LİYOFİLİZASYON İŞLEMİ ESNASINDA BAZI LAKTİK ASİT BAKTERİLERİNİN CANLILIKLARI ÜZERİNE KRİYOJENİK
KORUYUCU MADDELERİN ETKİLERİ
Yasemin PINARKARA
YÜKSEK LİSANS TEZİ GIDA MÜHENDİSLİĞİ
ANABİLİM DALI
i
LİYOFİLİZASYON İŞLEMİ ESNASINDA BAZI LAKTİK ASİT BAKTERİLERİNİN CANLILIKLARI ÜZERİNE
KRİYOJENİK KORUYUCU MADDELERİN ETKİLERİ
Yasemin PINARKARA
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Gıda Mühendisliği Anabilim Dalı Danışman: Yrd. Doç. Dr. Ahmet AYAR
2008, Sayfa: 106
Jüri: Yrd. Doç. Dr. Ahmet AYAR
Prof. Dr. Adem ELGÜN
Yrd. Doç. Dr. Mehmet AKBULUT
Bu çalışmada, Türkiye’nin çeşitli bölgelerinden (Konya, Kahramanmaraş,
Ereğli, Mut, Antalya, Ezine, Erzincan, Şanlıurfa) toplanan taze peynir ve yoğurt numunelerinden çeşitli laktik asit bakterileri izole edilmiş, bu bakteriler 12 farklı kriyojenik koruyucu maddenin (sakkaroz, laktoz, gliserol, Tween 80, maya ekstraktı, kazein peptonu, askorbik asit, trisodyum sitrat, monosodyum glutamat, malt ekstrakt, dimetil sülfoksit, jelatin) 4 farklı oranı (% 0.1, % 1, % 5, % 10) ile muamele edilerek liyofilize edilmiştir. 20 hafta boyunca -180 C’de depolanan bu liyofilize bakterilerin canlı hücre sayıları tespit edilerek, ilave edilen kriyojenik koruyucu maddelerin kültür canlılığı üzerine olan etkisi araştırılmıştır.
Araştırma sonuçları kriyojenik koruyucu madde ilavesinin kültür canlılığı üzerinde olumlu etkisi olduğunu göstermiştir. Farklı kriyojeniklerin farklı konsantrasyonları kültürler üzerine spesifik etkilerde bulunmuştur. 20 haftalık süre sonunda en olumlu etkiyi Str. thermophilus için % 5 pepton, Lc. lactis ssp. cremoris,
Lc. lactis ssp. lactis, Lb. casei, Lb. bulgaricus ve Lb. plantarum için % 10 trisodyum
sitrat, Lb. delb. ssp. lactis için % 0.1 askorbik asit, Lb. helveticus için % 5 maya ekstraktının gösterdiği tespit edilmiştir. Daha sonraki çalışmalarda bu kriyojenik koruyucu maddelerin farklı kombinasyonlarının etkisi üzerinde durulmasında fayda olabilir.
Anahtar Kelimeler: Kryoprotektan, Liyofilizasyon, Laktik Asit Bakterisi, Yoğurt,
ii
EFFECTS OF CRYOGENIC ADDITIVES ON THE SURVIVAL OF SOME LACTIC ACID BACTERIA IN LYOPHILIZATION
Yasemin PINARKARA Selcuk University
Graduate School of Natural and Applied Sciences Department of Food Engineering
Supervisor: Asist. Prof. Dr. Ahmet AYAR
2008, Pages: 106
Jury: Asist. Prof. Dr. Ahmet AYAR
Prof. Dr. Adem ELGÜN
Asist. Prof. Dr. Mehmet AKBULUT
In this study, different lactic acid bacteria were isolated from fresh cheese and yogurt that was collected from different areas of Turkey (Konya, Kahramanmaraş, Ereğli, Mut, Antalya, Ezine, Erzincan, Şanlıurfa). These bacteria mixed with 4 different ratio (% 0.1, % 1, % 5, % 10) of 12 different cryoprotective agent (sucrose, lactose, glycerole, Tween 80, yeast extract, peptone from casein, ascorbic acid trisodium citrate, monosodium glutamate, malt extract, dimethyl sulphoxide, gelatine) and then lyophilised. These lyophilised bacteria were stored at -180C for 20 weeks than colony forming units were evaluated to determine the best cryoprotectant.
Results showed that there is a positive effect of cryoprotectants on survival of cultures. Different ratios of different cryoprotectants have specific effects. As a result of statistical analyses, at the and of the 20 weeks storage time % 5 peptone from casein for Str. thermophilus, % 10 trisodium citrate for Lc. lactis ssp. cremoris, Lc. lactis ssp. lactis, Lb. casei, Lb. bulgaricus and Lb. plantarum, % 0.1 ascorbic acid for Lb. delb. ssp. lactis , % 5 yeast extract for Lb. helveticus were determined as cryoprotective agent for having the best survival rate.
iii
Mikroorganizmaların dondurularak kurutulması sonucunda elde edilecek canlılık ve aktivite üzerine en önemli etkiyi koruma ortamımın yaptığı konu ile ilgili hemen hemen tüm araştırmalarda önemle vurgulanmakta ve araştırıcılar farklı özelliklerde kryojenik katkıları önermektedirler.
Bu çalışmada Türkiye’nin çeşitli yörelerinden toplanan yoğurt ve peynir numunelerinden bakteri izolasyonu ve identifikasyonu yapılmış, seçilen bakteriler üzerinde 12 farklı kryojenik katkı 4 farklı oranda denenmiş ve sonuçlar istatistikî analiz yöntemi ile değerlendirilmiştir.
Bana bu konuda çalışma imkânı sunan, tez çalışmam esnasında bilgi ve deneyimleri ile bana öncülük eden, çalışmalarımı büyük bir özveri ile yürüten tez danışmanım Yrd. Doç. Dr. Ahmet AYAR’A, tez konusunun şekillenmesinde katkısı olan Prof. Dr. Necla ARAN’ a, baştan itibaren bu araştırmanın planlanıp yürütülmesinde benden hiçbir desteğini esirgemeyen Gıda Mühendisi Erdal ALSANCAK’a, yoğun laboratuar çalışmalarımda bana yardımcı olan İNTERMAK A.Ş. çalışanları Raziye TAGIL ve Burcu AKBULAK’a, istatistikî analizlerin yürütülmesinde katkıda bulunan Arş. Gör. Durmuş SERT’e ve her zaman yanımda olduklarını hissettiren, özveriyle bana destek olan aileme sonsuz teşekkürlerimi sunarım.
Ayrıca bu araştırmayı bir proje ile destekleyen Selçuk Üniversitesi Bilimsel Araştırma Projeleri Koordinatörlüğü’ne (S. Ü. BAP) çok teşekkür ederim.
iv ÖZET………….. ... i ABSTRAKT… ... ii ÖNSÖZ…..…... iii İÇİNDEKİLER... iv ŞEKİL LİSTESİ ... vi ÇİZELGE LİSTESİ ...xv 1. GİRİŞ………1 2. KAYNAK ARAŞTIRMASI... 3
2.1. Laktik Asit Bakterileri... 3
2.1.1. Lactobacillaceae Familyası... 5
2.1.1.1. Lactobacillus delbrueckii ssp. bulgaricus... 6
2.1.1.2. Lactobacillus delbrueckii ssp. lactis... 8
2.1.1.3. Lactobacillus helveticus... 9
2.1.1.4. Lactobacillus casei ...10
2.1.1.5. Lactobacillus plantarum ...11
2.1.2. Streptococcaceae Familyası ...12
2.1.2.1. Streptococcus salivarius ssp. thermophilus ...12
2.1.2.2. Lactococcus lactis ssp. lactis (Str. lactis) ...14
2.1.2.3. Lactococcus lactis ssp. cremoris (Str. cremoris) ...15
2.2. Liyofilizasyon (Dondurarak kurutma)...16
2.3. Kryojenik Katkılar (kryoprotektant, koruma ortamı)...18
3. MATERYAL VE METOT...24
3.1. Materyal ...24
3.2. Metot ...24
3.2.1. İzolasyon ve identifikasyon...24
v
3.2.1.3. İzolatların üretilmeleri ve saklanmaları ...27
3.2.1.4. İzole edilen suşların identifikasyonları ...28
3.2.2. Liyofilizasyon işlemi ...30
3.2.3. Canlı hücre sayımı ...31
3.2.4. İstatistik değerlendirme...32
4. ARAŞTIRMA SONUÇLARI VE TARTIŞMA ...33
4.1. Çeşitli Laktik Asit Bakterilerinin İzolasyonu ve İdentifikasyonu ...33
4.2. Kryojenik Maddelerin Streptococcus salivarius ssp. thermophilus Suşunun Canlılığı Üzerine Etkisi ...34
4.3. Kryojenik Maddelerin Lactococcus lactis ssp. cremoris Suşunun Canlılığı Üzerine Etkisi...43
4.4. Kryojenik Maddelerin Lactobacillus delbrueckii ssp. lactis Suşunun Canlılığı Üzerine Etkisi...51
4.5. Kryojenik Maddelerin Lactococcus lactis ssp. lactis Suşunun Canlılığı Üzerine Etkisi ...59
4.6. Kryojenik Maddelerin Lactobacillus plantarum Suşunun Canlılığı Üzerine Etkisi...67
4.7. Kryojenik Maddelerin Lactobacillus casei Suşunun Canlılığı Üzerine Etkisi.75 4.8. Kryojenik Maddelerin Lactobacillus helveticus Suşunun Canlılığı Üzerine Etkisi...83
4.9. Kryojenik Maddelerin Lactobacillus bulgaricus Suşunun Canlılığı Üzerine Etkisi...91
5. SONUÇ VE ÖNERİLER ...99
6. KAYNAKLAR ...101
vi
Şekil 2.1. Lactobacillus delbrueckii ssp. bulgaricus’un elektron mikroskobundaki
görüntüsü... 7
Şekil 2.2. Lactobacillus delbrueckii ssp. lactis’in Gram boyama yöntemiyle boyanmış görüntüsü... 8
Şekil 2.3. Lactobacillus helveticus’un elektron mikroskobundaki görüntüsü... 9
Şekil 2.4. MRS agar üzerinde gelişen Lactobacillus casei kolonileri ...10
Şekil 2.5. Lactobacillus casei’nin elektron mikroskobundaki görüntüsü ...10
Şekil 2.6. Lactobacillus plantarum’un Gram boyama yöntemiyle boyanmış görüntüsü...11
Şekil 2.7. Streptococcus salivarius ssp. thermophilus’un elektron mikroskobundaki görüntüsü...13
Şekil 2.8. Lactococcus lactis ssp. lactis’in Gram boyama yöntemiyle boyanmış görüntüsü...14
Şekil 2.9. Lactococcus lactis ssp. cremoris (Str. cremoris)’in elektron mikroskobundaki görüntüsü...15
Şekil 2.10. Denemelerde kullanılan liyofilizatör...18
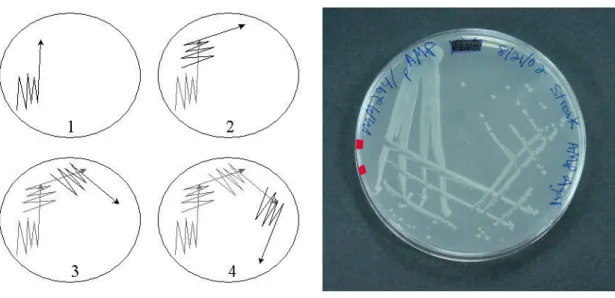
Şekil 3.1. Örneklerin izolasyon amacıyla ekimi...26
Şekil 3.2. Sürme tekniği ile saf koloni eldesi ...27
Şekil 3.3. Saf koloni eldesi...27
Şekil 3.4. API 50 CHL (Biomerieux,Marcy l’Etoile France) tanımlama kiti ...29
Şekil 3.5. Kültürel sayım yöntemi ...31
Şekil 4.2.1. Str. thermophilus suşunun farklı sukroz konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...39
Şekil 4.2.2. Str. thermophilus suşunun farklı laktoz konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...39
Şekil 4.2.3. Str. thermophilus suşunun farklı gliserol konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...39
vii
Şekil 4.2.5. Str. thermophilus suşunun farklı maya ekstraktı konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...40 Şekil 4.21.6. Str. thermophilus suşunun farklı pepton konsantrasyonlarında
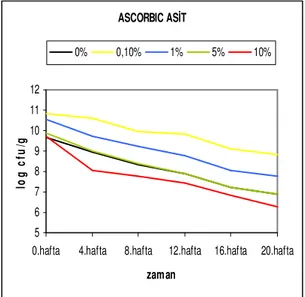
depolama süresine bağlı canlılık değişim grafiği ...40 Şekil 4.2.7. Str. thermophilus suşunun farklı ascorbic asit konsantrasyonlarında
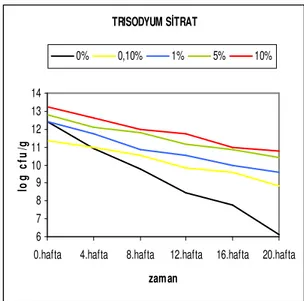
depolama süresine bağlı canlılık değişim grafiği ...40 Şekil 4.2.8. Str. thermophilus suşunun farklı trisodyum sitrat konsantrasyonlarında
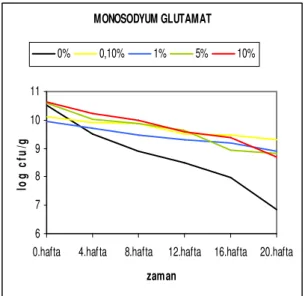
depolama süresine bağlı canlılık değişim grafiği ...40 Şekil 4.2.9. Str. thermophilus suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
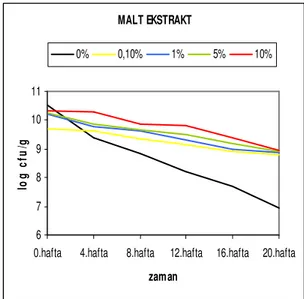
grafiği...41 Şekil 4.2.10. Str. thermophilus suşunun farklı malt ekstrakt konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...41 Şekil 4.2.11. Str. thermophilus suşunun farklı dimetil sülfoksit konsantrasyonlarında
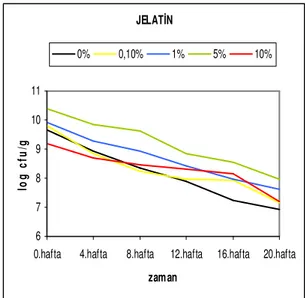
depolama süresine bağlı canlılık değişim grafiği ...41 Şekil 4.2.12. Str. thermophilus suşunun farklı jelatin konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...41 Şekil 4.3.1. Lc. lactis ssp. cremoris suşunun farklı sukroz konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...47 Şekil 4.3.2. Lc. lactis ssp. cremoris suşunun farklı laktoz konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...47 Şekil 4.3.3. Lc. lactis ssp. cremoris suşunun farklı gliserol konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...47 Şekil 4.3.4. Lc. lactis ssp. cremoris suşunun farklı Tween 80 konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...47 Şekil 4.3.5. Lc. lactis ssp. cremoris suşunun farklı maya ekstraktı
konsantrasyonlarında depolama süresine bağlı canlılık değişim
viii
Şekil 4.3.7. Lc. lactis ssp. cremoris suşunun farklı ascorbic asit
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...48 Şekil 4.3.8. Lc. lactis ssp. cremoris suşunun farklı trisodyum sitrat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...48 Şekil 4.3.9. Lc. lactis ssp. cremoris suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...49 Şekil 4.3.10. Lc. lactis ssp. cremoris suşunun farklı malt ekstrakt
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...49 Şekil 4.3.11. Lc. lactis ssp. cremoris suşunun farklı dimetil sülfoksit
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...49 Şekil 4.3.12. Lc. lactis ssp. cremoris suşunun farklı jelatin konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...49 Şekil 4.4.1. Lb. delbrueckii ssp. lactis suşunun farklı sukroz konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...55 Şekil 4.4.2. Lb. delbrueckii ssp. lactis suşunun farklı laktoz konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...55 Şekil 4.4.3. Lb. delbrueckii ssp. lactis suşunun farklı gliserol konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...55 Şekil 4.4.4. Lb. delbrueckii ssp. lactis suşunun farklı Tween 80
konsantrasyonlarında depolama süresine bağlı canlılık değişim
ix
grafiği...56 Şekil 4.4.6. Lb. delbrueckii ssp. lactis suşunun farklı pepton konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...56 Şekil 4.4.7. Lb. delbrueckii ssp. lactis suşunun farklı ascorbic asit
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...56 Şekil 4.4.8. Lb. delbrueckii ssp. lactis suşunun farklı trisodyum sitrat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...56 Şekil 4.4.9. Lb. delbrueckii ssp. lactis suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...57 Şekil 4.4.10. Lb. delbrueckii ssp. lactis suşunun farklı malt ekstrakt
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...57 Şekil 4.4.11. Lb. delbrueckii ssp. lactis suşunun farklı dimetil sülfoksit
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...57 Şekil 4.4.12. Lb. delbrueckii ssp. lactis suşunun farklı jelatin konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...57 Şekil 4.5.1. Lc. lactis ssp. lactis suşunun farklı sukroz konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...63 Şekil 4.5.2. Lc. lactis ssp. lactis suşunun farklı laktoz konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...63 Şekil 4.5.3. Lc. lactis ssp. lactis suşunun farklı gliserol konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...63 Şekil 4.5.4. Lc. lactis ssp. lactis suşunun farklı Tween 80 konsantrasyonlarında
x
Şekil 4.5.6. Lc. lactis ssp. lactis suşunun farklı pepton konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...64 Şekil 4.5.7. Lc. lactis ssp. lactis suşunun farklı ascorbic asit konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...64 Şekil 4.5.8. Lc. lactis ssp. lactis suşunun farklı trisodyum sitrat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...64 Şekil 4.5.9. Lc. lactis ssp. lactis suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...65 Şekil 4.5.10. Lc. lactis ssp. lactis suşunun farklı malt ekstrakt konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...65 Şekil 4.5.11. Lc. lactis ssp. lactis suşunun farklı dimetil sülfoksit
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...65 Şekil 4.5.12. Lc. lactis ssp. lactis suşunun farklı jelatin konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...65 Şekil 4.6.1. Lb. plantarum suşunun farklı sukroz konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...71 Şekil 4.6.2. Lb. plantarum suşunun farklı laktoz konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...71 Şekil 4.6.3. Lb. plantarum suşunun farklı gliserol konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...71 Şekil 4.6.4. Lb. plantarum suşunun farklı Tween 80 konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...71 Şekil 4.6.5. Lb. plantarum suşunun farklı maya ekstraktı konsantrasyonlarında
xi
Şekil 4.6.7. Lb. plantarum suşunun farklı ascorbic asit konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...72 Şekil 4.6.8. Lb. plantarum suşunun farklı trisodyum sitrat konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...72 Şekil 4.6.9. Lb. plantarum suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...73 Şekil 4.6.10. Lb. plantarum suşunun farklı malt ekstrakt konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...73 Şekil 4.6.11. Lb. plantarum suşunun farklı dimetil sülfoksit konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...73 Şekil 4.6.12. Lb. plantarum suşunun farklı jelatin konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...73 Şekil 4.7.1. Lb. casei suşunun farklı sukroz konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği...79 Şekil 4.7.2. Lb. casei suşunun farklı laktoz konsantrasyonlarında depolama süresine
bağlı canlılık değişim grafiği...79 Şekil 4.7.3. Lb. casei suşunun farklı gliserol konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...79 Şekil 4.7.4. Lb. casei suşunun farklı Tween 80 konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...79 Şekil 4.7.5. Lb. casei suşunun farklı maya ekstraktı konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...80 Şekil 4.7.6. Lb. casei suşunun farklı pepton konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...80 Şekil 4.7.7. Lb. casei suşunun farklı ascorbic asit konsantrasyonlarında depolama
xii
Şekil 4.7.9. Lb. casei suşunun farklı mono sodyum glutamat konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...81 Şekil 4.7.10. Lb. casei suşunun farklı malt ekstrakt konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...81 Şekil 4.7.11. Lb. casei suşunun farklı dimetil sülfoksit konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...81 Şekil 4.7.12. Lb. casei suşunun farklı jelatin konsantrasyonlarında depolama süresine
bağlı canlılık değişim grafiği...81 Şekil 4.8.1. Lb. helveticus suşunun farklı sukroz konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...87 Şekil 4.8.2. Lb. helveticus suşunun farklı laktoz konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...87 Şekil 4.8.3. Lb. helveticus suşunun farklı gliserol konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...87 Şekil 4.8.4. Lb. helveticus suşunun farklı Tween 80 konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...87 Şekil 4.8.5. Lb. helveticus suşunun farklı maya ekstraktı konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...88 Şekil 4.8.6. Lb. helveticus suşunun farklı pepton konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...88 Şekil 4.8.7. Lb. helveticus suşunun farklı ascorbic asit konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...88 Şekil 4.8.8. Lb. helveticus suşunun farklı trisodyum sitrat konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...88 Şekil 4.8.9. Lb. helveticus suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
xiii
Şekil 4.8.11. Lb. helveticus suşunun farklı dimetil sülfoksit konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...89 Şekil 4.8.12. Lb. helveticus suşunun farklı jelatin konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...89 Şekil 4.9.1. Lb. bulgaricus suşunun farklı sukroz konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...95 Şekil 4.9.2. Lb. bulgaricus suşunun farklı laktoz konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...95 Şekil 4.9.3. Lb. bulgaricus suşunun farklı gliserol konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...95 Şekil 4.9.4. Lb. bulgaricus suşunun farklı Tween 80 konsantrasyonlarında depolama süresine bağlı canlılık değişim grafiği ...95 Şekil 4.9.5. Lb. bulgaricus suşunun farklı maya ekstraktı konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...96 Şekil 4.9.6. Lb. bulgaricus suşunun farklı pepton konsantrasyonlarında depolama
süresine bağlı canlılık değişim grafiği ...96 Şekil 4.9.7. Lb. bulgaricus suşunun farklı ascorbic asit konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...96 Şekil 4.9.8. Lb. bulgaricus suşunun farklı trisodyum sitrat konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...96 Şekil 4.9.9. Lb. bulgaricus suşunun farklı mono sodyum glutamat
konsantrasyonlarında depolama süresine bağlı canlılık değişim
grafiği...97 Şekil 4.9.10. Lb. bulgaricus suşunun farklı malt ekstrakt konsantrasyonlarında
depolama süresine bağlı canlılık değişim grafiği ...97 Şekil 4.9.11. Lb. bulgaricus suşunun farklı dimetil sülfoksit konsantrasyonlarında
xv
Çizelge 2.1. Süt ürünlerinde starter bakteri olarak kullanılan Lactobacillus
genusuna giren türlerin ayırt edilmesinde önemli özellikler... 6 Çizelge 2.2. Bazı Streptococcus türlerinin ayrımında kullanılan test
toleransları ...12 Çizelge 3.1. MRS Agar bileşimi ...25 Çizelge 3.2. M17 Agar bileşimi ...25 Çizelge 4.1. Liyofilizasyon uygulamasında farklı kryojenik koruyucu maddelerin
genel olarak denemeye alınan laktik asit bakterilerinin canlılık
değerleri üzerine etkisi ...34 Çizelge 4.2. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
kriyojenik koruyucu maddelerin depolama süresince Streptococcus salivarius ssp. thermophilus suşunun canlılık oranına ait
istatistiksel analiz sonuçları………...………….36 Çizelge 4.3. Denemede kullanılan Streptococcus salivarius ssp. thermophilus
suşunun liyofilizasyon öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık
durumları...42 Çizelge 4.4. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
kriyojenik koruyucu maddelerin depolama süresince Lactococcus lactis ssp. cremoris suşunun canlılık oranına ait istatistiksel analiz sonuçları ...44 Çizelge 4.5. Denemede kullanılan Lactococcus lactis ssp. cremoris suşunun
liyofilizasyon öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık
durumları ...50 Çizelge 4.6. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
xvi
Çizelge 4.7. Denemede kullanılan Lactobacillus delbrueckii ssp. lactis suşunun liyofilizasyon öncesi canlı hücre sayılarına kıyasla ilk hafta
(liyofilizasyondan hemen sonra) ve 20. hafta % canlılık
durumları ...58 Çizelge 4.8. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
kriyojenik koruyucu maddelerin depolama süresince Lactococcus lactis ssp. lactis suşunun canlılık oranına ait istatistiksel analiz
sonuçları ...60 Çizelge 4.9. Denemede kullanılan Lactococcus lactis ssp. lactis suşunun
liyofilizasyon öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık
durumları...66 Çizelge 4.10. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
kriyojenik koruyucu maddelerin depolama süresince Lactobacillus plantarum suşunun canlılık oranına ait istatistiksel analiz
sonuçları ...68 Çizelge 4.11. Denemede kullanılan Streptococcus salivarius ssp. thermophilus
suşunun liyofilizasyon öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık
durumları ...74 Çizelge 4.12. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
kriyojenik koruyucu maddelerin depolama süresince Lactobacillus casei suşunun canlılık oranına ait istatistiksel analiz sonuçları ...76 Çizelge 4.13. Denemede kullanılan Lactobacillus casei suşunun liyofilizasyon
öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık durumları ...82 Çizelge 4.14. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
xvii
Çizelge 4.15. Denemede kullanılan Lactobacillus helveticus suşunun liyofilizasyon öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık durumları ...90 Çizelge 4.16. Liyofilizasyon denemesinde farklı konsantrasyonlarda kullanılan
kriyojenik koruyucu maddelerin depolama süresince Lactobacillus delbrueckii ssp. bulgaricus suşunun canlılık oranına ait istatistiksel analiz sonuçları...92 Çizelge 4.17. Denemede kullanılan Lactobacillus bulgaricus suşunun liyofilizasyon
öncesi canlı hücre sayılarına kıyasla ilk hafta (liyofilizasyondan hemen sonra) ve 20. hafta % canlılık durumları ...98
1. GİRİŞ
Mikrobiyoloji bilim dalının doğuşu ile birlikte, tabiatta çok yaygın olarak bulunduğu bilinen laktik asit bakterileri ile ilgili çalışmalar da başlamıştır. İlk kez 19. yüzyıl sonlarında sütte fermantasyona ve koagülasyona yol açan bakteriler laktik asit bakterileri olarak isimlendirilmiş ve daha sonraki yıllarda Lactobacillaceae familyası içinde sınıflandırılmışlardır (Çon ve Gökalp 2000).
Morfolojik açıdan çok değişken özellik gösteren (kısa veya uzun çubuk veya kok şekilli) familya üyeleri fizyolojik açıdan oldukça benzer özellik göstermektedirler. Tüm üyeler; Gram pozitif, katalaz negatif ( düşük oranda şeker ihtiva eden ortamda pseudokatalaza sahip suşlar görülebilir), Sporolactobacillus inulinus hariç spor oluşturmayan, fakültatif anaerobik, hareketsiz, çubuk veya kok şeklinde bakteriler olarak tanımlanmaktadır (Shape ve ark. 1966; Nickerson ve ark. 1974).
Mutlak fermantatiftirler ve asıl fermantasyon ürünü olarak laktik asit üretmektedirler. Doğal ortamları süt ve süt mamülleri, işlenmemiş taze veya çürümüş
bitkiler, insan ve hayvanların barsak mukozalarıdır (Schlegen 1986; Andersson 1989).
Laktik asit bakterileri, tabiatta yaygın oluşları, çeşitli gıda maddelerinde sıkça rastlanılan bozulmalara neden olmaları ve bazı gıdaların üretim ve olgunlaştırılmasında önemli rol oynamaları nedeni ile gıda teknolojisinde büyük önem taşımaktadırlar (Çon ve Gökalp 2000).
Bazı laboratuarlar çeşitli peynirler, tereyağı, yoğurt ve diğer fermente süt ürünleri için özel kültürler hazırlamaktadır. Bu kültürlerde yer alan mikroorganizmalar her saf kültürde değişiklik gösterebilir. Bunlar tek mikroorganizma türü olduğu gibi birkaç tür mikroorganizmayı da içerebilir. Bu kültürler; mezofil kültürler, termofil kültürler ve özel bakteri kültürleri
olabilmektedir. Değişik türden bakteri tür ve suşlarını içeren kültürler önceleri sıvı olarak üretilmiş ancak, bazı zararlı yönleri ve olumsuzlukları görüldüğünden ve özellikle de dondurma ve liyofilizasyon işlemlerinin endüstriyel boyutlarda kullanılması ile yerini dondurulmuş ve liyofilize kültürlere bırakmıştır (Yaygın ve ark. 2006).
Püskürterek kurutulmuş bakterilerde tatmin edici bir canlılık ve aktivite elde edilemediği, buna karşılık diğer kurutma yöntemi olan ve liyofilizasyon adı ile de bilinen dondurarak kurutmanın bakterilerin korunması açısından en iyi yöntemlerden biri olduğu belirtilmektedir (Stadhouders ve ark. 1969; Tamime ve Robinson 1976). Bilimsel araştırma ve endüstriyel amaçlı biyolojik örneklerin uzun süreli depolanması ve kullanım kolaylığı sağlaması gibi birçok avantaja sahip olduğu için liyofilizasyon işlemi kullanılmaktadır. Ancak liyofilizasyon işlemi dehidrasyon işlemi esnasında sitoplazma zarında meydana getireceği zararlar nedeniyle canlılar için ölümcül olabilmektedir (Lievense ve ark. 1994). Bu zararlı etkileri azaltmak için çeşitli şekerler (sakkaroz, laktoz, maltoz, trehaloz), şeker alkoller (inositol, sorbitol), aminoasitler (sodyum glutamat) kryojenik maddeler olarak kullanılmaktadır (Font de Valdez ve ark. 1983; Leslie ve ark. 1995; Abadias ve ark. 2001).
Bu çalışmada Türkiye’nin çeşitli bölgelerinden (Konya, Kahramanmaraş,
Ereğli, Mut, Antalya, Ezine, Erzincan, Şanlıurfa) toplanan taze peynir ve yoğurt numunelerinden çeşitli laktik asit bakterilerinin izole edilmesinin ardından bu bakterilerin farklı kryojenik maddelerle kombine edilerek liyofilize edilmesi ve 20 hafta boyunca -180 C’de depolanmış bu liyofilize bakterilerin canlı hücre sayılarının ölçülmesi amaçlanmıştır. Elde edilen bu sonuçların gerek araştırma laboratuarlarında gerekse endüstrinin ilgili kesimlerinde kullanılan mikroorganizmaların kültürel özelliklerini yitirmeden uzun süre korunmalarına yardımcı olacağı düşünülmektedir. Ayrıca Selçuk Üniversitesi Ziraat Fakültesi Gıda Mühendisliği bölümünde daha sonra yürütülecek olan laktik asit bakterileri ve starter kültürlerle ilgili çalışmalara da ön bilgi sağlayacaktır.
2. KAYNAK ARAŞTIRMASI
2.1. Laktik Asit Bakterileri
Laktik asit bakterileri çeşitli gıdalardaki faaliyetleri sonucu karbonhidratlardan
(heksozlardan) laktik asit üretme yeteneğine sahip mikroorganizmalardır. Bu mikroorganizmalar cins ve tür özelliklerine bağlı olarak karbonhidratlardan laktik asit yanında asetik asit, karbondioksit, alkol ve bazı tat ve aroma maddeleri de üretebilmektedirler. Bu maddelerin üretimiyle çok az da olsa gıdanın kalori değerinde bir değişim söz konusu olmaktadır. Ayrıca laktik asit bakterileri gıdaların bozulmasında rol oynayan mikroorganizmalar ve insanlarda hastalıklara neden olan patojen mikroorganizmalar üzerinde de ürettikleri asit ve bazı antimikrobiyal maddeler (bakteriosinler vb.) nedeniyle antogonistik etkiye sahiptirler. Bu nedenle laktik asit bakterilerinin faaliyetiyle üretilen fermente gıdalar gıda zehirlenmeleri ve enfeksiyonları düşünüldüğünde insan sağlığı açısından daha güvenilir gıdalar olarak kabul edilebilir (Ünlütürk ve Turantaş 2003)
Kluvyer laktik asit bakterilerini glikoz metabolizması sonucunda ürettikleri son ürüne bağlı olarak iki ana gruba ayırmıştır:
1) Heksoz şekerlerin fermentasyonu sonucu birincil derecede laktik asit üreten türlere homofermentatif laktik asit bakterileri adı verilmiştir. Homofermentatif laktik asit bakterilerinin homolaktik karakteri ortamın pH’ sı, besin maddeleri ve glikoz konsantrasyonu gibi kültürel koşulların değişmesiyle farklılık gösterebilir. Homofermentatif özellik heksoz şekerlerin metabolizmasında gözlenir. Bazı homofermentatif laktik asit bakterileri pentozları kullanarak asetik asit ve laktik asit üretebilirler. Bu gruba dahil laktik asit bakterileri heterofermentatif laktik asit bakterilerine kıyasla iki kat daha fazla enerji üretebilmektedirler.
2) Metabolik faaliyetleri sonucu heksozlardan laktik asit, CO2, ve etanol üreten
laktik asit bakterileri ise heterofermentatif laktik asit bakterileri olarak adlandırılırlar. Heterofermentatif türler asetaldehit ve diasetil gibi tat ve aroma maddelerinin üretimi açısından önem taşırlar.
Homofermentatif ve heterofermentatif laktik asit bakterileri genetik ve fizyolojik farklılıklarından dolayı glikozu farklı metabolik yollar izleyerek kullanmakta ve farklı son ürünler üretmektedirler. Homofermentatif laktik asit bakterileri heksoisomeraz (glukoz fosfat isomeraz) ve aldolaz enzimine sahipken, fosfoketolaz enzimini içermezler ve Embden-Meyerhof-Parnas (EMP) metabolik yolunu izleyerek bir molekül glikozdan iki molekül laktik asit üretirler. Heterofermentatif laktik asit bakterileri ise bunun tam tersi heksoisomeraz ve aldolaz enzimi yerine fosfoketolaz enzimine sahiptirler. Bu gruba giren laktik asit bakterileri glikozun yıkımında EMP metabolik izyolu yerine fosfoketolaz glikolitik izyolunu kullanırlar (Ünlütürk ve Turantaş 2003).
Laktik asit bakterilerinin taksonomik sınırları henüz kesin olarak çizilememiştir ve bu konuda değişik genetik çalışmalar sürdürülmektedir. Şu ana kadar tanımlanmış ve laktik asit bakterileri grubuna dahil edilmiş mikroorganizmaların baı genel özellikleri şöyledir:
a) Bu mikroorganizmalar gelişebilmek için aminoasitler ve B grubu vitaminler gibi besin ögeleri ile pürin ve pirimidin bazlarına ihtiyaç duyarlar.
b) Çoğu mezofilik mikroorganizmalardır. Ancak bazıları 50 C’nin altında, termofilik türler ise optimum 450 C gibi yüksek sıcaklıklarda gelişebilirler. c) Bu mikroorganizmaların çoğu optimum 4,0–4,5 pH’da gelişebilmelerine
rağmen bazıları 3,2 gibi düşük ve 9,6 gibi yüksek pH’larda da gelişebilmektedir.
d) Ayrıca bu mikroorganizmaların bazıları zayıf proteolitik ve lipolitik özelliğe sahiptir (Ünlütürk ve Turantaş 2003).
Burada sadece tez kapsamında kullanılan laktik asit bakterilerinin özellikleri üzerinde durulacaktır.
2.1.1. Lactobacillaceae Familyası
Lactobacillus Genusu ile temsil edilir. Çubuk şeklinde olan bu bakteriler düzgün çubuk, kokobasil veya uzun zincir oluşturan çubuk şeklinde bulunabilirler. Çubuk, bazı durumlarda kurve şeklinde veya filament uzantılı da olabilir. 0.5–2.0 mµ çaplı, 1.5–10.0 mµ uzunluğundadır. Çok z tür veya suşun dışında hareketsizdirler. Lactobacillus genusu türleri doğada oldukça fazla yaygındır. Örneğin silajda, hayvan ve inan bağırsağında bulunurlar. Normal süt florasında yer alırlar. Çiğ sütlerde bu genusa dahil birçok bakteriyi belirlemek mümkündür. Meydana getirdikleri süt asidi miktarı% 1–3 arasında değişir. Genellikle nitratı redükte etmezler. Katalaz enzimi üretmezler. Porfirin sistemleri yoktur. Laktik asidi fermente etmezler. Asetik asit, formik asit, suksinik asit, CO2, etil alkol ile iki karbonludan daha yüksek C’lu yağ
asidi oluşturamazlar (Kılıç 2001).
G(+) reaksiyon veren bakteriler, kültürlerin eskimesiyle G(-)reaksiyon verebilirler ve bu durumda uzun zincir formları gözlenir. Bu form geç logaritmik fazda da oluşabilir. Çoğunlukla hareketsizdirler. Spor oluşturmazlar anaerob veya mikroaerofildirler. Genellikle 5-530C arasında gelişebilirler. pH istemleri ise 5.5–5.8 arasında değişir. Patojen özellik göstermezler. Aksine oluşturdukları antibakteriyel özellikteki maddeler sayesinde saprofit ve patojen bakterilerin gelişmelerini engeller. 550C’nin üzerinde proteolitik etkinlikleri önemli ölçüde azalır. Bu özellikleri nedeniyle, yoğurt ve benzeri ürünlerin yapımında ve sert peynirlerin olgunlaştırılmasında kültür olarak kullanılırlar. Süt ürünlerinde tat ve aroma oluşumunda rolleri önemli düzeydedir (Kılıç 2001). Süt ürünlerinde starter bakteri olarak kullanılan Lactobacillus genusuna giren türlerin ayırt edilmesinde önemli özellikler Çizelge 2.1.’de verilmiştir.
Çizelge 2.1. Süt ürünlerinde starter bakteri olarak kullanılan Lactobacillus genusuna giren türlerin ayırt edilmesinde önemli özellikler (Desmazeaud 1992; Dellaglio ve ark. 1994)
Türler
Fermentasyon bulgaricus lactis helveticus acidophilus casei plantarum
Cellobiose - - - - + + Amygladine - - - + + + Galaktoz + + + + + + Glukoz + + + + + + Laktoz + + + + + + Maltoz - + + + + + Mannoz - + W + + + Salisin - + - + + + Trehaloz - + ± + + + Ramnoz - - - - - 150C - - - - + 450C + + + + - Eskülin hidroliz - W - + + Arginin’den NH3 - - - - Glikozdan gaz - - - -
+: %90 oranında pozitif, -: %90 oranında negatif, W: seyrek
2.1.1.1. Lactobacillus delbrueckii ssp. bulgaricus
Bu bakteri ilk olarak 1900’lü yıllarda Bulgar Grigoroff tarafından tanımlanmıştır. Morfolojik olarak Lb. delbrueckii ssp. lactis’ten ayırmak oldukça zordur. Oluşturduğu laktik asit miktarı, konsantrasyonu, DNA yapısındaki G+C oranı, hücre duvarı yapısı ve antijen grubu itibariyle hemen hemen aynıdır. Tek ayırıcı özellik maltozu kullanamamasıdır (Accolas ve ark. 1980).
Lb. delbruecki ssp. bulgaricus süt endüstrisinde çok önemli görevler üstlenmektedir. Yoğurt üretimi için hazırlanan saf kültürlerde Streptococcus salivarius ssp. thernophilus ile birlikte bulunur. Her iki bakteri arasında önemli bir ilişki söz konusudur. Bu ilişki proto-kooperasyon olarak isimlendirilmiştir. Bu olayda, yani yoğurdun oluşumu sırasında (her iki bakteriyi içeren kültürle aşılandıktan sonra) sütte önce Streptococcus salivarius ssp. thernophilus faaliyet göstererek laktozu parçalar ve az da olsa bir miktar (L+) laktik asit oluşturur, ortam pH’sını düşürür ve oksijeni kullanarak Lb. delbruecki ssp. bulgaricus için anaerob ortam yaratır. Bu arada Streptococcus salivarius ssp. thernophilus’un oluşturduğu folik asit, bu bakterinin gelişmesin hızlandırır. Lb. delbruecki ssp. bulgaricus ise kendisi için hazırlanan ortamda laktozu hızla parçalar ve % 1,8 oranında (D-) tipte laktik asit oluşturur. Optimum gelişme sıcaklığı 42-450C’ler arasındadır. DNA’da % G+C oranı 50.3 dolayındadır. Hücre duvarında peptidoglikan yapısı L-lisin, D-aspartat şeklinedir. Çiğ süt ve sert peynirlerde doğal olarak bulunması yanında (Fransa’da Grana, İsviçre’de Emmental, İtalyan peynirleri Mozzarella ve Taioggo gibi) sert, pıhtısı pişirilen peynirlerde kültür olarak kulanılmaktadır (Accolas ve Auclair 1983; Dellaglio ve ark. 1994).
Şekil 2.1. Lactobacillus delbrueckii ssp. bulgaricus’un elektron mikroskobundaki görüntüsü
2.1.1.2. Lactobacillus delbrueckii ssp. lactis
Morfolojik özelliği ve birçok fizyolojik özellik itibariyle Lb. delbruecki ssp. bulgaricus’tan ayırmak oldukça zordur. 2 mµ’dan daha kısa çaplı, iplik şeklindeki formlanmaya eğilimli yapısı tipiktir. Tekli veya çiftli çubuk şeklindedir. Homofermentatif olup % 1,8 oranında D(-) tipte laktik asit oluşturur. Optimum gelişme sıcaklığı 42-430C’dir. 150C’nin altında gelişmez, ancak 520C’de çoğalabilir. Metilen mavisi ile boyandığında granüllü yapısı görülebilir. Arginin’den NH3
oluşturur. Kuvvetli proteolitik özelliği nedeniyle pıhtısı yüksek sıcaklıkta pişirilen sert ve yarı-sert peynirlerin yapımında kültür olarak kullanılır. Gelişme ve çoğalmaları için besin madde gereksinimi çok yüksektir. Ca pantotenat, niasin, riboflavin gibi B kompleks vitaminler gelişmesini teşvik eder. DNA’daki %G+C oranı 50.2 dir. Hücre duvarında gliserol teichoic asit bulunur (Buchanan ve Gibbons 1974; Gasser ve ark. 1994).
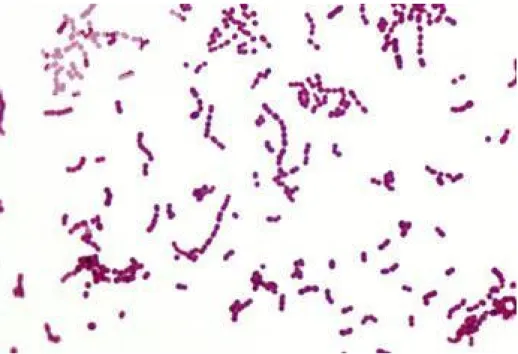
Şekil 2.2. Lactobacillus delbrueckii ssp. lactis’in Gram boyama yöntemiyle boyanmış görüntüsü
2.1.1.3. Lactobacillus helveticus
Lb. helveticus, ilk olarak 1919’da Orla-Jensen tarafından isimlendirilmiştir. Çiğ sütte, sert peynirlerde ve şirdenden hazırlanan peynir mayasında doğal olarak bulunur. Genellikle 0.6-1.0 µm çapında, 2-6 µm uzunluğunda tekli veya zincir şeklindedir. Homofermentatif olup DL formunda % 2.7 oranında laktik asit oluşturabilir. 40-420C’de optimum gelişme gösterir, ancak 150C’nin altında gelişemez, 530C’de ise çoğalma gösterir. Metilen mavisiyle metakromatik granülleri görünmez. Hücre duvarı yapısında gliserol teichoic asit bulunur. DNA’daki % G+C oranı 39.3’tür (Yaygın ve Kılıç 1993).
Zengin besiyerlerinde çok iyi gelişir. Maya ekstreli, fermente olabilen şekerler olduğu sürece iyi çoğalma gösterir. Niasin, Ca pentotenat, Pridoksamin, Mg++ gibi vitamin ve mineral maddelere gereksinimi oldukça yüksektir. Tween 80 veya Na’lu besiyerlerinde gelişmesi iyidir ve geniş ve buğulu koloni oluşturur (Tunail ve Köşker 1981).
2.1.1.4. Lactobacillus casei
Streptobacterium grubu içinde yer alır. 1.5 µm’den daha küçük çaplı ve uçları uzun veya kısa çubuk şeklinde, zincir oluşturabilen, flagellasız ve hareketsizdir. Homofermentatif bir bakteridir. Bazen pentozları kullanarak L+ laktik asit ve asetik asit de oluşturabilir. Ribozu laktik asit ve asetik asite CO2 çıkararak fermente eder; %
4 glukonatlı besiyerlerinde CO2 oluşturur. Optimum gelişme sıcaklığı 28-320C’dir.
150C’de ve onun altındaki sıcaklıklarda, bazı koşullarda 6-70C’lerde bile yaşamını sürdürür. Genellikle ekşi hamur, inek gübresi, insan ağız ve sindirim sistemi ile süt, peynir, sağım kapları ve makinalardan, kısaca sütün bulunduğu her yerden Lb. casei’yi izole etmek mümkündür. DNA’daki + G+C oranı 46.4 moldür (Kılıç 2001).
Şekil 2.4. MRS agar üzerinde gelişen Şekil 2.5. Lb. casei’nin elektron
2.1.1.5. Lactobacillus plantarum
Uç kısımları yuvarlak çubuk şeklinde genellikle 0.9-1.2 µm ende, 3-8 µ m uzunlukta, tekli, ikili veya kısa zincir şeklinde bulunur. Hareketlilik ve flagella normalde yoktur, fakat yan flagellalı suşlarda hareketlilik belirlenmiştir (Yaygın ve Kılıç 1993).
Anaerobik olup, yüzeyde gelişen kolonileri 3 mm çapta, yuvarlak, mat, kompakt, beyaz, seyrek olarak da açık veya koyu sarıdır. Gelişme ortamında yoğun bulanıklık oluşturur. Çoğu zaman α-metil-D-glukozit ve melezitoz’u fermente eder. Bazı suşları α-metil-D-mannozit’i fermente eder. DL laktik asit oluşturur. Fruktoz 1,6-difosfat aldolaz enzimine sahiptir. Glukonatlı besiyerinde CO2 oluşturarak
gelişir. Ribozu 1 mol laktik asit ve 1 mol asetik asite çevirir. Diğer pentozları da fermente eder (Weber 1996).
Genellikle 150C’de gelişir, 450C’de ise gelişme göstermez. Optimum gelişme sıcaklığı 30-350C’dir. Ca pantotenat ve niasin’e gereksinimi vardır. DNA’daki % G+C oranı 45’tir. Bulunuş yeri oldukça geniştir. Süt ürünleri, çevre, fermente olmuş bitkiler, silaj, turşu ile insan dudağı ve sindirim sisteminden izole edilebilir (Wegner 1981).
Şekil 2.6. Lactobacillus plantarum’un Gram boyama yöntemiyle boyanmış görüntüsü
2.1.2. Streptococcaceae Familyası
Sferik veya oval şeklindeki hücre tekli, ikili veya değişik uzunlukta zincir veya tetrat formu oluşturur. Hareketsiz veya nadiren hareketlidir. Endospor oluşturmaz ve G(+)’tir.
Kemoorganotrof yapıdadır. Karbonhidratlardan, fermentatif metabolizması sonucu laktik, asetik, formik asit ile alkol, karbonhidrattan ise CO2 oluşturur. Besin
gereksinimleri kompleks ve değişkendir. Katalaz testi genellikle negatif (-) olup bazen değişken sonuç verir. Fakültatif anaerobdur. Genel olarak DNA’daki % G+C oranı 33-44 arasındadır. Glukozu heksoz difosfat yoluyla fermente ederler ve L(+) asit oluştururlar (Deibel ve Seeley 1974). Bazı Streptococcus türlerinin ayırımında kullanılan test toleransları Çizelge 2.2.’ de verilmiştir.
Çizelge 2.2. Bazı Streptococcus türlerinin ayrımında kullanılan test toleransları (Buchanan ve Gibbons 1974). Türler 100C 450C %0.1 metilen mavisi %6.5 NaCl %40 safra tuzu pH 9.6 da gelişme 600C’ye 30 dk tolerans Str.thermophilus - + - - - - - Lc.lactis ssp. lactis + - + - + - d Lc.lactis ssp. cremoris + - d - + - d
+: %90 oranında pozitif, -: %90 oranında negatif, d: değişken sonuç
2.1.2.1. Streptococcus salivarius ssp. thermophilus
Küre veya oval olan hücreleri 0.7-0.9 µm çaplıdır. İkili veya uzun zincir oluşturur (Desmazeaud 1983). Glukoz buyyonunda son gelişme pH’sı 4.0-4.5’tur.
tercih ettiği disakkarit sakkaroz ve laktozdur. Glukoz, fruktoz, laktoz ve sakkarozdan sit üretir. Trehaloz, maltoz, inülin, gliserol, mannitol, sorbitol veya salisinden seyrek olarak asit üretirken; rafinoz, ksiloz veya arabinozdan asit üretmez. Sentetik besiyerinde seri transferlerde B kompleks vitaminlerden pantotenik asit ve riboflavine gereksinimi vardır. Pridoksin, biotin, tiamin ve niasin gelişmesini stimüle eder. Purin ve piridin’e gereksinim duymaz. Aminoasit gereksinimi yüksektir ve mevsimlere göre değişkenlik gösterdiği yapılan araştırmalarda ortaya konmuştur (Accolas 1979, Desmazeaud 1983).
Obligat homofermentatif özellik gösteren bakteri sitratı fermente etmez. Gaz oluşturmaz. Proteolitik ve lipolitik aktivitesi zayıftır. Asetaldehit, aseton, asetoin ve diasetil gibi aroma maddelerini oluşturur. Alkol, hidrojen peroksit ve vitamin üretimi (B6, B12) iz miktardadır. Sütü % 0.7-0.8 oranında L(+) laktik asit oluşturarak pıhtılaştırır. Optimum gelişme sıcaklığı 37-420C olarak bildirilmiştir. 500C’de gelişir, 530C’de ise gelişmez. 200C’nin altında gelişme göstermez. Sıcağa toleransı için rakam vermek gerekirse 630C’de 30 dakikadır. Süt ve süt ürünleri, İsviçre tipi peynir ve yoğurt kaynağını oluşturur. Bu ürünlerin yapımındaki kültürde kullanılır (Deibel ve Seeley 1974; Yaygın ve Kılıç 1993).
Şekil 2.7. Streptococcus salivarius ssp. thermophilus’un elektron mikroskobundaki görüntüsü
2.1.2.2. Lactococcus lactis ssp. lactis (Str. lactis)
1873’te Lister, Bacterium lactis olarak adlandırmıştır. Oval hücreleri 0.5-1.0 µ ve düzgündür. İkili formu kısa zincir formuna göre daha sıklıkla bulunur. Bazı suşları uzun zincir oluşturur. Son gelişme pH’sı glukoz buyyonunda 4.0-4.5’tur. sütte % 0.5-0.7 oranında L(+) laktik asit oluşturur. Proteolitik aktiftir. Mannitol, trehaloz, sakkaroz, arabinoz, ksiloz, laktoz, maltoz ve glukozdan asit üretir. Raffinoz, inülin, gliserol veya sorbitol’den asit oluşturmaz. Bazı suşlar nisin üretir (Hirsch ve Grimsted 1951; Hirsch ve ark. 1951). Birçok Gram (+) bakteriyi inhibe eder. % 4 NaCl’de gelişir. % 6.5’ta gelişmez. 9.2 pH’da gelişir, 9.6’da ise gelişme göstermez. % 0.3 metilen mavili sütte gelişir. Bazı türleri Lösin’i 3-metilbutanol’e dönüştürür ve malt tadı oluşturur. Besin istekleri çoktur. 4-5 vitamin, 10-13 aminoasidi sentetik besiyerlerinde isterler. Purin ve pirimidin’i stimülatör olarak kullanırlar. Optimum gelişme sıcaklıkları 300C’dir. Bazı türler 410C’de gelişmez, 450C’de ise hiç gelişme göstermez. Süt ürünlerinde adil kontaminanttır. DNA’sındaki % G+C oranı 38.4-38.6 mol’dür (Kılıç 2001).
Şekil 2.8. Lactococcus lactis ssp. lactis’in Gram boyama yöntemiyle boyanmış görüntüsü
2.1.2.3. Lactococcus lactis ssp. cremoris (Str. cremoris)
Hücreler yuvarlak ve oval şeklinde düz bir hat boyunca genellikle uzun zincir olacak şekilde sıralanır. Hücre çapları 0.6-1 µm dolayındadır. Sütte uzun zincir şeklinde bulunduğu gibi bazı kültürlerde predominant olarak ikili bulunur (Buchanan ve Gibbons 1974).
Son pH aralığı glukoz buyyonu için 4-4.5’tur. Sütte % 0.5-0.7 oranında L(+) laktik asit oluşturarak sütü pıhtılaştırır. Proteolitik aktiftir. Az miktarda asetoin, asetaldehit ve diasetil oluşturur. Glukoz ve laktozdan asit üretir. Seyrek olarak maltoz, sakkaroz, raffinoz’u fermente eder. Arabinoz, ksiloz, inülin, gliserol ve sorbitolü ise fermente etmez. Ortamda fermente edilebilir şeker bulunduğunda bazı suşları sitrat’ı parçalar ve CO2, asetik asit ile diasetil oluşturur. Bazı suşlar
antibiyotik benzeri maddeler üretirler (Dellaglio ve ark. 1994; Tunail ve Köşker 1981). Optimum gelişme sıcaklığı 28-330C’dir. 400C’de ise gelişmez. Çiğ süt ve süt ürünleri kaynağını oluşturur. Lc. lactis ssp. lactis’ ten ayırımı, arginin’den NH3
oluşturması ile olur. Genellikle % 4 NaCl’lü buyyonda gelişir. % 0.3 metilen mavili sütte ve 9.2 pH’ya ayarlanmış ortamda gelişemez. DNA’sındaki % G+C oranı 38-40 mol’dür. Bazı suşları polisakkarit oluşturur ve hidrojen peroksit üretir.
Şekil 2.9. Lactococcus lactis ssp. cremoris (Str. cremoris)’in elektron mikroskobundaki görüntüsü
2.2. Liyofilizasyon (Dondurarak kurutma)
Dondurarak kurutma terimi; genellikle biyolojik kaynaklı olan nemli
materyallerin ya da sulu çözeltilerin değişikliğe uğramadan kurutularak korunması anlamına gelmektedir. Ürün dondurulur ve daha sonra düşük nispi neme sahip bir atmosfere maruz bırakılarak üründe oluşturulan buz kristallerinin süblimasyonu sağlanır. Pratikte gerekli olan düşük nispi nem, vakum oluşturularak sağlanır. Oluşan gözenekli kek vakum altında kurutulur ve raf ömrünün uzun olması amacıyla oksijensiz ve nemsiz ortamda paketlenir. Oluşan ürün; dondurulan materyalle aynı şekle ve boyutlara sahiptir, kullanılıncaya kadar oda sıcaklığında aylarca hatta yıllarca depolanabilir. Elde edilen ürün uygun miktar ve kalitedeki su ile gerektiğinde tekrar çözelti haline getirilebilir (Rowe ve Snowman 1976).
Dondurarak kurutma konusundaki bilinen en eski çalışma 1890 yılında Altman
tarafından yapılandır. Bu araştırıcı vakumlu desikatörde -20 0C’de donmuş doku parçalarını kurutmuştur. Daha sonraki yıllarda liyofilizasyon tekniğinde belirli aşamalar kaydedilmiş, ortamdan uzaklaştırılması gereken nemin kimyasal maddeler yerine soğuk yüzeylere tutunması üzerine çalışmalar yapılmış ve bu konu ile ilgili yayınlanan ilk patentte 1934 yılında Esler kuru karbondioksitli nem tutucuyu açıklamıştır. 1936 yılında Greaves ve Adair, 1939 yılında Greaves kan plazmasının kurutulması sırasında oluşan köpürme sorununu çözerek bilim ve mühendislik temellerini beraberce kullanmışlar ve buhar basıncı, vakum parametrelerini açıklamışlardır. 2. Dünya Savaşında fazla miktardaki plazma gereksinimini karşılamak amacıyla dondurarak kurutma konusundaki çalışmalar yoğunlaşmış, bunun sonucunda yeni cihaz modelleri ve yeni yöntemler geliştirilmiştir (Meryman 1966).
Dondurarak kurutma genel olarak 3 aşamaya ayrılır. 1) ön dondurma
2) birincil kurutma (vakum altında süblimasyonla kurutma) 3) ikincil kurutma
Kurutmadan önceki dondurma işlemi karbondioksit, alkol, buz, tuz, dielektrik metal bloklar kullanmak gibi çeşitli şekillerde yapılabilir. Bu işlem bir evaporatif dondurarak kurutma makinasının bir bölmesinde de gerçekleşebilir. Burada materyalden dışarı çıkan su buharının gizli evaporasyon ısısı dondurmaya yeterlidir (Lapage ve ark. 1970)
Birincil kurutma aşamasında donmuş materyalin tümündeki buz kendiliğinden buharlaşamaz ancak donmuş arayüz sürekli olarak dıştaki kuru kısıma doğru hareket eder. İkincil kurutmada ise ürün optimum su içeriğine sahip oluncaya kadar, birincil kurutma sonrasında kalan bir miktar su da uzaklaştırılır (Rowe ve Snowman 1976). Liyofilizasyon makinelerinin en basit şekli bir vakum pompasına bir desikatörün ilavesi ile yapılabilir. Vakum odasında hızlı evaporasyon ve buna bağlı olarak çabuk soğuma sağlanabilir. Burada desikatörün görevi ortamdan uzaklaştırılan su buharını tutmaktır. Bu amaçla fosfor pentaoksit gibi yüksek düzeyde nem tutan kimyasal maddeler veya gelişmiş sistemlerde olduğu gibi soğutmalı kondansör kullanılabilir (Rowe 1960).
Materyal yeteri kadar donmadan yüksek vakum uygulanırsa köpürme oluşur ve materyal kaptan dışarı çıkar. Bu sorun kullanılan liyofilizatörün özelliğine göre şu şekilde çözülebilir: ilk olarak materyal önce -30- -40 0 C’ de dondurulur ve sonra vakum sistemine bağlanır. Santrifüjlü liyofilize sistemlerinde ise bu sorun kendiliğinden çözülmüştür. Vakum odası içinde bir santrifüj başlığına dışa doğru eğik olarak yerleştirilen tüpler içindeki materyal donma aşamasında eğime ve dönüş hızına bağlı olarak merkezkaç kuvveti etkisi ile aşağı doğru bastırılır. Bu sistemlerde santrifüjün iki fonksiyonu daha vardır. Bunlar; materyalin tüp çeperine yapışması ile geniş bir yüzey sağlayarak evaporasyonu hızlandırmak ve santrifüjün dönüşü sırasında vakum odasında az da olsa sürtünmeden gelen bir sıcaklık oluşturarak yine evaporasyonu hızlandırmaktır (Robson ve Rowe 1960).
Gelişen teknolojiye bağlı olarak bugün pek çok biyolojik materyal çeşitli amaçlarla liyofilize edilmektedir. Mikroorganizmalardan birçok bakteri, maya, küf,
virüs, bazı algler ve birkaç protozoa cinsi yanında başta BCG olmak üzere pek çok aşı, enzimler ve proteinler, eritrosit, spermatozoa, organ naklinde kullanılmak üzere kornea, serum plazma, diğer biyolojik materyal ve gıdalar liyofilizasyon tekniği ile korunabilmektedir (Meryman 1966; Lapage ve ark. 1970; Sawada 1975; Cemeroğlu 1976).
Şekil 2.10. Denemelerde kullanılan liyofilizatör.
2.3. Kriyojenik Koruyucu Maddeler (kriyoprotektan, koruma ortamı):
Mikroorganizmaların dondurarak kurutulması sonucunda elde edilecek canlılık
ve aktivite üzerinde en önemli etkiyi kriyojenik katkıların yaptığı konu ile ilgili hemen hemen tüm araştırmalarda önemle vurgulanmakta ve araştırıcılar farklı özelliklerde kriyoprotektanlar önermektedirler. Dondurarak kurutma sırasında hücrelerde oluşan zararların biyokimyasal mekanizması henüz tam olarak açıklığa kavuşamadığı için canlılık ve aktivitenin korunmasında en önemli faktör olduğu
kabul edilen koruyucu maddelerin (kriyoprotektan katkılar) koruyucu mekanizmaları da tam olarak anlaşılmamıştır. Yoğurt bakterilerinin liyofilizasyonu üzerine yapılan çeşitli araştırmalarda liyofilizasyondan hemen sonra % 100 canlılık elde edilmesine karşılık ölümlerin depolama süresi uzadıkça artması sonucu kriyoprotektanlar üzerindeki çalışmalar yoğunlaşmıştır (Halkman ve ark. 1986).
Yukarıda da değinildiği gibi liyofilizasyon sırasında meydana gelen canlılık ve aktivite kayıplarının nedeni bugüne kadar kesin olarak açıklanamamış olduğundan bu kayıpları azaltmaya yönelik önlemler de araştırıcıların kişisel görüşlerini yansıtmaktan öteye gidememektedir. Dolayısı ile canlılık ve aktivitenin korunmasında en önemli faktör olduğu pek çok araştırıcı tarafından kabul edilen kriyoprotektanların koruma mekanizmaları da açıklanamamıştır.
Kriyoprotektanların yapıları ve etkileri konusunda en ayrıntılı bilgilere Japon araştırıcı Morichi ve arkadaşlarının çalışmalarında rastlanmıştır. Morichi ve ark. (1963) glutamik asit ve ilgili bileşenlerinin dondurarak kurutulmuş bakteri hücreleri üzerindeki etkilerini araştırdıkları bir çalışmalarında 45 bileşiğin koruyucu etkisini 26 farklı bakteri üzerinde denemişlerdir. Araştırıcılar bu bileşiklerin koruyucu etkilerinin fizikokimyasal yapılarından kaynaklandığını ancak metabolizma ile ilgilerinin bulunmadığını belirtmişlerdir. Bu araştırıcılar koruyucu etkileri olan bazı bileşiklerin hücre tarafından metabolize edilemediğini, aynı şekilde metabolize edilebilen bazılarının ise koruyucu etkilerinin bulunmadığını, glutamik asitin koruyucu etkisinin optik aktivitesi ile değişmediğini göstererek bu görüşlere varmışlardır. Yine Morichi ve ark. (1965) tarafından yapılan benzer bir çalışmada araştırıcılar bu kez 44 farklı bakteri üzerine arjinin ve ilgili bileşikleri olan 14 koruma ortamı denemişlerdir. Araştırma sonuçlarından;
a) Arjinin’in α-COOH, α-NH2 ve guanidino gruplarının etkili olduğunu,
b) Arjinin içine metilen katılmasının koruyucu özellikte önemli etki yapmadığını
c) Guanidino grubunun –NH2 ye dönüşmesinin koruyucu etkiyi önemli ölçüde
d) Arjinin molekülünün α-NH2 grubunun –OH grubu ile koruyucu etkide bir
farklılık olmadan yer değiştirebileceğini
e) Arjininin koruyucu etkisinin optik aktivitesinden bağımsız olduğunu bulan araştırıcılar bu görüşler doğrultusunda etkili bileşenlerin kimyasal yapılarının üç ya da daha fazla hidrojen bağlayıcının ve/veya uygun konsantrasyondaki iyonize edicinin varlığı ile karakterize edilebileceğini öne sürmüşlerdir. Morichi (1975) ise 24 farklı şeker ve polialkol arasında liyofilize bakterilerde canlılık üzerinde en büyük etkiyi disakkaritlerin yaptığını, bunu aldoheksozların ve diğer bileşenlerin izlediğini belirtmiştir.
Kriyojenik koruyucu maddelerin etkileri konusunda yorum getiren başka araştırıcı grubu Sinha ve arkadaşlarıdır. Sinha ve ark. (1972) 11 koruyucu ortamı S.
lactis suşuna karşı denemişler ve bu koruyucu ortamlar arasında görülen etki
farklarını yorumlamışlardır. Bu araştırıcılar hücre içi suyun aşırı derecede uzaklaştırılması halinde hücre ölümlerinin artacağını, iyi bir koruma ortamı olan yağsız süt ortamının hücre içi suyun kalmasına yardım edeceğini, yağsız süt besiyerinin askorbik asit, tiyoüre ve amonyum klorür ile desteklenmesinin canlılığı artırdığını, buna karşılık destek maddelerinin tek başlarına kullanılması ile yeteri kadar canlılık elde edilemediğini ve dolayısı ile koruma ortamlarının canlılık artırmadaki yararlı etkilerinin birden fazla mekanizmaya bağlı olduğunun sanıldığını belirtmişlerdir. Araştırıcılar ayrıca koruma ortamlarının etkisinin oksijene duyarlı kısımların bağlanması ile ilgili olduğunu, yağsız süt besiyerini desteklemede kullanılan maddelerin bu kısımlardaki oksijenin toksik etkisini azaltmada kullanıldığını, askorbik asidin antioksidan etkilerinin bilindiğini ve bu maddenin tiyoürenin ortamda bulunması ile stabilitesini kazandığını, amonyum klorür varlığında muhtemelen benzer etkiye sahip olduğunu ancak kesin etkisinin bilinmediğini belirtmişlerdir. Bu araştırıcılara göre iyi bir koruma ortamı olan sodyum glutamatın canlılığı korumadaki yüksek etkisi suya karşı gösterdiği affiniteden ve aynı zamanda donmuş gıdalarda görülen antioksidan özelliğinden gelebilir.
Özalp ve Özalp (1978) tarafından Etlik Veteriner Araştırma ve Kontrol Enstitüsünde liyofilize yoğurt kültürü üretmek üzere yapılan bir çalışmada kültürler % 1.5 sodyum glutamatın kriyojenik koruyucu madde olarak kullanılması ile liyofilize edilmişlerdir. Türkiye’de liyofilize yoğurt kültürü yapılan 2. yer olan Ege Üniversitesi Ziraat Fakültesinde ise herhangi bir kryojenik katkı kullanılmamaktadır. Kılara ve ark. (1976) denedikleri 12 kryoprotektan arasında L. bulgaricus için liyofilizasyondan hemen sonra % 10 malt ekstraktın % 54, 0.1 M laktozun % 56 canlılık sağladığını, 48 haftalık depolama sonunda her iki koruma ortamının % 1’ in altında canlılık verdiğini, buna karşılık başlangıçta % 33 canlılık veren % 10 oranındaki kasitonun 48 hafta sonunda % 15 canlılık sağladığını, diğer 11 kriyojeniğin bu süre sonunda %1’in altında canlılık verdiğini tespit etmişlerdir.Aynı koruma ortamlarını S. thermophilus için deneyen bu araştırıcılar tarafından bu bakteri için % 10 malt ekstrakt çözeltisi, başlangıçta % 60, 48 haftalık depolamadan sonra % 30 canlılık sağlayarak en iyi koruma ortamı olarak bulunmuştur.
Morichi ve ark. (1965) arjinin ve ilgili bileşenleri olan çeşitli koruma ortamları arasında dondurarak kurutma işleminden hemen sonra olmak üzere Lb.
bulgaricus’un 3 suşu için pH 7’deki L-arginin ile % 20, % 20, % 35, Str.
thermophilus’un 2 suşu için pH 7 olan 0.06 M DL-malik asit ile % 52 ve % 59 canlılık elde etmişlerdir. Morichi ve ark. (1963) Lb. bulgaricus için glutamik asit ve ilgili bileşenleri olan L-glutamin, DL-norvalin, γ amino bütirik asit ve glutarik asit arasında liyofilizasyondan hemen sonra olmak üzere pH 7 deki 0.06 M L-glutamik asidin % 11 canlılık ile diğerlerinden daha iyi sonuç verdiğini göstermişlerdir. Morichi (1975) ise yine dondurarak kurutma işleminden hemen sonra Lb. bulgaricus için laktoz ile % 32, sakaroz ile % 38, arjinin hidroklorid ile % 37, % 1 arjinin içeren yağsız süt besiyeri ile % 85, Str. thermophilus için % 1 sodyum malat içeren yağsız süt besiyeri ile % 80-100 arasında canlılık elde etmiştir. Morichi (1974) bir başka araştırmasında ise yoğurt kültürlerinin liyofilizasyondan hemen sonra en yüksek canlılığa sırası ile her biri 0.06 M olmak üzere glutamat, malat, arjinin ile desteklenmiş yağsız süt besiyeri ile ulaştığını belirlemiştir. Yine Morichi ve ark. (1972) bu kez Str. thermophilus için pH 6.6-6.8 deki % 1 malat içeren % 8 yağsız süt
besiyeri ile dondurarak kurutmadan hemen sonra % 80 den fazla canlılık elde ettiklerini, 380 C’ de 6 ay depolamadan sonra ise canlılığın hala tatmin edici düzeyde olduğunu belirtmişlerdir.
Sinha ve ark. (1974) yaptıkları bir seri çalışma sonunda geliştirdikleri pH’sı 7 olan yağsız süt besiyeri ve her biri % 0.5 olmak üzere askorbik asit, tiyoüre ve amonyum klorür karışımının Lb. bulgaricus ‘un 2 suşunda liyofilizasyondan hemen sonra % 13.7 ve % 19.8 vakum altında 2 ay depolanmış kültürlerde % 6.1 ve % 9.3 normal atmosfer basıncında 2 ay depolanmış kültürlerde % 1.2 ve % 2.9 canlılık elde etmişlerdir. Aynı koruma ortamı ile Str. thermophilus ‘un 3 suşu için liyofilizasyon sonunda % 95.5 ; % 76.6 ; % 92.3, vakum altında 2 ay depolama sonunda % 79 ; % 9.8 ; % 58.9 normal atmosfer basıncında 2 ay depolamada ise % 53.1 ; % 5.5 ; % 7.9 canlılık sağlanmıştır.
Lagoda ve Bannikova (1970, 1975a, 1975b) Lb. bulgaricus için bileşimi % 5 sodyum asetat, 2 atmosfer basınç altında 2 saat otoklavlanmış % 5 jelatin, % 2 sodyum glutamat ve % 10 sakkaroz karışımı olan koruma ortamı ile % 78; % 82 , Str. thermophilus için bileşimi % 5 sodyum sitrat, 2 atmosfer basınç altında 2 saat otoklavlanmış % 5 jelatin, % 2 sodyum glutamat ve % 10 sakkaroz karışımı olan koruma ortamı ile % 85; % 96 canlılık elde etmişlerdir.
Lapage ve ark. (1970) Lb. bulgaricus için steril at serumu, nutrient broth, glukoz karışımı, Str. thermophilus için steril at serumu ve nutrient broth karışımını koruma ortamı olarak önermişlerdir.
Gavin (1968) oda sıcaklığında ve tropik koşullarda (350 C) depolanacak liyofilize yoğurt kültürleri için yağsız süt besiyeri, % 10 sakkaroz ve % 1-2 glutamat karışımının iyi sonuçlar vereceğini belirtmiştir.
Porubcan ve Sellars (1975) 2 litre % 12 yağsız süt besiyerinde ürettikleri Lb. bulgaricus kültürü üzerine 40 g askorbik asit, 25 g monosodyum glutamat ve 25 g inositol katmışlar, pH % 50 NaOH ile 6.1’e ayarlanmış ve kültürü liyofilize
etmişlerdir. Araştırıcılar 210 C’ de 3 ay depolama sonunda yüksek canlılık aldıklarını bildirmişlerdir.
Mitic ve Otenhajmer (1974) % 0.3 maya ekstraktı ile zenginleştirilmiş yağsız süt besiyerini Str. thermophilus için, % 0.2 pankreatik ekstraktlı yağsız süt besiyerini Lb. bulgaricus için en iyi koruma ortamları olarak göstermişlerdir.
Carvalho ve ark. (2003) 5 farklı türdeki laktik asit bakterisinin liyofilizasyonunda yağsız süt ortamına % 1.25 oranında ilave edilen sorbitol ve mono sodyum glutamatı kryoprotektant olarak denemişler ve liyofilizasyondan hemen sonra monosodyum glutamat ilave edilen Lb. plantarum ve Lb. bulgaricus suşları için sırasıyla 8.62 ve 7.08 log cfu/ml değerlerini elde etmişlerdir.
Champagne ve Gardner (2001) Streptococcus thermophilus’un 2 suşunun liyofilizasyonu esnasında pepton, laktoz, trehaloz ve peyniraltı suyu-sakkaroz karışımını denemişlerdir ve liyofilizasyondan hemen sonra sırasıyla % 0.6, % 17, % 31, % 50 canlılık elde etmişlerdir.
De Giulio ve ark. (2005) yaptıkları araştırmada Lb. acidophilus, Lb. bulgaricus ve Str. thermophilus’un liyoflizasyonunda koruma ortamı olarak trehaloz, maltoz, sakkaroz, glikoz ve laktozu kullanmışlar ve liyofilizasyon sonrası canlılıklarını belirlemişlerdir. Bu araştırmanın sonucuna göre Lb. bulgaricus ve Str. thermophilus suşları için sırasıyla trehaloz ortamı % 95, % 87, maltoz ortamı % 90, % 81, sakkaroz ortamı % 95, % 82, glikoz ortamı % 86, % 18, laktoz ortamı ise % 93, % 82 canlılık sağlamıştır.
3. MATERYAL METOT
3.1. Materyal
Bu çalışmada laktik asit bakterisi materyali olarak Türkiye’nin çeşitli
yörelerinden (Konya, Kahramanmaraş, Ereğli, Mut, Antalya, Ezine, Erzincan, Şanlıurfa) temin edilen 5 adet peynir ve 5 adet yoğurt numunesi kullanılmıştır. Kullanılan yoğurt ve peynir numunelerinin tamamı evlerinde kendi tüketimleri için üretim yapan köylülerden temin edilmiştir. Bu numunelerden izole edilen bakterilerden seçilen 8’i liyofilizasyonla kurutmada kryoprotektan denemesinin materyalini oluşturmuştur.
Kullanılan kriyojenik koruyucu maddeler ise bir kimyasalcıdan analitik saflıkta temin edilmiştir.
3.2. Metot
3.2.1. İzolasyon ve identifikasyon
3.2.1.1. İzolasyonda kullanılan besiyerleri
Yoğurt ve peynir örneklerinde bulunan laktik streptokokların izolasyonu amacıyla M17 Agar, Laktobasillerin izolasyonunda ise MRS Agar kullanılmıştır. Bu besiyerlerinin bileşimi Çizelge 3.1 ve Çizelge 3.2’de verilmiştir.
Çizelge 3.1. MRS Agar bileşimi (de Man, Rogosa ve Sharpe 1960)
Bileşen Miktar (g/L)
Peptone from casein 10.0
Meat extract 10.0
Yeast extract 4.0
D (+) Glucose 20.0
K2HPO4 2.0
Tween 80 1.0
di-Ammonium hydrogen citrate 2.0
Sodium acetate 5.0
MgSO4 0.2
MnSO4 0.04
Agar-agar 14.0
Çizelge 3.2. M17 Agar bileşimi (Terzaghi ve Sandine 1975)
Bileşen Miktar (g/L)
Peptone from soymeal 5.0
Peptone from meat 2.5
Peptone from casein 2.5
Yeast extract 2.5 Meat extract 5.0 Lactose mono-hydrate 5.0 Ascorcic acid 0.5 Sodium β-glycerophosphate 19.0 Magnesium sulfate 0.25 Agar-agar 12.75