansatech Instruments Ltd.) flüometresi ile; DeğiĢken flüoresans / Maksimum flüoresans indüksiyonu (Fv/Fm) oranları belirlenmiĢtir.
3.2.2.2.5. Prolin analizi (µmol/g)
Onuncu hafta sonunda serbest prolin içeriğinin belirlenmesi için, her çeĢitten 4 tekerrürlü olarak yaprakların orta kısımlarından örnekler alınmıĢ ve analiz Bates ve ark. (1973)’na göre yapılmıĢtır. Sıvı fazdan aspire edilen toluen fraksiyonunun 520 nm'deki absorbansı spektrofotometreden okunmuĢtur. Prolin konsantrasyonu, kalibrasyon eğrisi kullanılarak hesaplanmıĢ ve µmol prolin g-1
taze ağırlık olarak ifade edilmiĢtir. 3.2.2.3. Element analizi (Cl, Na, K, Ca)
Onuncu hafta sonunda her genotipe ait bitkilerin toprak üstü aksamı hasat edilmiĢtir. Hasat sonrası kurutulan örnekler alınarak element analizleri yapılmıĢtır. 3.2.2.3.1. Klor tayini (Cl)
Onuncu hafta sonunda klor tayini (Cl), öğütülmüĢ ve kurutulmuĢ bitki örneklerinden 0.1 g tartılıp, 50 ml kapasiteli santrifüj tüpüne konularak üzerine 25 ml saf su ilâve edilmiĢtir. Santrifuj tüpü kauçuk tıpa ile kapatılıp 10 dakika çalkalama makinesinde çalkalanmıĢ ve sonrasında 4000 devir/Dakika dönen santrifüj makinesinde santrifüj edilmiĢtir. Berrak eriyikten 20 ml alikot porselen kapsüle konularak üzerine 1 ml potasyum kromat indikatörü ilâve edilmiĢ ve gümüĢ nitrat (AgNO3) eriyiği ile titre edilmiĢtir. Klorun tamamı gümüĢ klorür halinde çökeldiği zaman ortamda teĢekkül eden açık kahverengi gümüĢ kromat titrasyonda son noktayı gösterecektir. Kacar (1972)’a göre numunedeki klor miktarı aĢağıdaki formüle göre hesaplanmıĢtır.
Numunede ekstrakte edilebilir klor (%): NA-B X100
Kullanılan eĢitlikte; N: Numune titrasyonunda kullanılan gümüĢ nitrat miktarı (ml), B: Blank (boĢ) titrasyonunda kullanılan gümüĢ nitrat miktarı (ml) ve A: Analiz için alınan bitki numunesi miktarı (mg). (1 ml AgNO3 1 mg Cl’a tekabül etmektedir.)
3.2.2.3.1. Sodyum (Na), Potasyum (P) ve Kalsiyum (Ca) Analizi
Onucu hafta sonunda kurutulan bitki materyalleri yakma sırasında kullanılacak beyaz tüplere tartılarak (0.2 gr) aktarılmıĢtır. Her bir örneğin üzerine 5 ml nitrik asit (HNO3) ve 2 ml hidrojen peroksit (H2O2) ilave edilmiĢtir. Yakma iĢlemi mikrodalgada gerçekleĢtirilmiĢtir. Yakma iĢlemi tamamlanınca artıklar 25 ml’lik cam tüplere
aktarılarak numunelerin hacmi saf su ile 25 ml’ye tamamlanmıĢtır. Daha sonra filtre kâğıdıyla süzülerek farklı tüplere aktarılarak numuneler bir okumada 28 elementin miktarını belirleyebilen ICP-AES cihazında elementler otomatik olarak analiz edilmiĢtir. ICP-AES ile bu çalıĢma için hazırlanan materyaller sadece K, Ca ve Na analizlerine tabi tutulmuĢtur.
3.2.3. Ġstatistikî analiz ve değerlendirmeler
AraĢtırmadan elde edilen değerler JMP 7 paket programı kullanılarak “Tesadüf Parselleri Deneme Desenine” göre varyans analizine tabi tutulmuĢtur. F testi yapılmak sureti ile farklılıkları tespit edilen iĢlemlerin ortalama değerleri LSD önem testine göre gruplandırılmıĢtır.
4. ARAġTIRMA SONUÇLARI VE TARTIġMA
Denemede ele alınan tuz seviyelerinde 6 adet arpa genotipinin farklı morfolojik ve fizyolojik özellikleri incelenmiĢ ve elde edilen özellikler ayrı ayrı baĢlıklar altında verilmiĢtir.
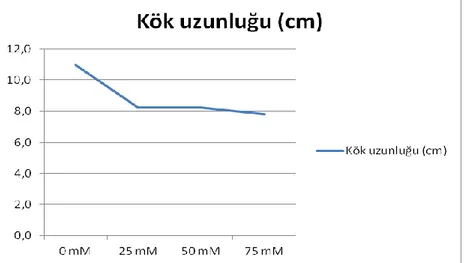
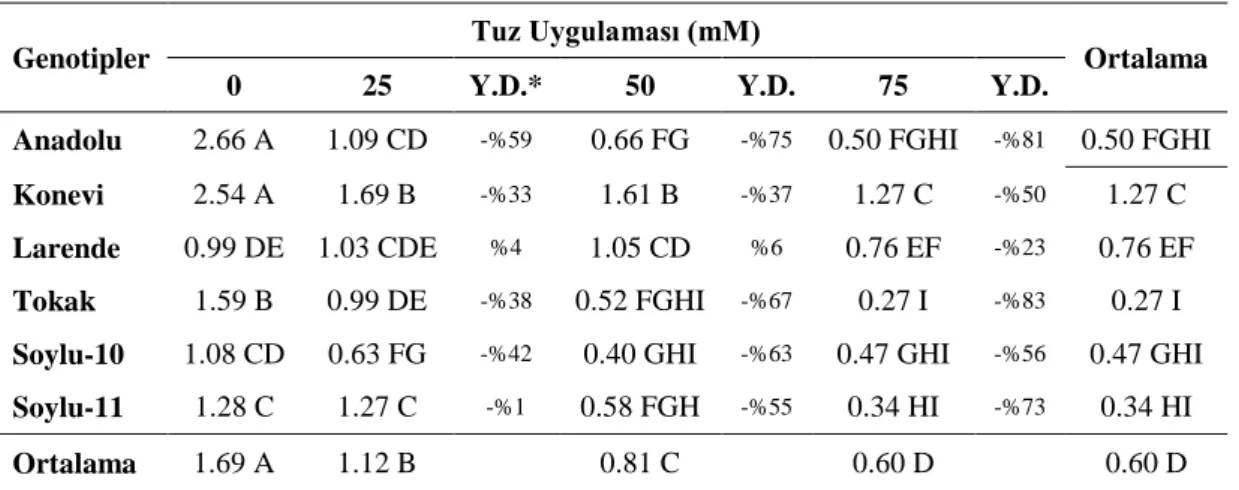
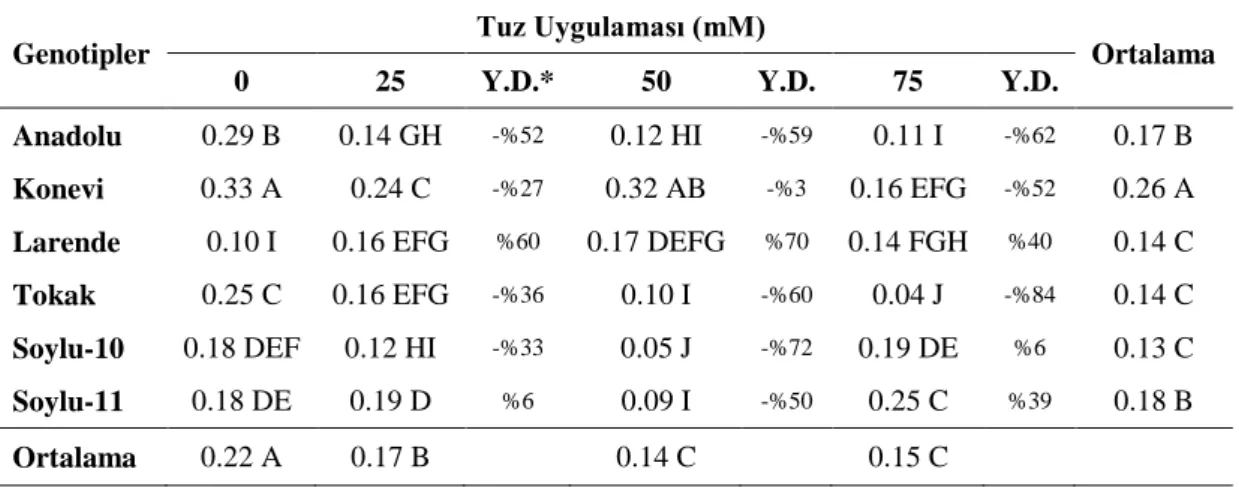
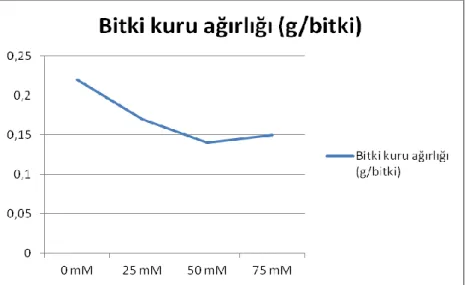
4.1. Büyüme Parametreleri 4.1.1. Bitki ve kök uzunluğu
Dört farklı tuz konsantrasyonunda 10 hafta yetiĢtirilen 6 adet arpa genotipinde ölçülen bitki uzunlukları Çizelge 4.1.’de verilmiĢtir. Yapılan çalıĢmada gövde uzunlukları arasındaki farklılık genotip, tuz uygulaması ve genotip x tuz uygulaması arası interaksiyonu bakımından %1 seviyesinde önemli bulunmuĢtur.
Genotipik açıdan en yüksek gövde uzunluğu ortalaması 38.1 cm ile “Konevi” çeĢidinden elde edilmiĢtir. Bunu azalan sıra ile 33.42 cm ile “Larende”, 33.14 cm ile “Anadolu”, 32.09 cm ile “Tokak”, 30.44 cm ile Soylu-10 çeĢitleri izlemiĢtir. En düĢük bitki uzunluğu ortalaması ise 29.68 cm ile “Soylu-11” olarak belirlenmiĢtir.
ġekil 4.1. Farklı tuz uygulamalarında yetiĢtirilen arpa genotiplerinin 10. hafta sonundaki bitki uzunluğu ortalaması
Farklı tuz seviyeleri içerisinde ġekil 4.1.’e göre en yüksek bitki uzunluğu 35.08 cm ile kontrol uygulamasında belirlenmiĢtir. Bunu azalan sıra ile 25 mM NaCl’de 33.40 cm, 50 mM NaCl’de 32.51 cm ve 75 mM NaCl’de ise 29.53 cm olarak takip etmiĢtir.
Çizelge 4.1. Arpa çeĢitlerinde tespit edilen bitki uzunlukları (cm/bitki) ve % değiĢimleri
Genotipler Tuz Uygulaması (mM) Ortalama
0 25 Y.D.* 50 Y.D. 75 Y.D.
Anadolu 39.8 AB 28.4 HIJ -%27 33.3 CDEFG -%16 31.1 EFGH -%22 33.1 B Konevi 35.6 BCDE 37.9 BC %6 42.8 A %20 36.1 BCD %2 38.1 A Larende 34.5 CDEF 35.1 CDE %2 31.2 EFGH -%10 32.9 DEFGH -%5 33.4 B Tokak 39.8 AB 33.6 CDEFG -%16 30.3 FGH -%24 24.7 IJ -%38 32.1 BC Soylu-10 33.5
CDEFG 31.5 DEFGH -%6 28.5 HIJ -%15 28.3 HIJ -%16 30.4 CD
Soylu-11 31.6
DEFGH 33.9 CDEF %7 29.1 GHI -%8 24.2 J -%23 29.7 D
Ortalama 35.1 A 33.4 B 32.5 B 29.5 C
*Y.D. Kontrol uygulamasına göre hesaplanmıĢ % değiĢim oranları.
Genotip x tuz uygulamaları interaksiyonu incelendiğinde; çeĢitler aynı tuzluluk derecelerine gövde uzunlukları değiĢik oranlarda tepki göstermiĢlerdir. ÇeĢitler içerisinde; Tokak çeĢidinde en yüksek bitki boyu kontrol uygulamasından elde edilmiĢ ve tuz konsantrasyonu arttıkça bitki boyları sırasıyla %16, %24, %38 oranında azalmıĢtır. Benzer Ģekilde kontrole göre Soylu-10’da %6, %15, %16; Anadolu çeĢidinde %27, %16, %22; Larende çeĢidinde 25 mM’da %2 oranında artıĢ olurken, 50 mM ve 75 mM tuz konsantrasyonunda sırasıyla %10, %5 azalma meydana gelmiĢtir. Soylu-11 ve Konevi çeĢidinde ise 25 mM tuz uygulaması ile birlikte bitki boyları sırasıyla %7 ve %6 oranında artıĢ gösterirken, 50 ve 75 mM tuz uygulamasında Soylu-11’ta kontrole göre azalma Ģeklinde değiĢmiĢtir. Konevi çeĢidinde ise azalma meydana gelmemiĢtir.
Bu durum tuz stresinin artırılması ile bütün çeĢitlerin bitki boyları azalırken Konevi çeĢidinin bu tepkisi bitki boyuna göre bu çeĢidin tuza dayanıklılığını göstermektedir.
Konu ile ilgili yapılan araĢtırmalarda Mer ve ark. (2000) tuzlu koĢullarda büyüyen bitkilerin büyüme hızlarının düĢük olduğunu ve bitkinin bodur bir yapı sergilediğini belirtmiĢleridir. Bunun, hücre büyümesi ve bölünmesinde meydana gelen yavaĢlamadan olduğunu ve bu yavaĢlamanın bitki bünyesindeki sitokinin miktarındaki azalma sonucu ortaya çıktığını ileri sürmüĢlerdir. Tuzlu koĢullarda yetiĢtirilen H. marinum ve H. vulgare fidelerinin kontrol gruplarına göre yaprak boy uzunluklarında azalma ve nodyum aralıklarında kısalma görülmüĢtür (Kesercioğlu, 1986). Benzer Ģekilde Islam ve ark. (2007) ve Malik ve ark. (2008), 200 mM NaCl’nin H. marinum’un gövde uzunluğunda azalma oluĢturduğunu rapor etmiĢlerdir. Irshad ve ark. (2002),