T.C.
SELÇUK ÜNĠVERSĠTESĠ SAĞLIK BĠLĠMLERĠ ENSTĠTÜSÜ
AKUT YÜZME EGZERSĠZĠ YAPTIRILAN SIÇANLARDA
RESVERATROL UYGULAMASININ KAN LEPTĠN VE
KARACĠĞER GLĠKOJEN DÜZEYLERĠNE ETKĠSĠ
Mürsel Oğuzhan Duran
YÜKSEK LĠSANS TEZĠ
FĠZYOLOJĠ TIP ANABĠLĠM DALI
DanıĢman
Prof. Dr. Abdülkerim Kasım BALTACI
T.C.
SELÇUK ÜNĠVERSĠTESĠ SAĞLIK BĠLĠMLERĠ ENSTĠTÜSÜ
AKUT YÜZME EGZERSĠZĠ YAPTIRILAN SIÇANLARDA
RESVERATROL UYGULAMASININ KAN LEPTĠN VE
KARACĠĞER GLĠKOJEN DÜZEYLERĠNE ETKĠSĠ
Mürsel Oğuzhan Duran
YÜKSEK LĠSANS TEZĠ
FĠZYOLOJĠ TIP ANABĠLĠM DALI
DanıĢman
Prof. Dr. Abdülkerim Kasım BALTACI
Bu araştırma Selçuk Üniversitesi Bilimsel Araştırma Projeleri Koordinatörlüğü tarafından 14202012 proje numarası ile desteklenmiştir
i
ÖNSÖZ
Egzersizin serbest yağ asidi ve glikoz metabolizmasını artırdığı bilinmektedir. Leptinin gıda alımını azaltmak ve enerji tüketimini artırmak gibi fizyolojik etkileri göz önüne alındığında, birçok araştırıcı leptin ve egzersiz arasındaki ilişkiyi araştırmaya yönelmiştir. Sonuçlar çelişkili olmakla beraber fiziksel aktivitenin leptin salınımında değişiklikler meydana getirdiği kabul edilmektedir. Egzersizin leptin konsantrasyonunu artırdığını ileri süren bulguların aksine, akut egzersizin leptin düzeylerini değiştirmediği, egzersizin leptin üretimini baskılamadığını bildiren raporlar da söz konusudur. Buna karşın; akut egzersizden 9 saat sonra leptin düzeylerinde bir azalmaya dikkat çekilirken, uzun mesafe yüzücülerinde veya maraton koşucularında yoğun egzersizi takiben leptin düzeylerinde anlamlı azalmaların meydana geldiğini bildiren yayınlarda bulunmaktadır.
Resveratrol (3,4,5-trihidroksistilben) üzüm tanelerinde bol miktarda bulunan polifenol yapıda doğal bir antioksidan maddedir. Yapılan klinik ve deneysel çalışmalarda resveratrolün trombosit agregasyonunu engellediği, dokuları iskeminin zararlı etkilerinden koruduğu ortaya konmuştur. Resveratrol‟ün; antioksidan, antikanser, östrojenik, antiplatelet, iskemi-reperfüzyon hasarından koruyucu, antienflamatuvar, antimikrobiyal aktiviteye sahip olduğu gösterilmesine karşın egzersiz ve performansla ilişkisini araştıran yayınlar oldukça sınırlıdır. Karaciğer glikojeninin egzersizde azaldığı, tüketici bir egzersizin karbonhidratların kullanımında defektler oluşturduğu rapor edilmiştir. Sağlıklı bireylerde orta ve yüksek yoğunluktaki egzersizin karaciğer kaynaklı glukoz üretiminde artışa yol açtığı bildirilmiştir. Egzersizden hemen sonra karaciğerde glukoz üretimi başlar, fiziksel aktivite devam ettikçe de bu üretim yoğun olarak egzersiz sonuna kadar artan bir şekilde devam eder.
Yukarıdaki bilgiler bir arada değerlendirildiğinde, resveratrol-egzersiz, resveratrol-karbonhidrat metabolizması ve leptin-egzersiz arasında olası ilişkilerden bahsedilebilir. Bu çalışmanın amacı da akut yüzme egzersizi yaptırılan sıçanlarda resveratrol uygulamasının plazma leptin ile karaciğer glikojen düzeylerini nasıl etkilediğini araştırmaktır
ii ĠÇĠNDEKĠLER: Sayfa SĠMGELER VE KISALTMALAR iv 1.GĠRĠġ 1 1.1. Leptin 1 1.1.1 Leptin Reseptörleri 1
1.1.2. Leptinin etki Mekanizması 2
1.1.3.Leptin Serbestlenmesi 2
1.1.4.Leptinin Fizyolojik Fonksiyonlar Üzerine Etkisi 3
1.1.4.1 Leptin Ve İskelet 3
1.1.4.1.1. Leptin-İskelet Etkileşimleri: (İn vitro deneysel) Kanıtlar 4 1.1.4.1.2.İnsan Gelişiminde Leptin ve Kemik Metabolizması 5 1.1.4.1.3.Yetişkin İskeletinde Leptin ve Kemik Metabolizması 7
1.1.5.Leptinin Üreme Sistemi Üzerine Etkisi 8
1.1.6.Ġnfeksiyon ve Ġnflamasyonda Leptin 9
1.1.7. Leptinin Beslenme Davranışı Üzerine Etkisi 11
1.2. Leptin ve Egzersiz 13
1.3.Resveratrol 14
1.3.1. Resveratrol Metabolizması 16
1.3.2.Resveratrolun Antioksidan Etkileri 17
1.3.3.Resveratrol ve Egzersiz 18
2. GEREÇ VE YÖNTEM 20
iii
2.1.2. Deney Hayvanları 20
2.2.Yüzme Egzersizi 21
2.3.Analizler 22
2.3.1.Plazmada Leptin Analizi 22
2.3.2.Ġmmünohistokimyasal Yöntemle Karaciğer Dokusunda
Glikojen Analizi 22
2.4.Ġstatistiksel Değerlendirmeler 23
3. BULGULAR 24
3.1.Plazma Leptin Bulguları 24
3.2.Karaciğer Glikojen Bulguları 25
4.TARTIġMA ve SONUÇ 28
4.1.Bulguların Tartışılması 28
4.1.1.Plazma Leptin Düzeylerinin Tartışılması 28 4.1.2.Karaciğer Glikojen Düzeylerinin Tartışılması 30
5. SONUÇ ve ÖNERĠLER 32 6.ÖZET 33 7.SUMMARY 34 8. KAYNAKLAR 35 9.EKLER 42 10. ÖZGEÇMĠġ 43
iv
SĠMGELER KISALTMALAR: Simgeler Açıklamalar
ALT Alanin amino tranferaz
ARN Arkuatnukleus
AST Aspartat amino transferaz
BMD Kemik mineral yoğunluğu
BMI Vücut kitle indeksi
CART Anfetamin düzenleyici transkript
cm Santimetre
CRH Kortikotropinserbestleştirici hormon
DMN Dorsomedialnukleus
FSH Folikülstimule edici hormon GnRH Gonadotropinreleasing hormon hMS2 Ġnsan kemik iliği hücreleri ICTP Kemik rezorbsiyon markerı
IL Ġnterlökin
ĠBH Ġnce bağırsak hastalıkları
kDA Kilodalton
Kg Kilogram
LHA Lateralhipotalamik alan
LH Luteinize edici hormon
v
MCH Melanin konsantre edici hormon
Mg Miligram Ml Mililitre NPY Nöropeptid Y NO Nitrik oksit NTx N-crosslinkedtelopeptid Na Sodyum Ng Nanogram
PAS Periyodik asit shift
PC Bilgisayar
PVN Paraventriküler nükleus PICP Kemik formasyonu markerı
RIA Radioimmunassay
STAT Signal ve transkripsiyon aktivatörü
1
1.GĠRĠġ
1.1. Leptin
Leptin, yağ doku tarafından sentezlenen ve serbestlenen yakın zamanda tanımlanmış protein yapılı bir hormondur (Funahashi ve ark1999, Haynes ve ark 1997, Kaplan 1998, Tritos ve ark 1997).
Adipoz dokunun bir hormon salgılayarak vücüt ağırlığını kontrol edebileceği fikri ilk olarak 1953 yılında ortaya atılmış ve lipostatik teori adını almıştır (Tritos ve ark 1997).
Yaklaşık 20 yıl sonra, normal ve genetik olarak obez fareler kullanılarak yapılan parabiozis deneylerinde lipostatik teoriyi destekleyen sonuçlar elde edilmiş ve hormonal bir faktörün iştah sinyali olarak etki edebileceği öne sürülmüştür (Considine ve ark 1997, Schwartz ve ark 1997, Tritos ve ark 1997).
1994 yılında Friedman grubundan Zhang ve arkadaşları (1994) sıçanlarda yaptıkları pozisyonel klonlama yöntemi ile şişmanlık genini ve bu genin 167 aminoasitten oluşan ürününü leptin hormonu olarak tanımlamışlardır (Haynes ve ark 1997, Kaplan 1998, Tritos ve ark 1997).
Leptin fare ve insan plazmasında 16 kDA‟luk moleküler ağırlığa sahip bir protein olarak dolaşır (Meister 2000).
Leptin, Yunanca zayıf anlamına gelen leptos kelimesinden türetilmiştir.
1.1.1 Leptin Reseptörleri
Leptin reseptörleri (Ob-R) klas I sitokin reseptör familyasının bir üyesidir (Frühbeck ve ark 1998).
I125 –leptin bağlayan bölgelerin gösterilmesinden sonra leptin reseptörleri ekspresyon klonlaması kullanılarak ilk olarak fare koroid fleksusundan izole edilmiştir (Meister 2000).
2
Ob-Ra, Ob-Rb, Ob-Rc, Ob-Re ve Ob-Rf olmak üzere altı varyantı bulunmaktadır ( Haynes ve ark 1997).
Leptin metabolik etkilerinin çoğunu merkezi sinir sisteminde ve periferik dokularda (akciğer, böbrek, karaciğer, pankreas, andrenal bezler, overler, hematopoietik hücreler) bulunan spesifik reseptörlerle etkileşerek gösterilir (Teker ve ark 2002).
Leptin reseptörleri, beyin hipotalamus bölgesindeki arkuat nükleus (ARN), VMN, paraventriküler nükleus (PVN), LHA, ventral premamiller nükleus ve dorsomedial nükleusda (DMN) bulunmaktadır (Haynes ve ark 1997).
1.1.2. Leptinin etki Mekanizması
Leptin kan-beyin bariyerinin konsantrasyon gradyentine göre geçerek hipotalamusa etki eder. Leptin reseptöre bağlandıktan sonra Janus Kinase‟ı (JAK) aktive eder. Leptin reseptörlerinin hücre dışı membran içi ve intra sitoplazmik bölümleri vardır. Leptin reseptörü hücre dışı parçasına bağlandıktan sonra, reseptörün sitoplazmik kısmına tirozin fosforilasyonu oluşur. Fosfotirozini “signal ve transkripsiyon aktivatörü ” (STAT) 3, 5 ve 6 proteinlerine bağlanır. Bu proteinler dimer halinde reseptörlerden ayrılarak nükleusa girerler ve hedef gendeki DNA‟da “STAT responsive elements” bölgesinde transkripsiyonu başlatırlar. STAT‟ın aktivasyonu leptin dozuna bağlıdır. Leptin reseptörleri ile nörotransmitter /nöromodulatör ürünleri yapan golgi aparatı nöron içinde birbirine çok yakındır. Leptinin bu ürünler vasıtası ile etkisini gösterdiği ileri sürülmektedir (Fruchbeck ve ark 1998).
1.1.3.Leptin Serbestlenmesi
Leptin, hipotalamusta bulunan özel reseptörlerinden aldığı sinyal sonrasında beyaz yağ dokusundan üretilmesine karşılık çok az miktarda da kahverengi yağ dokusundan üretilmektedir (Auwerx ve ark 1998).
3
Yağ hücresinden leptinin salgılanması β-3 adrenerjik reseptör aracılığıyla olur β-3 Adrenerjik reseptör agonistinin leptin mRNA yapımını azalttığı gösterilmiştir (Prolo ve ark 1998).
Leptin diurnal ve ultradiyan bir ritme sahip olup (Rexfor ve ark 2000) yaklaşık olarak 30 dakikada dalga şekline pulsatil bir salgılanma gösterir. Serum leptin seviyesi gece yarısı ve sabahın erken saatlerinde en yüksek, öğle saatlerine yakın zamanda öğle sonrası ortalarına kadar en düşük düzeyde seyreder (Frühbeck ve ark 1998).
Leptinin seviyesi yağ dokusu kitlesi pozitif olarak ilişkilidir. Obez insanlar daha yüksek leptin mRNA‟sına ve plazma leptin seviyelerine sahiptirler. Buna karşılık açlık süresince leptin seviyeleri hızla düşer (Meister 2000). Üretilen leptin kanla taşınır ve beyin üzerinde direkt etki gösterir (Funahashi ve ark1999). Ġnsanlarda leptinin yarılanma ömrü 24.9±4.4 dakikadır (Prolo ve ark 1998).
1.1.4.Leptinin Fizyolojik Fonksiyonlar Üzerine Etkisi 1.1.4.1 Leptin ve Ġskelet
Çeşitli kanıtlara göre, kemiklerin yeniden oluşumu ve buna bağlı olarak iskelet homeostazisi, endokrin ve/veya humoral faktörler tarafından yönetilmektedir (Riggs ve Melton 1986). Antropometrik ve metabolik faktörler arasında vücut ağırlığı kemik yoğunluğunun temel belirleyicisidir (Mazess ve ark 1987). Obezlerde, obezite oluşumu yıllarında daha fazla kemik oluşmakta ve yaşamın daha sonraki dönemlerinde kemik kaybı oranı daha yavaştır (Tremollieres ve ark 1993). Bu etkiler, kas kitlesinden çok yağ kitlesi ile ilgili görünmektedir (Reid ve ark 1992). Bu güne kadar obezitenin koruyucu etkisi oldukça şaşırtıcıdır ve adrenal androgenlerin periferal yağ dokusunda estrogenlere dönüşmesi veya mekanikal yüklenme faktörleri ile açıklanabilir. Ancak bu iki mekanik model obezite ve artan kemik mineral yoğunluğu arasındaki ilişkiyi tam olarak açıklamamaktadır. Çünkü yağ kitlesi ve kemik yoğunluğu arasında, östrojen durumu ayarlandıktan sonra da bir bağ bulunmaktadır (Reid ve ark 1992) ve bu ilişki hem kilo alınan hem de kilo artışı olmayan bölgelerde eşit derecede varlığını sürdürmektedir. (HIa ve ark 1996) Leptin, vücut yağları ile pozitif olarak bağlantılı olduğundan ve reprodüktif hormonal
4
regülasyonuna katıldığından dolayı, iskelet fizyolojindeki rolünün tanımlanması giderek daha fazla ilgi çekmektedir.
1.1.4.1.1. Leptin-Ġskelet EtkileĢimleri: (Ġn vitro deneysel) Kanıtlar
Thomas ve ark (2001) insan kemik iliği hücreleri hM52-12 hücreleri üzerinde insan rekombinan leptin rolünü incelemişlerdir. Bu hücre grubu osteoblast veya adipoz fenotipi olarak farklılaşma potansiyeline sahiptir. Stromal (bağ dokusu) hücrelere leptin uygulanması mRNA ve leptin reseptörü için protein oluşması, ayrıca adiposit fenotipe karşı seçici osteoblast farklılaşmasında artış şeklinde sonuçlanmıştır. hMS2-12 hücrelerinde yer alan erken yanıt geni cbfa1‟in (Komorive ark 1997) bulunmaması ve alkalen fosfataz ile osteokalsin varlığı nedeniyle farklılaşmanın olgunlaşma evresinde gerçekleştiği sonucu çıkarılmıştır. Özet olarak Halloway ve ark (2000)‟nın bulgularına göre leptin, kemik üzerinde kültürlerde insan mononükleer hücrelerinin osteoklastogenezisini önleyebilmektedir.
Reseland ve ark (2001)‟nın bulgularına göre, kültür içinde izole edilmiş insan osteoblastları, hem leptin hem de leptin reseptörü üretebilmektedir. Bu bulgu tek bir nokta zaman ile sınırlıdır ve leptin oluşumu osteoblast farklılaşmasındaki mineralizasyon ve/veya osteosit geçişi süresi ile kısıtlanabildiği ileri sürülmüştür.
Deneylerden elde edilen ilk kanıtlara göre, leptin, osteoblastları kendiliğinden uyarma ve osteoklast farklılaşmasını önleme kapasitesine sahip bir hormondur. Bu bulguların toplanması sonucunda leptin, anabolik ve iskelet koruyucu fonksiyonlarını doğrudan etkileme potansiyeline sahip olabilir. Obezite ve kemik mineral dansitesi arasında bir bağ olduğu için, leptinin vücut kompozisyonu ve kemik mineral dansite arasındaki dengeyi yönettiği ileri sürülebilir.
Hayvanlar üzerinde yapılan ilk deney sonuçlarından elde edilen kanıtlara göre, Leptin, kemirgen iskeletleri üzerinde potansiyel bir yönetim oluşturmaktadır. Bu verilere göre, leptin aksiyonu yaş ve tür farklarına, uygulamanın zamanlama ve yöntemine bağlıdır. Leptin periferal olarak uygulandığında anabolik ve/veya iskelet koruyucu olarak görünmektedir; ancak santral uygulamada negatif etkiler ortaya çıkmaktadır. Bu nedenle, kemik üzerindeki net sonuç, serum leptin konsantrasyonu ve kan-beyin bariyer geçirgenliğine bağlı olarak, pozitif periferal ve negatif santral etkilerin kombinasyonuna sekonder olabilir. Leptinin etkileri diğer hormonların
5
varlığına bağlı görünmektedir; iskelet üzerindeki etkilerin belirlenmesi için, leptin-hormon etkileşmelerinin de detaylı olarak belirlenmesi gereklidir.
1.1.4.1.2.Ġnsan GeliĢiminde Leptin ve Kemik Metabolizması
Kanıtlara göre, leptin insan fetal gelişme sürecinde önemli bir büyüme faktörü olabilir. Göbek kordonu kan leptin konsantrasyonu ve yeni doğan bebeklerin ağırlığı, vücut kitle indeksi (BMI) ve koldaki yağ (derialtı yağ dokusu kalınlığı) arasında pozitif bir korelasyon saptanmıştır (Hassink ve ark 1997.) Fetal leptin konsantrasyonunun temel belirleyicisi yağ kütlesi birikimidir. Ogueh ve ark (2000)‟ı fetal kan leptin düzeylerini, tip I pro-kollagenin (PICP, kemik formasyonunun bir markerı) karboksi-uçlu talopeptid ve tip I kollagenin (ICTP, kemik rezorbsiyonunun markerı) düzeylerini ölçmüştür. Sonuçlara göre, leptin konsantrasyonu ve gebelik yaşı arasında pozitif bir korelasyon (r=0.240, p=0.042) ve ICTP ile negatif bir korelasyon (r=0.420, p=0.001) vardır. Leptin konsantrasyonu ve ICTP arasındaki korelasyon sonucuna göre, leptin, net bir şekilde kemik kütlesini arttırarak, kemik rezorbsiyonunu azaltabilmektedir.
Çocuklarda leptin konsantrasyonları ve adipozite arasında pozitif bir korelasyon vardır (Hassink ve ark 1996) ve bu çocuklarda, puberte dönemi, normal ağırlıktaki çocuklara göre daha erken başlamaktadır. Buna ek olarak obeziteden bağımsız olarak, hem normal kilodaki hem de obez çocuklarda, gelişmenin ilerlemeyen Tanner evrelerinde leptin konsantrasyonlarının azaldığı görülmektedir (Hassink ve ark 1996). Buna bağlı olarak postpubertal döneme oranla prepubertal dönemdeki leptin konsantrasyonlarının daha yüksek olması, daha hızlı büyüme ve gelişme için bir düzenlenme mekanizmasını yansıtabilir. Matkovic ve ark (1997)‟ı büyümenin zirve düzeyine ulaştığı 343 sağlıklı beyaz genç kız üzerinde leptin ve kemik kütlesi arasındaki ilişkiyi değerlendirmiştir. Elde edilen sonuçlar, serum leptin ve toplam vücut BMD (bone mineral density = kemik mineral yoğunluğu)‟si arasında pozitif bir ilişki olduğunu göstermektedir. (r=0.307, p<0.0001) Leptin, kemik mineral içeriği üzerinde herhangi bir etki göstermemiştir. Klein ve ark (1998)‟nın bulgularına göre, 30 normal kilolu kontrol grubu ile karşılaştırıldığında, yüksek leptin düzeylerine sahip 18 obez çocuğun kemik yoğunluğunda fark yoktur. Ancak obez çocukların, aynı pubertal evredeki obez olmayan çocuklara göre kemik
6
yaşları, kronolojik yaşlarına göre daha büyüktür. Bu araştırmalarda kemik dönüşümünün biyokimyasal ölçümleri elde edilmediği için, kemik morfolojisi veya olgunlaşmasındaki belirtilen değişikliklere neden olan iskelet modelinin oluşturulması ve/veya yeniden yapılanması spesifik mekanizmaları açıklanmamıştır.
Erken puberte, sekonder cinsiyet karakterlerinin hızlı gelişmesi ile karakterize hızlı fiziksel olgunlaşma olarak tanımlanır. Düşük doğum ağırlığı, büyümenin sonradan yakalanması çeşitli metabolik ve hormonal anormalliklerle birlikte olmaktadır (Ibanez ve ark 1999). Bu hormon bozukluklarından ikisi, hiperinsulinemi ve hiperandrojenizimdir; her ikisinde de yaşamın ilerleyen yıllarında BMD artışı gözlenmektedir. Ibanez ve ark (2000)‟nın bulgularına göre, erken puberte görülen 52 normal kilolu kızdan oluşan bir denek grubunda (yaş sınırı 6.9-14.9 yıl) BMI derecelerine göre leptin düzeyleri daha yüksektir. Bu deneklerde lumbal vertebra BMD‟sinde artış görülmüştür. (r=0.42, p<0.05) ve kontrol grubu ile karşılaştırıldığında, kemik yaşları, kronolojik yaşlarından büyüktür. Bu sonuçlara göre, leptin düzeyleri, belli koşullarda, obeziteye bağlı olmadan yüksek olabilir ve belki de, diğer endokrin faktörler arasındaki etkileşime bağlı olabilir. Ancak bu deneklerin kontrol grubu ile karşılaştırılmasında BMI kullanılmadığı için, bu bulgulara dayanılarak kesin bir sonuç çıkarılamaz.
Leptin eksikliği görülen 9 yaşındaki bir kız üzerinde Farroqi ve ark (1999)‟ı tarafından yapılan detaylı bir araştırma, leptinin iskelet fizyolojisi gelişimi üzerindeki rolü hakkında ek bilgiler sağlamıştır. Özet olarak; hasta normal doğum ağırlığına sahiptir; ancak belirgin bir hiperfaji söz konusudur ve ilk dört ayda aşırı kilo almaya başlamıştır. Obeziteye bağlı olarak, bacaklarda valgus bozuklukları gelişmiştir. Hastaya yan tibial osteotomi uygulanmıştır. Hasta 9 yaşında vücut ağırlığı 94,4 kg‟a ulaştığında (yaşa göre >99,9 uncu persentil) günlük leptin enjeksiyonu ile tedavi başlatılmıştır. Leptin dozajı, çocuğun önceden tahmin edilen normal serum leptin konsantrasyonunun % 10‟una eşittir.
Hasta, tedavinin başlanmasından sonraki 2 hafta içinde kilo kaybetmeye başlamıştır ve 12 aylık tedavi periyodunda kilo kaybı devam etmiştir. Bu sürede hasta, % 95‟i adipozite olduğu düşünülen toplam 16,4 kg kaybetmiştir. Kilo kaybına rağmen, kemik mineral kütlesi 0.15 kg artmıştır. Kemik kütlesindeki artış çeşitli faktörlere bağlı olsa da, deri altı leptin uygulanması, azalan vücut ağırlığı, adipozite
7
ve azalan yiyecek tüketimine rağmen, kemik kütlesini korumakta ve/veya potansiyel olarak arttırmaktadır.
Leptinin, insan iskelet gelişimi üzerindeki rolünü inceleyen araştırmalara göre, hormonun etkileri, anabolik bir etki veya antiresorptif etki ya da bunların tümü ile ifade edilebilir. Buna ek olarak kemik gelişimi leptine bağlı mekanizmalarla artmaktadır. Gelişme yıllarındaki leptin rolünün belirlenmesi, deneklerin kronolojik, reprodüktif ve kemik yaşı ile spesifik arşitektürel parametreler ve iskelet yeniden şekillenmesi arasında uzun süreli araştırmalarla gerçekleştirilebilecektir.
1.1.4.1.3.YetiĢkin Ġskeletinde Leptin ve Kemik Metabolizması
Leptinin yetişkin kemikleri üzerinde pozitif bir etkiye sahip olduğu ileri sürülmektedir (Thomas ve ark 2001). Bugüne kadar yapılmış olan en geniş araştırmada, Thomas ve ark (2001)‟ı, serum leptin, insülin veya östrojen düzeylerinin, tek başına ya da birlikte, adipozite ve BMD arasındaki ilişkiyi düzenlediği hipotezini test etmişlerdir. Araştırma popülasyonu 343 erkek ve 349 dişi denekten oluşmaktadır. BMD, vücut kompozisyonu ve iskelet turn-overlarının biyokimyasal belirleyicileri değerlendirilmiş ve açlık serum hormon değerleri ilişkilendirilmiştir. Bulgularına göre serum leptin düzeyleri kadınlarda BMD ile bağlantılıdır; ancak erkeklerde böyle bir ilişki yoktur. Öncelikle erkeklerde, leptinin BMD değişkenliği, total kalça, orta lateral omurga, orta distal radiusta sıra ile % 0,3, % 1 ve % 0,1 olarak belirlenmiştir. Menapoz öncesi kadınlarda bu oranlar, % 10, % 0,3 ve % 5 ve post menapozal kadınlarda ise % 19, % 6 ve % 10‟dur. Kadın denekler arasında, leptin, kemik turn-over‟unun çeşitli belirleyicileri [kemik alkalen fosfataz, osteokalsin, PICP, N-crosslinked telopeptidler (NTx)] ile ters orantı eğilimi göstermektedirler. Bu bağlantıların en sabit olanı, idrar NTx ile kaydedilmiştir. (Menopoz öncesi kadınlar için r=-0.24, p<0.01) buna göre hormon antiresorptif bir etkiye sahiptir.
Leptinin adipozite ile yakından ilgisi olmasına rağmen, Pasco ve ark (2001)‟ı, 214 sağlıklı ve obez olmayan kadın üzerinde yaptıkları bir araştırmada, serum leptin ve BMD arasında, vücut ağırlığı ve vücut yağ kütlesinden bağımsız olarak bir ilişki olduğunu göstermiştir. Spesifik olarak, yaş, vücut ağırlığı ve vücut yağ kütlesi hesaplarında kemik yoğunluğuna bağlı değişken olarak alındığında, leptinin doğal
8
logaritması ile ilişkisi, lateral omurgada önemli bir düzeye ulaşmıştır (kısmı r2=0.030, p=0.011). Ward üçgeni ve torakanterde sınır düzeyde önem taşımaktadır (kısmı-parsiyel r2
=0.012-0.017 p=0.058-0.120). Aynı zamanda ölçülen tüm bölgelerde leptin ve kemik mineral içeriği arasında pozitif korelasyonlar kaydedilmiştir. Buna ek olarak, 51 diyaliz hastasından oluşan bir denek grubunda kadınlar için, serum leptin ve distal radius kemik dansitesi (BMD) arasında pozitif korelasyon (r=0.469, p<0.02) vardır; ancak erkeklerde yoktur (Yoneda ve ark 2001).
1.1.5.Leptinin Üreme Sistemi Üzerine Etkisi
Leptinin üreme fonksiyonları ile olan ilişkisi ilk kez 1996 yılında rapor edilmiştir. Bu sonuç ob/ob genli erkek ve dişi farelerde obezite ve infertilitenin birlikte bulunması ile ortaya çıkmıştır. Gıda alımının kısıtlanması ile vücüt ağırlığındaki azalma fertilitenin düzelmesinde etkili olmamış, buna karşılık rekombinant leptin uygulaması fertilitede düzelmeye neden olmuştur (Goumenou ve ark 1999).
Gerçekten de leptinin GnRH, LH ve FSH salınımını uyararak hem erkek, hem de dişilerde puberteyi başlatan güçlü sinyaller oluşturduğuna dikkat çekilmektedir. Ancak leptin düzeyleri yönünden cinsiyetler arasında farklılıklar söz konusudur. Erkeklerde leptin seviyesi çocukluk döneminde başlayıp pubertenin erken safhasında en yüksek seviyeye ulaşırken daha sonra azalmaktadır ve sonuçta leptin seviyeleri dişilerde erkeklere göre 3-4 kat daha yüksek bulunmaktadır. Puberteden sonra serum testosteron ve testis hacmi leptin seviyesi ile ters ilişkiliyken, östrojenler leptin salgısını uyarmaktadır.
Hipotalamus-hipofiz üreme eksenindeki leptin reseptörlerinin lokalizasyonu, leptinin üreme sisteminde önemli bir nöroendokrin rolünün olabileceğini düşündürmektedir (Darrel ve ark 2002).
Leptin gonadotropin sekresyonunu uyarırken, endojen leptinin bloklanması dişi sıçandaki östrus siklusunu pulsatil luteinizan hormon sekresyonunu bozar. Leptin ve leptin reseptör yetersiz hipogonadodotropik hipogonadizm ve primer amenore birlikte görülür (Sempere ve ark 2002).
9
1.1.6.Ġnfeksiyon ve Ġnflamasyonda Leptin
Leptinin ve leptin reseptörünün yapılarının sitokinlere olan benzerlikleri nedeniyle, leptin bir sitokin olarak da sınıflandırılabilmektedir. Leptinin yapısı interlökin (IL)-6 ve IL-11 ile benzerlik gösterirken, leptin reseptörü de IL-6 reseptörü ile homoloji göstermektedir. Leptinin lökosit sentezi üzerine stimüle edici etkisinin yanı sıra, eritropoietinin eritrositler üzerindeki uyarıcı etkisini kuvvetlendirdiği gösterilmiştir. Bu nedenle, leptin eksikliği hematopoezde de aksamalara neden olur. Bakteriyel antijenlere benzer şekilde, leptin makrofajları aktive eder, makrofajların fagositik aktivitelerini artırır ve makrofajlardan pro-inflamatuvar ve anti-inflamatuvar sitokinlerin sekresyonunu uyarır. Leptin neovaskülarizasyonu ve yara iyileşmesini de hızlandırmaktadır. Ayrıca, leptin eksikliği infeksiyona ve inflamasyona yatkınlığı artırmakta ve bu artış sitokin yapımında bozuklukla ilgili bulunmaktadır (Faggioni ve ark 2001).
Leptinin doğal ve edinsel immünitede önemli rol oynadığı bilinmektedir. Ġnfeksiyon/ inflamasyon sırasında leptin düzeyinin artması nedeniyle, konağın inflamasyona verdiği yanıtta leptinin önemli bir faktör olduğu öne sürülmüştür. Ġnfeksiyonların seyri sırasında görülen anoreksinin konağın akut faz yanıtı olduğuna inanılmaktadır. Ancak, başlangıçta yararlı olmasına rağmen, uzun süreli anoreksinin iyileşmeyi geciktirip zararlı olduğu da bilinmektedir. Bakteri hücre duvarı bileşikleri (lipopolisakkaritler ve peptidoglikanlar gibi), mikrobiyal nükleik asitler ve viral glikoproteinler akut faz reaksiyonunu ve dolayısıyla da anoreksiyi tetiklemektedir. Bakteri/virüs ürünleri de proinflamatuvar sitokinlerin (IL‟ler, tümör nekroz faktörü-alfa-TNF α, interferonlar) yapımını uyarır. Sitokinler de yağ dokusunda leptin ekspresyonunu artırır. Hem mikrobik ürünler, hem de oluşan sitokinler ve leptin gıda alımını azaltır. Bu nedenle, inflamasyon ve infeksiyon sırasında gelişen anoreksiden özellikle TNF-α, IL-1 ve IL-6‟nın sorumlu olduğu ve sitokinlerin bu etkilerinde kısmen leptinin aracılık ettiği düşünülmektedir (Maruna ve ark 2001)
Leptin-yokluğu veya leptin-reseptör-yokluğu olan hayvanlarda, ya da açlık veya malnütrisyon durumlarında, immün ve inflamatuvar yanıtlar değişmektedir. Malnütrisyonun immün yetmezliğe yol açtığı ve infeksiyonun ölümcül olmasına yol açtığı bilinmektedir. Açlık özellikle T-lenfosit yanıtlarını baskılar ve infeksiyona
10
rezistansı azaltır. T lenfositlerin proliferasyonu ve gelişmesi için gerekli olan leptin, T hücre yanıtlarını da düzenler; Th hücrelerini daha çok Th1 fenotipine doğru yönlendirir. Açlık sırasındaki nöroendokrin ve immün fonksiyon bozukluklarında düşük leptin düzeyleri aracılık etmektedir. Benzer şekilde, obez, leptin-yokluğu olan ob/ob farelerde inflamasyon sonrası ölüm riski artmaktadır; bu da leptinin inflamasyon yanıtında koruyucu rol oynadığını düşündürmektedir (Langhans 2000).
Bilindiği gibi, leptinin plazma düzeyi yağ dokusu hacmi ile korelasyon göstermekte ve kalori kısıtlaması ile leptin akut olarak azalırken, yeniden-beslenme ile artmaktadır. Leptin yokluğu olan farelerde obezite, enerji kullanımında azalma ve hiperfaji görülmektedir. Kilolu ve obez çocuklar ve erişkinlerde plazma C-reaktif düzeylerinin, IL-6, TNF-α ve leptin gibi inflamasyon göstergelerinin artması, aslında obezitenin sistemik inflamatuvar bir hastalık olduğunu düşündürmektedir. Endotoksin enjeksiyonu ile sitokinlerle ve deneysel peritonit modelinde yağ dokusunda leptin mRNA‟sının arttığı, plazma leptin konsantrasyonunu arttığı ve bu etkilerde IL-1 ile TNF-α‟nın aracılık ettiği, dolayısıyla inflamasyonun akut faz yanıtında leptinin tetikleyici olduğu bildirilmiştir. Akut sepsisteki hastalarda plazma leptin konsantrasyonunda önemli artış olduğu ve leptinin sirkadiyen ritminin ortadan kalktığı da gösterilmiştir (Brzozowski ve ark 2001, Hardwick ve ark 2001).
Etiyolojileri tam olarak bilinmeyen inflamatuvar barsak hastalıklarının (ĠBH) patogenezinde rol alan immünolojik ve inflamatuvar mekanizmalar ile inflamatuvar mediyatörler konusunda pek çok çalışma bulunmaktadır. Deneysel kolit modellerinde dolaşımda artan endotoksin ve sitokinler (IL-1, TNF-α ve INF-γ) bir taraftan immün sistem hücrelerini aktive ederken (nötrofiller, makrofajlar, mast hücreleri ve T-lenfositleri), diğer taraftan aktive olan bu hücreler siklooksijenaz ürünlerinin de katılımıyla nitrik oksit (NO) üretimini arttırmaktadır. Bu şekilde, ortama salınan endotoksin ve sitokinler gastrointestinal sistemde NO aracılı hasar oluşturmaktadır. Bütün bunlara ek olarak, inflamasyon sırasında bazı hormonların da arttığına dair bulgular vardır. Örneğin, ĠBH‟yi taklit eden çeşitli hayvan modellerinde leptin düzeylerinin yükseldiği gösterilmiştir. Sıçanlarda ince barsak inflamasyonunun erken döneminde hiperleptinemi geliştiğinin ve leptin sekresyonu engellendiğinde ise kolon inflamasyonunun azaldığının gösterildiği çalışmalar, leptinin pro-inflamatuvar olduğuna işaret etmektedir. ĠBH‟da arttığı gösterilmiş olan IL-1 ve TNF-α gibi sitokinlerin enjekte edilmesi ile leptinin plazma düzeylerinin de arttığı gösterilmiştir.
11
Ayrıca, artmış plazma leptin konsantrasyonu ile inflamasyonun derecesi ve anoreksi arasında da korelasyon bulunduğu gösterilmiştir. Buna karşın, etanol veya iskemi-reperfüzyon ile oluşturulmuş mide lezyonlarında sistemik ya da santral yolla verilen leptinin koruyucu etki gösterdiği ortaya konmuştur. Bu bulguyu destekler şekilde, leptin birçok hücre tipi için bir büyüme faktörü olarak görev almaktadır ve bu nedenle özellikle kolon kanseri patogenezindeki rolü araştırılmaktadır (Fantuzzi ve Faggioni 2000).
Sonuç olarak, akut inflamasyonda anoreksiye neden olan leptin, bazı patolojik durumlarda veya deneysel modellerde pro-inflamatuvar etki gösterirken, diğerlerinde ise anti-inflamatuvar etki sağlamaktadır. Bulguların çelişkili olması, olasılıkla farklı inflamasyon modellerinin kullanılmasından ve inflamasyonların farklı dönemlerinin araştırılmasından kaynaklanmaktadır. Laboratuvarımızda yaptığımız çalışmalarda, leptinin deneysel kolit modelinde ve yanıkta anti-inflamatuvar etki gösterdiğini ortaya koyduk. Barsaktaki anti-inflamatuvar etkisinde vagal afferent liflerin ve endojen glukokortikoidlerin aracılık ettiğini gösterdik. Literatürdeki bilgilerle elde ettiğimiz veriler bir arada değerlendirildiğinde, leptinin akut inflamasyonda arttığı ve organizmanın inflamasyonla başedebilmesinde leptin yanıtına gereksinim olduğu açıkça anlaşılmaktadır (Das 2001, Takahashi ve ark 1999).
1.1.7. Leptinin Beslenme DavranıĢı Üzerine Etkisi
Leptinin etkisini gösterebilmesi için en önemli hedef organ hipotalamustur. Leptin hipotalamusta bulunan ilgili reseptör aracılığıyla gıda alımını arttırıcı etkiye sahip olan nöropeptid Y (NPY) üzerine etki ederek gıda alımını kontrol etmektedir (Auwerx ve ark 1998).
NPY nin salınımını inhibe ederek gıda alımı üzerine uyarıcı etkisini azaltmaktadır (Baydas ve ark 2001).
Leptin uygulaması vücut yağ kitlesini ve gıda alımını azaltmaktadır. Herhangi bir nedenle Leptin yetersizliği veya Leptin reseptöründeki bir değişiklik sonucu hipotalamusta NPY‟nin sentez ve salınımı artmakta, buna bağlı olarak gıda alımı artarak obezite tablosu meydana gelmektedir (Canpolat ve ark 2001).
12
Ayrıca leptinden yoksun obez sıçanlarda hem NPY hemde NPY mRNA konsantrasyonlarının yüksek olduğu gözlenmiş ve leptin verildiğinde NPY seviyesinin azaldığı görülmüştür (Baydas ve ark 2001).
Midede özellikle fundus bölgesinde yoğun olmak üzere leptin reseptörleri ve mRNA‟sı bulunmuştur. Gastroprotektif etkisiyle midedeki gastrik lezyonları azalttığı gösterilmiştir. Ġnce barsakta ve daha çok jejunumda leptin reseptörleri bulunmaktadır. Leptin NPY yapımını inhibe ederek iştahı azaltır, dolayısıyla gıda alımı azalır (Bado ve ark 1998).
Hayvanlar üzerinde yapılan çalışmalarda, leptin eksikliği gıda alımında artışa neden olmakta ve leptin uygulaması sonrası gıda alımının azaldığı gözlenirken, açlık sonucu azalan leptin salınımı gıda alımı ile birlikte tekrar artmaktadır (Remesar ve ark 1997).
Melanın – concentrating hormon (MCH) iştah arttırıcı etkiye sahip bir ajandır. MCH mRNA seviyesi obez farelerde yüksektir ve bu seviye Leptin uygulamasından sonra normale döner (Meister 2000).
Benzer bir şekilde Shirashi ve arkadaşlarının yaptığı bir çalışmadan leptin uygulaması, iştah açıcı etkiye sahip oreksin seviyesini azaltmıştır (Shiraishi ve ark 2000).
Gıda alımını baskılayıcı etkiye sahip olan, PVN„daki kortikotropin serbestleştirici hormon (CRH) nönonlarında leptin reseptörleri gösterilmiştir. Leptin PVN‟da CRH mRNA‟sını arttırarak CRH yapımını uyarır. Aynı şekilde ARN‟da bulunan kokain ve anfetamin düzenleyici transkript (CART) içeren nöronlar leptin tarafından kontrol edilir. Bu nükleusta, açlık sırasında yada obez farelerde CART mRNA‟sı düşüktür (Meister 2000).
Besin alımı ve vücut yağ dokusunun düzenlenmesindeki biyolojik kontrollerle ilgili çok ileri gelişmeler sağlanmasına rağmen, obezitenin yaygınlığındaki artış önemli sağlık problemi olarak devam etmektedir. Leptin, Zhang et al. (1994) tarafından tanımlandıktan sonra üzerinde geniş incelemeler yapılan obezite geninin 167 aminoasitli hormonal protein ürünüdür. Başlangıçta doygunluk ve enerji dengesi ile ilgili olduğu tanımlanan leptinin adipositlerden hipotalamusa
13
feedback etki ile antiobezite faktörü olduğu ileri sürülmüştür. Bu nedenle de birçok çalışma leptinin besin alımının düzenlenmesi üzerine olan etkisine odaklanmıştır.
Ġnsülin ile leptin arasında da bir ilişki mevcuttur. Açlık esnasında dolaşımdaki leptin miktarı azalırken, karbonhidratla beslenen sıçanlarda leptin düzeyinde artış gözlenmiştir. Pankreasın β hücrelerinde leptin reseptörlerinin bulunması leptinin negatif feedback yoluyla insülin sentezine etki ettiğini göstermektedir (Remesar ve ark 1997).
1.2. Leptin ve Egzersiz
Obezite alınan ve tüketilen enerji arasındaki dengenin bozulması sonucunda artan enerji fazlasının yağ dokusunda birikmesi ile karakterize olan bir hastalıktır. Obezite başta kardiyovasküler ve endokrin sistemler olmak üzere vücudun tüm organ ve sistemlerini etkileyerek bozukluklara ve hatta ölüme yol açabilen bir hastalıktır. Leptin ise enerji alım - tüketim dengesinde rol alan bir antiobezite hormonudur. Akut egzersiz ile enerji tüketiminin artırılmasının leptin düzeyleri üzerinde bir etkisi olmamasına rağmen; kronik egzersiz sonucu yağ dokusundaki azalmaya bağlı olarak leptin seviyeleri azalmakta, vücut ağırlığı ve yağ dokusu için leptin seviyesinin azalması obezite ile gelişen leptin rezistansının normalleşmeye başlaması ve leptin duyarlılığının artması yönünde değerlendirilmektedir (Unal ve ark 2004).
Egzersizin serbest yağ asidi ve glikoz metabolizmasını artırdığı bilinmektedir. Leptinin gıda alımını azaltmak ve enerji tüketimini artırmak gibi fizyolojik etkileri olduğu gözönüne alındığında, birçok araştırıcı leptin ve egzersiz arasındaki ilişkiyi araştırmaya yönelmiştir. Leptinin öncelikle morbit obez ve obezlerde kilo kontrolünde bir umut olabilir mi tartışılırken profesyonel sporcularda da vücut yağ ağırlığının kontrolünde etkinliği gündeme gelmiştir.
Leptin max VO2 „nin %50-60‟larında yapılan egzersizlerde aerobik yolla serbest yağ asitlerini yakarak vücutta yağlanmayı azaltmaktadır. Profesyonel 10 futbolcu ve sedanter üzerinde yapılan bir çalışmada egzersiz yapan grupta leptin düzeyleri sedanter gruba göre daha düşük bulunmuştur. Genel olarak serum leptin düzeylerindeki artış bireylerin BMI‟ i ile doğru orantılı olmasına rağmen düzenli
14
olarak yapılan egzersizin leptin düzeylerini baskıladığı gözlemlenmiştir (Unal ve ark 2005a).
Max VO2„nin %70„lerinde ve altında yapılan 60 dakikalık aerobik egzersizlerden sonra serum leptin seviyesinde azalma başlamakta ve egzersiz sonrası 24. Saatte bu baskılanma iyice belirgin hale gelmektedir (Kamohara ve ark1998, Koistinen ve ark 1998).
Enerji dengesini değiştirebilecek ölçüde enerji tüketimine yol açan egzersizlerin leptin düzeylerini etkileyebileceği ve uzun süreli egzersizlerden 48 saat sonra serum leptin düzeylerinin azaldığı görülmüştür (Gokbel ve ark 2005).
Antrenman başına 1200-1300 kcal enerji tüketimi olan elit sporcuların 24 saat sonraki serum leptin seviyesinin daha düşük olduğu tespit edilmiştir (Ünal ve ark 2005).
Ġn vitro çinko uygulamasının leptin üretimini artırdığı, çinko eksikliğinin ise serum leptin düzeylerini azalttığı bildirilmiştir. Çinko eksikliği ve takviyesinin serum leptin düzeyleri ile zorlu egzersiz yaptırılan ratlarda araştırıldığı bir çalışmada zorlu egzersizden 48 saat sonra leptin düzeylerinin azaldığı, çinko takviyesinin ise leptinin egzersize cevabını bozduğu görülmüştür (Gökbel ve ark 2005).
Leptin ile ilgili yapılan çalışmalar sonucu leptinin vücudun yağ durumu ve enerji depoları hakkında beyne sinyal gönderen bir molekül olduğu gösterilmiş olsa da, vücuttaki bir çok organda reseptörünün gösterilmiş olması leptinin görevinin gıda alımının kısıtlanması ve enerji kullanımının artırılması ile sınırlı olmadığını düşündürmektedir. Leptin hormonunun fizyolojik mekanizmaları üzerindeki etkisi altında hala bir takım soru işaretleri bulunmaktadır. Birçok araştırmacının ilgi alanına girmesi ve çeşitli disiplinler tarafından üzerinde çalışılıyor olması yakın bir gelecekte leptinin etki mekanizmalarının tam aydınlığa kavuşabileceği sinyalini vermektedir.
1.3.Resveratrol
Bitkisel kaynaklı çeşitli yiyecek ve içecekler, bitkiler tarafından hasara ya da fungal ataklara karsı sentezlenen birtakım non-flavonoid yapıda fenolik bileşikler içermektedirler. Bunlar arasında resveratrol; stilben fitoaleksinlerin ana aktif bileşiği olarak belirlenmiştir. Resveratrol (3,4‟,5-trihidroksistilben, C14H12O3, M.A: 228,25
15
gram) kimyasal olarak flavonoid yapıda, polifenolik, steroid olmayan bir bileşik olup östrojen benzeri biyolojik aktiviteye sahiptir. Arichi ve ark. (1982) Polyganum
Cuspidatum adlı bitkinin kurutulmuş köklerinin geleneksel Japon ve Çin tıbbında
“Kojo-kon” adı verilen drog olarak kullanıldığını belirtmiştir. Kojo-kon “Itadori” olarak da adlandırılmış ve mantar hastalıkları, inflamasyon, kalp, karaciğer ve damar hastalıklarının tedavisinde kullanılmıştır ve bütün bu yararlı etkilerden Polyganum
Cuspidatum‟un primer aktif bileşeni olan resveratrolün sorumlu olduğu gösterilmiştir
(Arichi ve ark. 1982). Resveratrol, Polyganum Cuspidatum‟dan baska üzüm, yer fıstığı ve ananasda da bulunmaktadır. Trans-resveratrol ilk olarak 1976 yılında Langcake ve Pryce tarafındanasma yapraklarında (Vitis vinifera) tespit edilmiştir ve bileşiğin fungal infeksiyona (Botrytis cinerea) veya ultraviyole (UV) ışığa maruziyete cevap olarak yaprak dokuları tarafından sentezlendiği gösterilmiştir (Langcake ve Pryce 1976). Bitkiler ayrıca resveratrolün glukozid formunu da sentezlemektedir. Resveratrolun bu glukozid formu Resveratrol-3-0-β-D-glukozid, piceid olarakadlandırılmaktadır. Trihidroksistilben yapısında olan Resveratrol 3,4‟,5‟un, trans ve cis izomeri olmak üzere iki tip izomeri vardır. Trans izomerlerinin biyolojik aktiviteleri, cis izomerlerinden daha yüksektir (Bradamante ve ark 1997). Resveratrolün kimyasal yapısı sentetik estrojen, dietilstilbestrol ile nenzerlik gösterir. Ayrıca, glukozit ve polimerleri içeren bir moleküler ailesininde ana komponenti olup asma, yer fıstığı ve dutun içinde bulunduğu gösterilmiştir (Doğan 2005). Ġlk olarak Polygonum Cuspidatum bitkisinin köklerinde tespit edilen Resveratrol, bitkiler tarafından patojenik durumlardan korunmak amaçlı yaralanma ve fungal saldırı sonucu oluşmaktadır. Ayrıca, UV ışıkla da sentezi uyarılmaktadır. Daha sonrada,
Vitis-Vinifera ve saraplarda tespit edilmiştir (Soleas ve ark 1997; Lamuela-Raventos
ve ark. 1995; Wenzel ve ark. 1998). Vitis vinifera’da (asma yaprakları) Vst 1 geni tarafından sentezlendiği gösterilen resveratrolun bu gen bombardımanıyla buğday, arpa gibi yabancı bitkilerde de bir fitoaleksin olan resveratrol sentezlenebildiği rapor edilmiştir (Wenzel ve ark. 1998). Resveratrolün doğal kaynaklardan elde edilmesi hem zaman açısından kayba, hem de maddenin daha düşük miktarlarda elde edilmesine neden olmaktadır. Araştırmalar molekülün kimyasal yolla sentezi üzerine yoğunlaşmış ve transresveratrol elde edilmiştir. Trans-resveratrolün UV irradyasyonu ile cis- formu elde edilmektedir. Yapılan çalısmalar, trans-resveratrolün ışıktan korunduğu takdirde stabilitesini ve aktivitesini koruduğunu
16
göstermektedir (Langcake ve Pryce 1976). Trihidroksistilben yapısında olan Resveratrol 3,4‟,5‟un, trans ve cis izomeri olmak üzere iki tip izomeri vardır. Trans izomerlerinin biyolojik aktiviteleri, cis izomerlerinden daha yüksektir (Bradamante ve ark 1997). Resveratrolün kimyasal yapısı sentetik estrojen, dietilstilbestrol ile nenzerlik gösterir. Ayrıca, glukozit ve polimerleri içeren bir moleküler ailesininde ana komponenti olup asma, yer fıstığı ve dutun içinde bulunduğu gösterilmiştir (Doğan 2005). Ġlk olarak Polygonum Cuspidatum bitkisinin köklerinde tespit edilen Resveratrol, bitkiler tarafından patojenik durumlardan korunmak amaçlı yaralanma ve fungal saldırı sonucu oluşmaktadır. Ayrıca, UV ışıkla da sentezi uyarılmaktadır. Daha sonrada, Vitis-Vinifera ve saraplarda tespit edilmiştir (Soleas ve ark 1997; Lamuela-Raventos ve ark. 1995; Wenzel ve ark. 1998). Vitis-Vinifera’da (asma yaprakları) Vst 1 geni tarafından sentezlendiği gösterilen resveratrolun bu gen bombardımanıyla buğday, arpa gibi yabancı bitkilerde de bir fitoaleksin olan resveratrol sentezlenebildiği rapor edilmiştir (Wenzel ve ark. 1998).
1.3.1. Resveratrol Metabolizması
Sıçanlarda yapılan çalışmalardan elde edilen sonuçlara göre oral alınan resveratrolün plazmadan eliminasyon yarı ömrünün 12-15 dakika olduğu gösterilmiştir. Bu çalısmalarda resveratrol 2 mg/kg (Juan ve ark. 2002b) ve 50 mg/kg dozlarında sıçanlara (Marier ve ark. 2002), 20 mg/kg dozunda farelere (Asensi ve ark. 2002) intragastik olarak verilmiş ve plazmada ulaştıkları doruk konsantrasyon değerleri sırasıyla 0.5, 1.2, 6.6 μM olarak bulunmuştur. Sıçan ve farelerde yapılan çalışmalarda resveratrolün iyi bir sekilde absorbe edildiği ve karaciğer ve barsak epitel hücrelerinde hızla glukuronid ve sülfat konjugatlarına dönüştürüldüğü gösterilmiştir (Andlauer ve ark. 2000; Soleas ve ark. 2001; Vitrac ve ark. 2003; Yu ve ark. 2002). Sıçanlarda işaretli resveratrolün oral olarak uygulanmasıyla dozun %50-75‟i absorbe olmaktadır. Resveratrol glukuronidin plazmada ulastıgı doruk konsantrasyon düzeyi 105 μM olarak bulunmuş ve resveratrolün yoğun bir biçimde enterohepatik sirkülasyona maruz kaldığı gösterilmiştir (Soleas ve ark. 2001). Perfüze sıçan ince barsaklarında resveratrolün % 16,8‟i resveratrol glukuronide, % 3,4‟ü resveratrol sulfata dönüşmüş ve % 0,3‟ü serbest resveratrol olarak bulunmuştur (Andlauer ve ark. 2000). Sıçan izole ince barsaklarında 200 μM resveratrol
17
perfüzyonunundan sonra jejenum ve ileumda resveratrolün büyük oranda glukuronid konjugatına dönüştüğü ve sistemik dolaşıma katıldığı gösterilmiştir (Kuhnle ve ark. 2000). Resveratrolün insan, rodent karaciğer ve barsak hücrelerinde glukuronid ve sülfat konjugatlarına dönüştüğü gösterilmiştir (Yu ve ark. 2002). Bütün bu çalışmalar dikkate alındığında resveratrol sıçan ve farelerde gastrointestinal yoldan yüksek miktarlarda absorbe edilmekte, karaciğer ve barsakta konjugasyon ile metabolize edilmektedir. Sıçanlarda 50 mg/kg gibi yüksek oral dozun uygulanmasından sonra metabolize olmamış resveratrolün doruk konsantrasyonu 10 μM‟ın altına düşmekte ve eliminasyonu oldukça hızlı olmaktadır. Aksine resveratrolün konjugatları esas moleküle göre daha yüksek plazma düzeyine ulaşmaktadırlar (Marier ve ark. 2002). Meng ve ark.‟nın (2004) yaptıkları bir çalışma 30-50 yaş arasındaki erkek gönüllüler üzerinde gerçekleştirilmiştir. Bir gruba 0.03 mg/kg, 0,5 mg/kg veya 1 mg/kg viskide çözünmüş resveratrol içirilmiştir. Diğer bir gruba da 0.16 mg/100 ml resveratrol içeren üzüm suyundan 200, 400, 6000, 1200 ml verilmiştir ve idrar örnekleri toplanmıştır. LC-MS/MS analizi ile idrarda resveratrolün bulunduğu gösterilmiştir. 0.03 mg/kg dozunda ilk 2-3 saat içinde resveratrolün büyük kısmının itrah edildiği 1 mg/kg dozunda resveratrol uygulamasında ise resveratrolün itrahının 7-10 saat arasında gerçeklestiği gösterilmiştir (Meng ve ark. 2004). Resveratrolün suda çözünürlüğü oldukça düşüktür bu yüzden serumda yüksek konsantrasyonlarda bulunabilmesi için ya konjuge olmalı ya da proteinlere bağlanmalıdır (Belguendouz ve ark. 1997). Albumin plazmada resveratrolü taşıyan moleküllerden biridir (Jannin ve ark. 2004). Plazmada resveratrolün lipoproteinlerle de etkileştiği gösterilmiştir (Belguendouz ve ark. 1998). Resveratrolün toksik etkilerini arastırmak için, Sprague-Dawley sıçanlara 20 mg/kg dozda resveratrol oral olarak 28 gün boyunca verilmiş ve hematolojik, biyokimyasal ölçümler ve organlarda histopatolojik ölçümler yapılmıştır. Hepatik toksistenin göstergesi olan ALT ve AST enzim düzeyleri ve ölçülen diger hematolojik, biyokimyasal ve histopatolojik parametreler resveratrol alımından etkilenmemiştir (Juan ve ark. 2002a).
1.3.2.Resveratrolun Antioksidan Etkileri
Resveratrol‟ün antioksidan özelliği ve peroksi radikal süpürücü etkisi ile kalbi koruduğu düşünülmektedir (Dolinsky ve ark. 2013). Ġskemi ve
iskemi-18
reperfüzyon, dokuda lipit peroksidasyonunu arttırmakta, sonuçta da serbest radikaller oluşmaktadır. Koroner oklüzyonla iskemi ile iskemi-reperfüzyon hasarının oluşturulduğu bir çalışmada resveratrol uygulamasının, koroner damarlarda lipit peroksidasyonunun göstergesi kabul edilen malonildialdehit düzeyini anlamlı olarak azalttığı bildirilmiştir (Hung ve ark. 2002). Düşük yoğunluklu lipoprotein peroksidasyonlarının demir ve bakır iyonları ile arttırıldığı bilinmektedir. Serbest radikal süpürücü etkisinin araştırıldığı bir çalışmada resveratrolün demir ve bakır düzeyini anlamlı olarak düşürdüğü gösterilmiştir (Fauconneau ve ark. 1997). Kronik etanol kullanımının beyin dahil bir çok organda reaktif oksijen türleri ve lipit peroksidasyonu ile oksidatif hasara yol açtığı bilinmektedir. Bir çalışmada dopaminerjik nöronal hücreler olan pc12 hücrelerinde etanolun lipit peroksidasyonunu arttırarak hücre ölümüne yol açtığı, ancak resveratrol uygulaması ile etanolün olumsuz etkilerinin engellendiği gösterilmiştir (Sun ve ark. 1997) . Bir diğer çalışmada sıçanlarda, etanol uygulamasının nöronlarda, lipit peroksidasyonuna oldukça duyarlı olan Na-K ATPaz ve dopamin taşıyı protein erkinin azaldığı bulunmuştur. Resveratrol uygulaması ile bu nörodejeneratif değişikliklerin oluşmadığı gözlenmiştir (Sun ve ark. 2002). Bu çalışmalara dayanılarak; Resveratrol‟ün lipit peroksidasyonunu önlediği ve siklooksijenazı baskıladığı sonucuna varılmıştır.
1.3.3.Resveratrol ve Egzersiz
Beslenme davranışının yanı sıra fiziksel egzersizin de sağlıklı yaşam tarzının önemli bir bileşeni olduğu kabul edilir. Orta şiddette bir egzersizin kalp damar hastalıkları (Daviglus ve ark. 2006), tip-II diyabet (Bertram ve ark. 2010), metabolik sendrom (Church 2011), Alzheimer hastalığı gibi nörodejeneratif hastalıklar başta olmak üzere primer ve sekonder hastalıkların önlenmesinde yararlı olabileceği gösterilmiştir (Deweerdt 2011; Hurley ve ark. 2011). Hormesis terimi, toksik maddelerin düşük konsantrasyonlarında yaşamı devam ettirebilmeyi ve bu maddelere karşı geliştirilen adaptif olguları açıklamak için öne sürülen bir kavramdır (Malaguti ve ark. 2013). Fiziksel egzersize karşı metabolizmanın gösterdiği adaptasyon mekanizması da bu kavram içerisinde değerlendirilebilir. Yoğun fiziksel aktivitenin meydana getirdiği serbest radikal üretimindeki artışa cevap olarak metabolizma
19
antioksidan sistemin aktivasyonuyla buna cevap verir (Jii ve ark. 2009). Sağlıklı yaşam için önerilmesine rağmen orta düzeyde fiziksel egzersizin, oksidatif stres, inflamasyon ve kas hasarına neden olduğu bilinmektedir (Malaguti ve ark. 2013). Bu nedenle fiziksel aktivitenin yol açtığı oksidatif strese karşı diyet stratejileri geliştirmek konusunda yoğun bir çaba söz konusudur (Malaguti ve ark. 2013). Son yıllarda resveatrolun, fiziksel performansın modülasyonu ve oksidatif hasarı önleme yeteneği araştırımaya başlanmıştır (Malaguti ve ark. 2013). Yaşlı farelerde resveratrol uygulamasının lipid peroksidasyonunu önlediği gösterilmiştir (Murase ve ark. 2009). Benzer bulgular Dal-Ros ve ark. (2009) tarafından da gösterilmiştir. On dört sporcu üzerinde gerçekleştirilen bir çalışmada resveratrol uygulamasının egzersizin yol açtığı lipid peroksidasyonunu önlediği bildirilmiştir (McAnulty ve ark. 2013).
20
2. GEREÇ VE YÖNTEM
2.1. Hayvan Materyali ve Gruplar
Bu çalışma Necmettin Erbakan Üniversitesi Deneysel Tıp Uygulama ve Araştırma Merkezinden temin edilen sıçanlar üzerinde aynı merkezde gerçekleştirildi. Wistar-Albino cinsi 28 adet erişkin erkek sıçan kullanılan araştırmada, gruplar şu şekilde oluşturuldu.
Grup 1; (n:7) Kontrol grubu: Hiçbir uygulamanın yapılmadığı standart diyetle beslenen grup.
Grup 2; (n:7) Yüzme Grubu: Standart diyetle beslenen ve çalışmanın bitiminde 30 dakika akut yüzme egzersizi yaptırılan grup.
Grup 3; (n:7) Resveratrol Grubu: Günlük standart diyete ek olarak dört hafta boyunca içme suyu içerisinde (10 mg/kg) resveratrol verilen grup.
Grup 4; (n:7) Resveratrol Uygulanan Yüzme Grubu: Günlük standart diyete ek olarak dört hafta boyunca içme suyu içerisinde (10 mg/kg) resveratrol verilen ve çalışmanın bitiminde 30 dakika akut yüzme egzersizi yaptırılan grup.
2.1.2. Deney Hayvanları:
Deney hayvanları yıkamak suretiyle her gün temizlenen özel çelik kafeslerde beslendi. Yemler özel çelik kaplarda, su cam biberonlarda (normal çeşme suyu) verildi. Hayvanlar her gün vücut ağırlıklarının 100 gramı başına yaklaşık 10 g yemle beslendiler. Hayvan yemleri, normal sıçan yemi (pelletler halinde) olarak Necmettin Erbakan Üniversitesi Deneysel Tıp Araştırma ve Uygulama Merkezinden temin edildi. Deney hayvanları 12 saat karanlık, 12 saat aydınlık ve standart oda sıcaklığı (21±1oC) sağlanan ortamda tutuldu. Resveratrol uygulamaları sıçan başına 10 mg/kg/gün olacak şekilde içme suyu içinde grup 3 ve 4‟ü oluşturan sıçanlar üzerinde gerçekleştirildi. Dört hafta süren çalışmaların bitiminde sabah 09:00
-10:00 saatleri arasında dekapite edilen hayvanlardan kan ve karaciğer doku örnekleri alındı.
21
2.2.Yüzme Egzersizi
Egzersiz uzunluğu 80 cm, yüksekliği ve genişliği ise 50 cm olan ısıya dayanıklı (polietilen), yüzme havuzunda gerçekleştirildi. Egzersizler resveratrol uygulamalarının bitiminden 24 saat sonra, bir defaya mahsus olmak üzere 30 dakika süre ile yapıldı. Ġkişerli gruplar halinde yüzdürülen deney hayvanları yüzme egzersizinden hemen sonra dekapite edilerek kan (plazma) ve karaciğer doku örnekleri alındı.
Çizelge 2.2.1.Deney hayvanlarına verilen yemin bileşimi
Yem maddeleri Yüzdesi (%) Buğday Mısır Arpa Kepek Soya Küspesi Balık Unu E-Kemik unu Melas Tuz *Vitamin Karması **Mineral Karması 10 21 14 8 25 8 4 4 4 1 1
*Vitamin karması: Deney hayvanlarına verilen yemlerin vitamin karmasında A, D3, E, K, B1, B2, B6, B12 vitaminleri ile nikotinamid, folik asit, D-biotin ve kolin klorit bulunmaktadır.
**Mineral karması: Mangan, demir, çinko, bakır, iyot, kobalt, selenyum ve kalsiyumdan oluşmuştur.
22
2.3.Analizler
2.3.1.Plazmada Leptin Analizi
Plazmada leptin analizi, Rat Leptin RIA test kiti (Linco marka katolog no: RL-83K) kullanılarak yapıldı. Analizler şu şekilde yapıldı:
1.gün: Total count tüpü haricindeki diğer tüplere (NSB, Bo, Standart, Kontrol ve Numune tüplerine) analiz tampon çözeltisinden ilave edildi. Standart olarak 0.5 ng/ml, 1ng/ml, 2 ng/ml, 5 ng/ml, 10 ng/ml, 20 ng/ml, 50 ng/ml kullanıldı. Total, NSB, Bo, Standart ve Kontrol tüpleri iki kez çalışıldı. Total ve NSB tüpü hariç diğer tüplere Rat Leptin Antibody ilave edildi. Vortekslendi. Tüplerin ağzı kapatılarak oda sıcaklığında 20 saat inkübe edildi.
2.gün: Bütün tüplere I-125 Rat Tracer ilave edildi. Vortekslendi. Tüplerin ağzı kapatılarak oda sıcaklığında 22 saat inkübe edildi.
3.gün: Total tüp hariç diğer tüplere çöktürücü çözelti ilave edildi. +4 0
C de 20 dakika inkübe edildi. +4 0C de 4500 devir/dakika‟ da 20 dakika santrifüj edildi. Süpernatanları 60 saniye içinde alınarak atıldı. Tüpte kalan çökeltiler Gamma Counter yardımıyla ölçüldü. Değerler ng/ml olarak hesaplandı.
Rat Leptin analizinin limit sensivitesi 0.5 ng/ml ve limit linearitesi 50 ng/ml dir.
2.3.2.Ġmmünohistokimyasal Yöntemle Karaciğer Dokusunda Glikojen Tayini
Karaciğer doku örnekleri % 95 lik etil alkol içerisinde tespit edildi. Tespit olan dokulardan alınan örnekler ototeknikon takibine tutuldu. Ototeknikon takibi sonrası parafine gömülen doku örneklerinden mikrotom yardımı ile 5µm lik kesitler, lama alınarak PAS ile boyandı. Boyanan preparatlar Nikon Eclipse E400 ışık mikroskopu ile değerlendirildi. Işık mikroskopuna bağlı Nikon Coolpix 5000 digital fotoğraf makinesi ile uygun alanların görüntüleri fotoğraflandı. Fotoğraflama esnasında kalibrasyon amacı ile Nikon Stage Micrometer görüntüsü de aynı mikroskobik büyütmelerde alındı. Tüm görüntüler PC ortamına aktarıldı ve Clemex PE 3.5 görüntü analizi programı ile değerlendirildi. Görüntü analizi programı ile 0.1
23
mm² lik alanlar seçildi. Bu alanlardaki glikojen içeren (PAS ile pozitif boyanan) hepatositler işaretlenerek otomatik olarak saydırıldı.
2.4.Ġstatistiksel Değerlendirmeler
Bulguların istatistiksel değerlendirilmesi SPSS 21.0 bilgisayar paket programı ile yapılarak, bütün parametrelerin aritmetik ortalamaları ve standart sapmaları hesaplandı.Verilere uygulanan Shapiro-Wilks testine göre verilerin normal dağılım gösterdiği belirlendi. Gruplararası farklılıkların tespitinde Tek Yönlü Varyans Analizi, farklılığın hangi gruptan kaynaklandığının belirlenmesi için ise Asgari Önem Fark (Least Significant Difference “LSD”) testi kullanıldı. P<0,05 düzeyindeki farklılıklar anlamlı olarak kabul edildi.
24
3. BULGULAR
3.1.Plazma Leptin Bulguları
Çalışmamızda en yüksek plazma leptin düzeyleri Grup 1 (Kontrol) ve Grup 3 (Resveratrol uygulanan grup)‟de elde edildi (P<0.05). Grup 2 (Yüzme) ve Grup 4 (Resveratrol uygulanan yüzme)‟ün plazma leptin düzeyleri Grup 1 ve Grup 3‟den düşük (P<0.05) iken, her iki grubun (Grup 2 ve Grup 4) plazma leptin değerleri ise birbirinden farklı değildi (Çizelge3.1.1.).
Çizelge3.1.1. Çalışma Gruplarının Plazma Leptin Düzeyleri
Gruplar (N=7) Leptin Düzeyleri (ng/ml)
Kontrol Grubu (Grup 1)
115,81 ± 16,46A
Yüzme Grubu (Grup 2)
84,58 ± 12,73B
Resveratrol Grubu (Grup 3)
118,70 ± 29,04A
Yüzme+Resveratrol Grubu (Grup 4)
88,80 ± 10,75B
*A,B: Aynı sütunda farklı harf taşıyan ortalamalar arasındaki fark istatistiksel açıdan önemlidir (P<0,05).
25
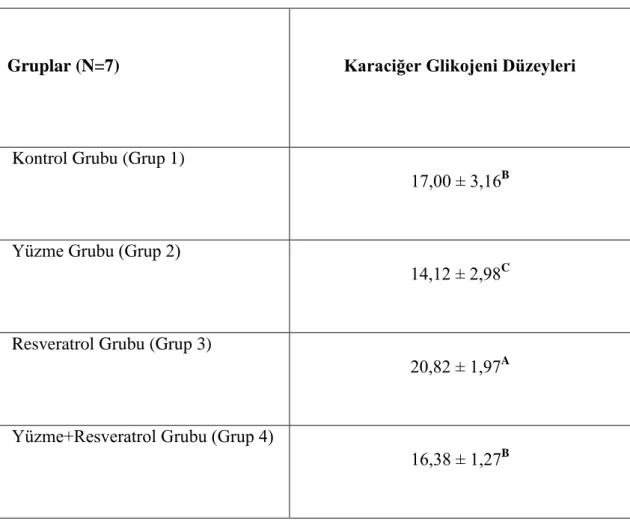
3.2.Karaciğer Glikojen Bulguları
Çalışmamızda en yüksek karaciğer glikojen seviyeleri Resveratrol uygulanan Grup 3‟de (P<0.05), en düşük karaciğer glikojen düzeyleriyse grup 2 (yüzme grubu)‟de elde edildi. Kontrol (Grup 1) ve Resveratrol Uygulanan Yüzme (Grup 4) gruplarının karaciğer glikojen düzeyleri arasında önemli bir farklılık tespit edilmedi (Çizelge 3.2.1., Resim 3.2.1., Resim 3.2.2. , Resim 3.2.3. , Resim 3.2.4.).
Çizelge 3.2.1. ÇalıĢma Gruplarını Karaciğer Glikojen Düzeyler
Gruplar (N=7) Karaciğer Glikojeni Düzeyleri
Kontrol Grubu (Grup 1)
17,00 ± 3,16B
Yüzme Grubu (Grup 2)
14,12 ± 2,98C
Resveratrol Grubu (Grup 3)
20,82 ± 1,97A
Yüzme+Resveratrol Grubu (Grup 4)
16,38 ± 1,27B
*A,B: Aynı sütunda farklı harf taşıyan ortalamalar arasındaki fark istatistiksel açıdan önemlidir (P<0,05).
26
Resim 3.2.1.Grup 1’e ait Karaciğer Glikojeni (normal karaciğer glikojeni)
27
Resim 3.2.3. Grup 3 resveratrol uygulamasıyla artmıĢ karaciğer glikojeni
Resim 3.2.4. Grup 4 Egzersizle resveratrol uygulamasıyla korunmuĢ karaciğer glikojeni
28
4.TARTIġMA ve SONUÇ
4.1.Bulguların TartıĢılması
4.1.1.Plazma Leptin Düzeylerinin TartıĢılması
Egzersizin serbest yağ asidi ve glikoz metabolizmasını artırdığı bilinmektedir. Leptinin gıda alımını azaltmak ve enerji tüketimini artırmak gibi fizyolojik etkileri olduğu gözönüne alındığında, birçok araştırıcı leptin ve egzersiz arasındaki ilişkiyi araştırmaya yönelmiştir. Leptinin öncelikle morbit obez ve obezlerde kilo kontrolünde bir umut olabilir mi tartışılırken profesyonel sporcularda da vücut yağ ağırlığının kontrolünde etkinliği gündeme gelmiştir.
Leptin max VO2 „nin %50-60‟ larında yapılan egzersizlerde aerobik yolla serbest yağ asitlerini yakarak vücutta yağlanmayı azaltmaktadır. Profesyonel 10 futbolcu ve sedanter üzerinde yapılan bir çalışmada egzersiz yapan grupta leptin düzeyleri sedanter gruba göre daha düşük bulunmuştur. Genel olarak serum leptin düzeylerindeki artış bireylerin BMI‟ i ile doğru orantılı olmasına rağmen düzenli olarak yapılan egzersizin leptin düzeylerini baskıladığı gözlemlenmiştir (Ünal ve ark 2005b).
Max VO2„nin %70„lerinde ve altında yapılan 60 dakikalık aerobik egzersizlerden sonra serum leptin seviyesinde azalma başlamakta ve egzersiz sonrası 24. Saatte bu baskılanma iyice belirgin hale gelmektedir (Kamohara ve ark1997, Koistinen ve ark 1998).
Enerji dengesini değiştirebilecek ölçüde enerji tüketimine yol açan egzersizlerin leptin düzeylerini etkileyebileceği ve uzun süreli egzersizlerden 48 saat sonra serum leptin düzeylerinin azaldığı görülmüştür (Gokbel ve ark 2005).
Antrenman başına 1200-1300 kcal enerji tüketimi olan elit sporcuların 24 saat sonraki serum leptin seviyesinin daha düşük olduğu tespit edilmiştir (Ünal ve ark 2004).
Sonuçlar çelişkili olmakla beraber fiziksel aktivitenin leptin salınımında değişiklikler meydana getirdiği kabul edilmektedir. Egzersizin leptin konsantrasyonunu artırdığını ileri süren bulguların (Van Agel-Leijssen ve ark 1999) aksine, Hickey ve Calsbeek (2001) akut egzersizin leptin düzeylerini
29
değiştirmediğini bildirirken, benzer şekilde Hilton ve Loucks (2000)‟da egzersizin leptin üretimini baskılamadığı sonucuna varmışlardır. Bir başka çalışmada ise akut egzersizden 9 saat sonra leptin düzeylerinde bir azalmaya dikkat çekilmektedir (Nindl ve ark 2002). Buna karşın uzun mesafe yüzücülerinde veya maraton koşucularında yoğun egzersizi takiben leptin düzeylerinde anlamlı azalmaların meydana geldiğini bildiren raporlar da söz konusudur (Karamouzis ve ark 2002, Zaccaria ve ark. 2002) Benzer şekilde Pagano ve ark. (1999)‟ı 30 dakikalık akut bir yüzme egzersizinin ratlarda leptin düzeylerini yaklaşık %30 oranında azalttığını bildirmişlerdir.
Gerçekleştirdiğimiz çalışmada akut yüzme egzersizinden hemen sonra plazma leptin düzeylerinin kontrol grubuyla mukayese edildiğinde önemli ölçüde azaldığı gözlendi. Egzersizin leptin düzeylerini değiştirmediği (Hilton ve Loucks 2000) veya akut egzersizin leptin düzeylerini vücut yağ kitlesinden bağımsız olarak değiştirmediği (Hickey ve Calsbeek 2001) ileri sürülmektedir. Buna karşın Nindl ve ark. (2002) akut yoğun egzersizden 9 saat sonrasına kadar leptin konsantrasyonunda bir değişiklik olmadığını, 9 saat sonra ise leptin düzeylerinde önemli azalmalar meydana geldiğini göstermişlerdir. Benzer şekilde Van Agel-Leijssen ve ark. (1999) egzersizin 24 saatlik leptin piki ve ortalama konsantrasyonunu azalttığını bildirmişlerdir. Bahsedilen çalışmaların sonuçları, gerçekleştirdiğimiz çalışmada elde edilen bulgularımızla çelişkilidir. Ancak uzun mesafe yüzücülerinde egzersizden sonra leptin düzeylerinde önemli bir baskılanma olduğunun (Karamouzis ve ark. 2002), maratoncularda da benzer şekilde leptin düzeylerinde önemli bir baskılanma olduğunun ortaya konulması (Zaccaria ve ark. 2002) akut egzersizden hemen sonra elde ettiğimiz azalmış leptin düzeylerini destekleyen önemli raporlardır. Zaccaria ve ark. (2002) maratoncularda elde ettikleri azalmış leptin düzeylerine paralel olarak, serbest yağ asidi düzeylerinde de önemli bir artışa dikkat çekerek, azalmış leptin düzeylerini serbest yağ asidi düzeylerindeki artışa bağlamışlardır. Bizim akut yüzme egzersizini takiben elde ettiğimiz azalmış leptin düzeylerini destekleyen en önemli bulguyu Pagona ve ark. (1999) rapor etmişlerdir. Bahsedilen araştırıcılar (Pagona ve ark. 1999) çalışmalarında, ratlarda yaptırdıkları 30 dakikalık tek bir yüzme egzersizinden sonra plazma leptininde yaklaşık %30 civarında bir azalma olduğunu bildirmişlerdir. Ayrıca Baltaci ve ark. (2012) tarafından akut yüzme egzersizi yaptırılan sıçanlarda leptin düzeylerinin önemli ölçüde baskılandığının rapor
30
edilmesi de, çalışmamızda elde ettiğimiz azalmış leptin düzeyleriyle kuvvetli bir uyum gösterir.
Gerçekleştirdiğimiz çalışmada resveratrol uygulaması yüzme egzersizi yaptırılan sıçanlarda (grup 4) plazma leptin düzeylerini, resveratrol uygulanmayan yüzme grubuyla (grup 2) mukayese ettiğimizde etkilemedi. Biz çalışmamızda benzer olarak, kontrol grubu (grup 1) ve sadece resveratrol uygulanmış ve yüzme egzersizi yaptırılmamış grubun (grup 3) plazma leptin seviyeleri arasında da farklılık tespit edemedik. Sıçanlarda obezite ve diyabette gözlenen leptin direncinde resveratrol uygulamasının özellikle obezite tedavisinde leptin direncini düzelterek yararlı olabileceği bildirilmiştir (Franko ve ark. 2014). Resveratrol ve leptin ilişkisinde benzer bulgular Juman ve ark. (2011) tarafından da gösterilmiştir. Resveratrolun yağ dokusu üzerinde düzenleyici bir etkiye sahip olabileceği, resveratrol uygulamasının yağ dokuda azalmış leptin mRNA sentezinde artışa yol açabileceği ileri sürülmüştür (Eseberri ve ark. 2013). Bahsedilen her üç çalışmanın bulguları da, bizim resveratrol uygulamasıyle leptin düzeylerinde bir değişikliğin olmadığı şeklinde sonuçlarımızla uyumlu değildir. Ancak bahsedilen araştırmalar sıçanlarda obezite ve diyabette ortaya çıkan leptin direncinde resveratrol uygulamasının etkilerine odaklanmıştır. Bizim çalışmamız ise akut yüzme egzersizinde resveratrolun leptin salınımı üzerine olası etkisini araştırmayı amaçlamıştır. Gerçekleştirdiğimiz çalışmanın bulguları resveratrol uygulamasının hem akut yüzme egzersizinde, hem de egzersiz yaptırılmayan kontrol sıçanlarda plazma leptinini değiştirmediğini göstermektedir. Çalışmamızın bu kısmıyla ilgili bulgular resveratrol ve leptin ilişkisinde orijinal bir sonuç olarak kabul edilebilir.
4.1.2.Karaciğer Glikojen Düzeylerinin TartıĢılması
Gerçekleştirdiğimiz çalışmada en düşük karaciğer glikojen düzeyleri grup 2 (yüzme grubu)‟de elde edildi. Karaciğer glukoz dengesi ile lipit ve enerji metabolizmasında bir rol oynamaktadır (Fritsch eve ark. 2008). Akut ve tüketici bir egzersiz dokuların karbonhidrat ve lipit kullanımında defektler oluşturur ve sonuç olarak karaciğer glikojeni belirgin olarak azalır (Straczkowski ve ark. 2000). Sağlıklı bireylerde orta ve yüksek yoğunluktaki koşu egzersizinde artan kan glukozunun karaciğer kaynaklı olduğu (Petersen ve ark. 2004) gösterilmiştir. Zaten akut bir
31
egzersizi takiben karaciğer glikojeninde azalmasıyla ilgili görüş ayrılığı da söz konusu değildir (Baltaci ve ark. 2003; Akil ve ark. 2011). Çalışmamızda Grup 2 (yüzme kontrol)‟de elde ettiğimiz azalmış karaciğer glikojeni akut yüzme egzersizinin bir sonucu olarak kabul edilebilir. Çalışmamızda resveratrol uygulaması yaptığımız akut yüzme egzersizi yaptırılan sıçanların (grup 4) karaciğer glikojen düzeyleri grup 2 (yüzme grubu)‟ye oranla daha yüksekti. Elde ettiğimiz bu bulgu resveratrol uygulamasının yüzme egzersizinde azalan karaciğer glikojen depolarını koruyabileceğini göstermektedir. Resveratrol uygulamasının fiziksel aktivitede kas performansını artırabileceğine dikkat çekilmektedir (Scribbans ve ark. 2014). Karaciğer hücre yenilenmesi üzerine resveratrol uygulamasının olumlu etkilere yol açabileceği ve bu uygulamanın karaciğer glikojenini de sonuç olarak olumlu etkileyebileceği rapor edilmiştir (Ahmad ve Ahmad 2014). Kas yorgunluğu ve performans üzerinde resveratrol uygulamasının glukoz metabolizması üzerinde düzenleyici bir etkiye sahip olduğu ve kan laktat düzeylerini azaltarak yorgunluğu geciktirebileceği Wu ve ark. (2013) tarafından bildirilmiştir. Benzer bulgular Ryan ve ark (2010) tarafından yaşlı fareler üzerinde gösterilmiştir. Resveratrol ve egzersiz ilişkisinin araştırılması konusunda artan bir ilginin olduğu söylenebilir (Sun ve ark. 2012). Çalışmamızda yüzme grubuna (grup 2) oranla, resveratrol uygulaması yaptığımız yüzme grubunda (grup 4) elde ettiğimiz artmış karaciğer glikojeni önemli ve orijinal bir bulgudur. Fiziksel performansı ve resveratrol ilişkisinin araştırılmasında önemli bir basamak olarak kabul edilebilir. Gerçekleştirdiğimiz çalışmada en yüksek karaciğer glikojeni resveratrol uygulanan grupta (grup 3) elde edildi. Egzersizden bağımsız olarak elde ettiğimiz bu bulgu, resveratrol uygulamasının karaciğer hücre yenilenmesi (Ahmad ve Ahmad 2014) ve glukoz metabolizmasının düzenlenmesinde etkili olabileceğini (Wu ve ark. 2013) bildiren çalışmalarla uyumludur.
Çalışmamızın bulguları bir bütün olarak incelendiğinde resveratrol uygulamasının egzersiz yapan ve yapmayan sıçanlarda plazma leptini üzerinde önemli bir etkisinin olmadığı, yine resveratrol uygulamasının egzersiz yapan ve yapmayan sıçanlarda karaciğer glikojen depoları üzerinde koruyucu ve/veya düzenleyici bir etkiye sahip olduğunu göstermektedir.