T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
SOYA FASÜLYESĠ [GLYCINE MAX (L.) MERR.] GENOTĠPLERĠ ARASINDAKĠ GENETĠK ÇEġĠTLĠLĠĞĠ
DEĞERLENDĠRMEK ĠÇĠN TOHUM DEPO PROTEĠN PROFĠLLERĠ ĠLE AGRONOMĠK ÖZELLĠKLERĠN
KARġILAġTIRMALI ANALĠZĠ
Ömer DURSUN YÜKSEK LĠSANS TEZĠ
Biyoloji Anabilim Dalını
Ağustos–2010 KONYA
TEZ BĠLDĠRĠMĠ
Bu tezdeki bütün bilgilerin etik davranıĢ ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalıĢmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all material and results that are not original to this work.
Ömer DURSUN
iv
ÖZET
YÜKSEK LĠSANS TEZĠ
SOYA FASÜLYESĠ [GLYCINE MAX (L.) MERR.] GENOTĠPLERĠ ARASINDAKĠ GENETĠK ÇEġĠTLĠLĠĞĠ DEĞERLENDĠRMEK ĠÇĠN TOHUM
DEPO PROTEĠN PROFĠLLERĠ ĠLE AGRONOMĠK ÖZELLĠKLERĠN KARġILAġTIRMALI ANALĠZĠ
Ömer DURSUN
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı
DanıĢman: Yrd. Doç. Dr. Emine ARSLAN 2010, 44 Sayfa
Jüri
Yrd. Doç. Dr. Emine ARSLAN Doç. Dr. Yavuz BAĞCI Doç. Dr. Özden ÖZTÜRK
Bu çalıĢmada, tohum depo proteinleri ve agronomik özellikler kullanılarak 12 soya fasülyesi (Glycine max L.) genotipi arasında genetik çeĢitliliği incelemek amaçlanmıĢtır. On iki Glycine max genotipinin olgun tohumları toplanmıĢtır. Her bir genotip için toplanan tohumlardan total protein izole edilmiĢtir. Sodyum dodesil sülfat-poliaakrilamid jel elektroforezi dikey bir slab jel üzerinde standart bir metod ile uygulanmıĢtır. Genotipler arasındaki genetik benzerlik Nei‟nin homolojisine dayanarak tahmin edilmiĢtir. 9.159-148.904 kDa arasında değiĢen 71 polipeptid bant kaydedilmiĢtir. Kümeleme analizi Bio1D++ bilgisayar programında UPGMA kullanılarak uygulanmıĢtır. Glycine max‟a ait 12 genotipin benzerlik kat sayısı, protein profillerindeki genetik varyasyonun düĢük çeĢitliliği ile %75-98 arasında değiĢmiĢtir. Agronomik veriler için kullanılan ortalama Euclid uzaklığı 11.42‟den 60.93‟e değiĢmiĢtir. Bu sonuçlar, hem protein profilleri hem de agronomik özelliklere göre elde edilen dendrogramlarda bazı genotiplerin aynı gruplarda yer aldığını göstermiĢtir.
Anahtar Kelimeler: soya fasülyesi, Glycine max, genotiplerin ayırımı, tohum
v
ABSTRACT MS THESIS
COMPARATIVE ANALYSIS OF SEED STORAGE PROTEIN PROFILES AND AGRONOMIC TRAITS TO EVALUATE GENETIC DIVERSITY AMONG THE
SOYBEAN [GLYCINE MAX (L.) MERR.] GENOTYPES
Ömer DURSUN
The Graduate School of Natural and Applied Science of Selçuk University Biology Branch
Advisor: Asist. Prof. Dr. Emine ARSLAN 2010, 44 Pages
Jury
Asist. Prof. Dr. Emine ARSLAN Doç. Dr. Yavuz BAĞCI Doç. Dr. Özden ÖZTÜRK
This research aimed to investigate the genetic diversity among 12 soybeans (Glycine max) genotypes using seed storage proteins and agronomic traits. Mature seeds of 12 Glycine max genotypes were collected. Total protein was isolated from seeds collected for each genotypes. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis was perfomed by a standard method on a vertical slab gel. Genetic similarity among genotypes was estimated basing on Nei‟s homology. Seventy one polypeptide bands in range of 9.159 kDa to 148.904 kDa were recorded. Cluster analysis was performed using UPGMA in Bio1D++ computer program. Similarity coefficients of 12 genotypes belonging to Glycine max ranged 75-98% with low diversity of genetic variation in protein profiles avarage Euclidian distance used for agronomic data ranged from 11.42 to 60.93. Those results showed that some genotypes were revealed in same groups in dendograms obtained according to both protein profiles and agronomic traits.
Keywords: soybean, Glycine max, genotypes identification, seeds storage
vi
ÖNSÖZ
Bu çalıĢmada, Glycine max kültürlerinden elde edilen olgun tohumlar kullanılarak soya fasülyesi genotipleri arası genetik benzerlikleri SDS-PAGE (Sodyum Dodesil Sülfat Poliakrilamit Jel Elektroforezi) yöntemi ile tespit edilen protein profilleri sonucunda bantların varlığı ve yokluğu değerlendirilerek akrabalık derecelerini gösteren dendogram elde edilmiĢtir. Agronomik özelliklerine dayanan ikinci bir dendogram elde edilerek her iki dendogram kıyaslanmıĢtır. Genotipler arası genetik benzerlikleri ile morfolojik özellikleri arasındaki iliĢkinin belirlenmesi ve seleksiyonun baĢarılı bir Ģekilde yapılması amaçlanmıĢtır.
Tez çalıĢmamın her anında üst düzey bilgi ve tecrübesi ile her konuda bana yol gösteren, danıĢmanlığımı yapan Sayın Hocam Yrd. Doç. Dr. Emine ARSLAN‟a yürekten sonsuz teĢekkürlerimi sunarım. Lisans ve yüksek lisans eğitimim esnasında ve çalıĢma materyalinin temininde, bana hep destek olan dayım Sayın Doç. Dr. Seydi Ahmet BAĞCI‟ya, agronomik özelliklerinin kaydında bana yardımcı olan Sayın Öğr. Gör. Ġrfan ÖZER‟e; agronomik özelliklerin değerlendirilmesinde ve istatistik analizinde bana yol gösteren Sayın Yrd. Doç. Dr. Ahmet TAMKOÇ ve Sayın Yrd. Doç. Dr. Seyit Ali KAYIġ‟a, deney aĢamalarında yardımlarını esirgemeyen Elif GÜLBAHÇE‟ye ve laboratuarda birlikte çalıĢtığımız yüksek lisans arkadaĢlarıma ve bu çalıĢmamda bana destek ve katkıda bulunan isimlerini saymadığım herkese Ģükranlarımı sunarım. Gurbette olduğum günler boyunca her türlü derdime koĢan yengem Kadriye BAĞCI‟ya; aramızda mesafeler olsa da maddi-manevi desteklerini esirgemeyen annem Fatma DURSUN, babam Sefer DURSUN ve kardeĢlerime her zaman minnettarım.
Ayrıca 10201014‟nolu proje ile tezimin yürütülmesinde araç-gereç ve maddi olanak sağlayan Selçuk Üniversitesi B.A.P Koordinatörlüğü‟ne teĢekkürlerimi sunarım.
Ömer DURSUN KONYA- 2010
vii ĠÇĠNDEKĠLER ÖZET ... iv ABSTRACT ... v ÖNSÖZ ... vi ĠÇĠNDEKĠLER ... vii
SĠMGELER VE KISALTMALAR ... viii
1. GĠRĠġ ... 1 2. KAYNAK ARAġTIRMASI ... 4 3. MATERYAL VE METOT ... 11 3.1. Materyal ... 11 3.2. Metod ... 12 3.2.1. Protein Ġzolasyonu ... 12 3.2.2. Jelin Hazırlanması ... 13
3.2.3. Ġzole Edilen Proteinlerin Elektroforezi ... 13
3.2.4. Jellerin Değerlendirilmesi ... 13
3.2.5. Agronomik Özelliklerin Belirlenmesi ... 14
3.2.6. Agronomik Özelliklerin Değerlendirilmesi ... 15
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 16
4.1. SDS-PAGE ile elde edilen sonuçlar ... 16
4.2. Agronomik özelliklerin değerlendirilmesi ... 20
4.3. Protein profilleri sonucu elde edilen dendrogram ile agronomik karakterlerin sayısal değerler sonucu elde edilen dendrogramın kombine değerlendirilmesi ... 23
5. SONUÇLAR VE ÖNERĠLER ... 26
KAYNAKLAR ... 27
EKLER ... 34
viii
SĠMGELER VE KISALTMALAR Kısaltmalar
APS Amonyum Persülfat
ark ArkadaĢları
kDa Kilodalton
mA miliamper
μl mikrolitre
RAPD Rastgele çoğaltılmıĢ polimorfik DNA RFLP Restriksiyon parça uzunluk polimorfizmi
SDS-PAGE Sodyum Dodesil Sülfat Poliakrilamid Jel Elektroforezis
SOD süper oksit dismutaz
TCA Triklorasetik Asit
TEMED N,N,N,N – Tetrametiletilendiamin
Tris Tris (hidroksimetilaminometan )
1. GĠRĠġ
Dünya nüfusunun hızlı artıĢı, çevresel ve coğrafi nedenlerden dolayı ekim alanlarının azalması, bilinçsiz tarım, çevre kirliliği ve kaynakların düzensiz kullanımı gibi sebeplerden dolayı dünyanın pek çok bölgesinde açlık, kıtlık ve toplu ölümler gözlenmektedir. Bu problemin çözümü bitkisel ve hayvansal üretimi arttırmak olarak düĢünülmektedir. En önemli protein kaynağı olan hayvansal proteinlerin zor bulunması, pahalı olması, çabuk bozulması ve kolesterol içeriklerinin yüksek olması bitkisel proteinleri ön plana çıkarmaktadır. Bu amaçla kültür bitkilerinin ıslahı üzerinde binlerce yıldan beri çalıĢılmakla birlikte ürünlerdeki nicelik ve nitelik artıĢı, ancak son 50 yılda geliĢtirilen ıslah yöntemleri ve uygun yetiĢtirme tekniklerinin modern teknolojiyle birleĢmesi sonucunda gerçekleĢebilmiĢtir. Islah edilmiĢ kültür bitkilerinin, yabani formlarıyla karĢılaĢtırıldıklarında birçok mantar, bakteri ve virüs hastalıkları ile zararlılara karĢı daha duyarlı olduğu, bu durumun genelde uygulanan ıslah yöntemlerinin eksikliğinden kaynaklandığı görülmüĢtür. Islah programlarında, seleksiyon çalıĢmalarında ürün kalitesi ve miktarı gibi özellikler ön planda tutulduğundan, hastalık ve zararlılara karĢı dayanıklılık her zaman ikinci planda kalmıĢtır (Özcan ve Özgen 1996).
Dünya nüfusunun beslenmesinde en çok kullanılan yaklaĢık 30 bitki grubu içinde en önemlileri tahıllar, endüstri bitkileri, sebzeler, meyve ağaçları ve baklagillerdir. Bununla beraber dünya florasında yaklaĢık 250 bin bitki türünün bulunduğu ancak bunlardan 3 bin türünün besin değerine sahip olduğu bildirilmektedir (Babaoğlu, 1998).
Soya, tohumlarında bulunan % 18-26 yağ ve % 40 oranındaki protein ile önemli bir yağ bitkisidir. Yağında bulunan oleik ve linoleik yağ asitlerinin yüksek, linolenik yağ asidi oranının düĢük olması nedeniyle kaliteli bir yağa sahiptir. Soya, Dünyada 250-300 değiĢik alanda kullanılır. Ülkemizde ise bisküvi, çikolata, hazır çorba gibi birçok gıda maddesinin yapımında kullanılmaktadır. Soya unu, lesitini, küspesi, proteini, yağı ve yan ürünleri çeĢitli (gıda, kağıt, vs gibi) sanayi kollarının hammaddesidir (Kolsarıcı ve ark. 2005). Ayrıca soya proteininde bulunan yüksek orandaki lycine nedeniyle kanatlı hayvan beslenmesinde önemli bir yere sahiptir. Bu özelliklerinden dolayı soya “Asrın harika bitkisi” olarak tanınmaktadır (Arıoğlu ve ark. 2003).
Soya tarımında birim alandan elde edilen verim; kullanılan çeĢide, uygulanan kültürel yöntemlere ve ekolojik Ģartlara göre değiĢmektedir (Uncu ve Arıoğlu, 2005). YetiĢme mevsimi kısa olan bölgelerde soya verimi, iklim faktörlerindeki yetersizlik nedeniyle düĢmektedir. Özellikle bu bölgelerde geçci çeĢitlerin kullanılmasında genotiplerin yüksek potansiyellerini verime çeviremedikleri gözlenmiĢtir. Oysa erken olgunlaĢan genotipler farklı çevre koĢullarına kendilerini daha iyi adapte ederler. Bu çeĢitler, dane verimine etkili olan çiçeklenmeye ve tane olumuna kadar geçen süreye daha uygun zamanda eriĢirler (Önemli ve AtakiĢi, 2001).
Ülkemizde soya üretimi 1940‟lı yıllarda baĢlamıĢ, önceleri Karadeniz Bölgesi‟nde yetiĢtirilmiĢ, ancak zaman içerisinde soyaya verilen birim fiyatın düĢüklüğü nedeniyle bu bölgede yetiĢtirilen diğer ürünlerle rekabet edememiĢ ve üretiminde fazla bir artıĢ sağlanamamıĢtır. Fakat 1980‟li yıllarda, ikinci ürün projesi kapsamında Çukurova bölgesinde yetiĢtirilmeye baĢlanılması ile üretiminde önemli artıĢlar sağlanmıĢ ve 1987 yılında soya üretimi 250 bin tona ulaĢmıĢtır. Son 5 yıl içinde soyanın ekim ve üretim miktarı Dünya verilerinde artıĢ gösterirken, ülkemizde düzensiz bir seyir izlemiĢtir. Örneğin, 2001 yılında Dünya‟daki ekiliĢ alanı yaklaĢık 77 milyon ha. ve üretim 178 milyon ton iken, 2005 yılında ekim alanı 91 milyon ha. ve üretim miktarı da 210 milyon tona yükselmiĢtir. Ülkemizde ise 2001 yılında 17 bin ha olan ekim alanı, 2002 ve 2003 yıllarında artıĢ göstererek 27 bin ha‟ a kadar yükselmiĢ, fakat 2005 yılında10 bin hektara düĢmüĢtür. Bu azalıĢa paralel olarak üretim miktarı da 30 bin tonlara kadar gerilemiĢtir (Anonim, 2006)
Soya bir baklagil bitkisi olduğu için ihtiyacı olan azotu havanın serbest azotunu kullanarak karĢılamaktadır. Havadan sağlanan bu azot kendinden sonra gelen ürünün azot ihtiyacının bir kısmının da karĢılanmasını sağlamaktadır. Bu sayede daha az azotlu gübre kullanılmaktadır. Daha az azot kullanımı ülkenin akarsu kaynaklarında daha az nitrik birikmesini ve dolayısıyla daha az kirlenmesini sağlamaktadır. Bu yüzden iyi bir münavebe (ekim nöbeti) bitkisi olarak kabul edilmektedir. Soyanın beslenme ve yağ eldesi adına önemi, 1940‟lara kadar anlaĢılamamıĢtır. 1940‟lardan sonra soya yetiĢtiriciliği ve genetiği üzerinde Amerika‟da yoğun araĢtırmalar baĢlatılmıĢtır. Öyle ki aĢırı yağlı besinlerle kolesterol yüklemesi yapan Amerikanlılar kurtuluĢu soya fasülyesinde aramıĢlardır. Yapılan araĢtırmalar ile soya fasülyesinin kolesterolü düĢürdüğü, göğüs kanserini önlediği ve kemikleri güçlendirdiği tespit edilmiĢtir (Fehr, 1987).
Dünyada en çok soya fasülyesi yetiĢtiren ülkeler sırası ile ABD, Çin, Rusya, Brezilya, Endonezya, Kore, Japonya ve Kanada‟dır. Bu ülkelere göre Avrupa‟da Romanya, Yugoslavya ve Türkiye önemli ölçüde soya yetiĢtiren ülkelerdendir. Ülkemizde 1950‟li yıllarda baĢlanılan soya üretimi, Karadeniz ve Akdeniz Bölgeleri‟nde yoğunlaĢmıĢtır. Üretimin büyük çoğunluğunu elinde bulunduran Akdeniz Bölgesi‟nde I. ve II. ürün Ģeklinde yetiĢtirilen soya, Karadeniz Bölgesi‟nde ana ürün olarak yetiĢtirilmektedir. Soya üretimi her ne kadar 1982 yılında ülke çapında ikinci ürün projesinin baĢlatılması ile çok hızlı bir artıĢ göstermiĢse de daha sonraki yıllarda bazı ekonomik ve tarımsal nedenlerden dolayı azalmaya baĢlamıĢtır (ĠĢler ve ÇalıĢkan, 1998).
Soya, bitki geliĢimi, verim ve kalite açısından ekolojik koĢullara tepkisi oldukça yüksek olan kültür bitkilerindendir ve özellikle gün uzunluğu, soya çeĢitlerinin adaptasyon alanlarını dar bir kuĢak içerisine sınırlamaktadır. Yapılan çalıĢmalarda soya fasülyesinin farklı olgunlaĢma grubuna giren çeĢitlerin performanslarının bölgelere göre değiĢtiği gibi bir bölgede aynı olgunlaĢma grubu içerisindeki çeĢitlerin göstermiĢ olduğu performansların da farklı olduğu görülmektedir (ĠĢler ve ÇalıĢkan, 1998).
Bu çalıĢmada, çeĢitli soya fasülyesi genotipleri arasındaki seleksiyonun baĢarılı bir Ģekilde yapılabilmesi için akrabalık derecelerinin belirlenmesi amacıyla tohum depo protein profillerinin ve agronomik özelliklerin karĢılaĢtırmalı olarak analizleri yapılmıĢtır.
2. KAYNAK ARAġTIRMASI
750 cins ve 18000‟den fazla türle temsil edilen Leguminosae familyası ekonomik öneme sahip türleri içeren en geniĢ familyalardan biridir (Ildis, 2001). Leguminosae familyasının ülkemizde 69 cinsi vardır ve bunlara ait takson sayısı 1128 olup, endemik takson sayısı 375‟dir. Endemizm oranı %39.1‟dir. Odunsu ya da otsudurlar. Yapraklar, alternate diziliĢli, genellikle stipullu, bipinnate, basit pinnate, digitate, trifoliate ya da basittir. Çiçekler, actinomorf ya da zygomorf simetrilidir, hypogin (ovaryum üst durumlu) ya da bazen perigin (ovaryum alt durumlu)‟dir. Genellikle hermafroditdirler ve rasem halindedirler. Spika ya da umbel ya da tek tek çıkarlar. Sepal (4-)5. Petaller (1-)5, kanat ya da imbricate Ģeklinde birleĢmiĢ bazen serbest ya da nadiren kısmi olarak yakınlaĢmıĢtır. Stamen 4- çok, genellikle 10, hepsi bir tüp Ģeklinde birleĢmiĢ (monodelphous) ya da en üstteki stamen ayrı, diğerleri tüp Ģeklinde birleĢmiĢ (diadelphous) dirler. Bazen de hepsi serbest olabilir. Karpel 1 superior, marginal plasentalanma vardır. Meyva bir legumedir (Hem ventral hem de dorsal olarak birleĢmiĢ uzunca açılan bir meyvedir). Tohumlar bir ya da daha çoktur (Davis 1970).
Papilionidae veya Fabonidae bir subfamilyaya ait dane legümler son yıllarda, beĢ trib içinde gruplanmıĢtır; Phaseoleae, Vicieae, Cicereae, Aeschynomenae ve Gemistae. Dane legümlerin çoğu ilk üç tribe aittir. Son yıllarda; dane legümler, tohumun yüksek protein içermesinden dolayı bitkisel kaynaklı proteinlerin araĢtırılmasında öncül rol oynarlar (Cereletti, 1979). Bu yüzden özellikle üçüncü dünya ülkelerinde insan diyetinde hayvan proteinlerine iyi bir alternatif olduğu düĢünülmektedir (Summerfield ve Roberts, 1985).
Anavatanı Çin olan soya, Uzak Doğu‟nun en eski ürünlerinden birisidir. 4000– 5000 yıl önce ortaya çıktığı düĢünülmektedir. Soyaya ait ilk yazılı kayıt M.Ö. 2838 yılında Çin imparatoru Shen Nong tarafından yazılan Materia Medica adlı kitapta yer almaktadır. Yüzyıllardır Çinliler, Japonlar, Koreliler ve Güneydoğu Asyalılar soya fasülyesini beslenmelerinde en önemli protein ve yağ kaynağı olarak kullanmıĢlardır. Bu sebeple ve soyanın protein içeriğinin yüksek olması nedeniyle, bu küçük ve eski fasülye “sarı mücevher”, “büyük hazine”, “doğanın mucize proteini” gibi isimler almıĢtır. ġimdi ise soya dünyadaki açlık sorununa bir çare ve geleceğin proteini olarak görülmektedir (Liu, 1997).
Kültürü yapılan soya çeĢitleri Soja alt cinsine aittir. Bu alt cinse dahil türler tek yıllıktır ve yabani olarak Orta ve Uzak Dogu Asya‟da yetiĢmektedir. Glycine max (L.) Merr., bu cinse ait iki önemli türden birisidir ve kromozom sayısı 2n=40‟dır. Günümüzde kültürü yapılan çok sayıda soya çeĢidi Glycine max (L.) Merr. türüne aittir (Arıoğlu, 1999; Weiss, 2000).
Kültür bitkisi olan Glycine max (L.) Merr., yıllık olarak yetiĢir. Soya bitkisi çalı Ģeklindedir ve yetiĢme koĢullarına göre seyrek veya yoğun olarak dallanır (Liu, 1997). Soyada bitki boyu, çeĢit, ekim sıklığı, ekim zamanı ve ortam koĢullarına bağlı olarak 30–150 cm arasında değiĢmektedir. Çoğu çeĢitlerde gövde tüylerle kaplıdır. Soya meyveleri ise legümen tipte olup üzeri tüylerle kaplıdır (Arıoğlu, 1999).
Soya kazık köklü bir bitkidir. Toprak ve yetiĢtirme koĢullarına bağlı olarak kökleri, 1.5–2.5 m derinlere kadar inebilmektedir. Soyada kök ve yan kökler üzerinde Rhizobium bredy japonicum bakterileri tarafından oluĢturulan nodüller bulunmaktadır (Gazzoni, 1994; Arıoğlu, 1999; Weiss, 2000).
Tohum sayısı, çeĢit ve yetiĢtirme koĢullarına bağlı olarak değiĢmekle birlikte genellikle 2–3 tohum/meyve‟dir. Tohumlar yuvarlak, oval veya elips seklindedir. Tohumun rengi genellikle sarıdır. Hilum rengi, sarı, siyah, kahverengi veya kızıl olabilmekte ve bu özellik çeĢitleri birbirinden ayırmada önemli bir kriter olarak kabul edilmektedir (Arıoğlu, 1999; Weiss, 2000).
Olgun tohumlar temel olarak tohum kabuğu, embriyo ve bir veya daha fazla besin depo yapıları olmak üzere üç bölümden oluĢur. Fakat soya tohumları, diğer birçok baklagil bitkisinde olduğu gibi, aslında endospermden yoksundur ve bir tohum kabuğu ve geniĢ, geliĢmiĢ bir embriyo içermektedir. Embriyo, besin depo yapıları olarak görev yapan iki tane kotiledon içerir. Soya dünyadaki en önemli kültür bitkilerinden biridir ve dünya piyasasında yağ üretimininin %48‟ini karĢılamaktadır. Ayrıca hem insan tüketimi hem de hayvan yemi için önemli bir protein kaynağıdır. Çin‟de soya temel yağlı tohum olup yaklaĢık 8 milyon hektar alanda yetiĢtirilmektedir. Bitki yetiĢtiricileri uzun süre soyanın kalitesini ve verimliliğini arttırmak için uğraĢmıĢlardır. Bir genetik bağlantı haritasının yapısı, agronomik (tarımla ilgili) özelliklerin geliĢmesi için önemli olan genetik bilgiyi sağlayacaktır (Liu, 1997).
Griffin ve Palmer (1989), kalitatif genetik çalıĢmalar, gen haritalaması ve bitki yetiĢtiriciliğinde yararlı olabileceği kanısıyla, bazı soya fasülyesi (Glycine max L.) ırklarında, “süper oksit dismutaz (SOD)” izoenziminin varyantlarını kodlayan lokuslar arasındaki baglantıları poliakrilamid jel elektroforezi ile incelemiĢlerdir.
Moleküler markerler içeren bazı genetik bağlantı haritaları 1990‟lı yılların baĢlarından beri yapılmaktadır (Keim ve ark., 1990, 1997; Muehlbauer ve ark., 1991; Akkaya ve ark., 1995; Shoemaker ve Specht, 1995; Zhang ve ark., 1997; Liu ve ark., 2000; Wu ve ark., 2001).
Glycine max X Glycine soja‟nın çaprazlanmasından elde edilen bir populasyon kullanılarak, 365 restriksiyon parça uzunluk polimorfizmli (Restriction Fragment length polymorfism: RFLP) 25 bağlantı grubu, 11 rastgele çoğaltılmıĢ polimorfik DNA (Random Amplified Polymorphic DNA: RAPD) primeri, 3 klâsik marker ve 4 izozim bölgesi içeren soyanın moleküler bir genetik bağlantı haritası oluĢturulmuĢtur. AraĢtırmacılar 1995‟te çeĢitli markırları yaygın bağlantı haritasına entegre etmeyi baĢarmıĢlardır (Shoemaker ve Specht, 1995).
Bakoğlu ve Ayçiçek (2005), Elazığ Ģartlarında soya fasülyesinin tarımsal özellikleri ve verimini belirlemek amacıyla ana ürün Ģartlarında yapmıĢ oldukları çalıĢmalarında, bitki boyu 64-118cm, ilk bakla yüksekliği 12-31cm, bitkide bakla sayısı 21-76 adet, metre karede tohum verimi 200-310 g ve 100 tane ağırlığı 6-17 g arasında değiĢmiĢtir. Ayrıca bakla sayısı ile metrekaredeki verim arasında 0.01 seviyesinde olumlu iliĢkiler bulunmuĢtur.
Malik ve arkadaĢları (2009), çalıĢmalarında 92 soya fasülyesi aksesyonunun tohumlarından Sodyum dodesil sülfat-poliaakrilamid jel elektroforez (Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis: SDS-PAGE) yöntemi ile protein profillerinin genetik varyasyonlarını analiz etmiĢlerdir. Elde edilen 26 bant; genetik çeĢitlilik ve analiz için kullanılan farklı protein peptidlerin sayısına göre değerlendirilmiĢtir. Bütün aksesyonlarda 10 büyük bant ortak olmasına rağmen toplam %50 polimorfik bant tespit edilmiĢtir. Ward‟ın yöntemine göre dendogram düzenlenmiĢ ve aksesyonlar baĢlıca 2 gruba ayrılmıĢ 4 kümeden oluĢtukları gözlenirken SDS-PAGE‟in aksesyonların ayrımında faydalı olduğu tespit edilmiĢtir.
Acarer ve arkadaĢları (2003), Bursa koĢullarında bazı soya fasülyesi çeĢitlerinin verim ve verim ögelerinin belirlenmesi amacıyla 8 soya çeĢidi kullanmıĢlar, araĢtırma sonucunda tohum verimi, bitki boyu, ilk bakla yüksekliği, bitkide bakla sayısı, 100 tane ağırlığı karakterlerinde çeĢitler, yıllar ve yıl x çeĢit interaksiyonunun istatistiki açıdan önemli olduğunu bitki boyunun 56.1–89.7cm, ilk bakla yüksekliğinin 11.2–15.6 cm, bakla sayısının 46.6–72.7 adet ve yüz dane ağırlığının 17.4–19.7 g arasında değiĢtiğini belirlemiĢlerdir.
Cardy ve Beversdorf (1984), elli beĢ soya fasülyesi çeĢitinin 11 izoenziminin elektroforetik bant desenlerini çıkararak, ticari soya türlerinin hızlı bir Ģekilde teĢhisini yapabilecek, etkin bir izozimik teĢhis sistemi geliĢtirmiĢlerdir.
SDS-PAGE ve RAPD-PCR analizleri ile altı soya çeĢitinin parmak izleri çıkarılarak genetik iliĢkileri incelenmiĢtir. Altı soya fasülyesi çeĢitinin identifikasyonu için protein profillerindeki polimorfizmin düĢük olması nedeniyle tek baĢına bu metodun yeterli olmadığı ancak RAPD-PCR profilleri ile desteklenerek elde edilen dendogramda genetik akrabalıkların belirlenebileceğini ortaya koymuĢlardır (Barakat, 2004).
Bir araĢtırma grubu, farklı çeĢitteki soya fasülyesindeki tohum depo proteinlerinin SDS-PAGE kullanarak genetik iliĢkilerini çalıĢmıĢlardır. Elde ettikleri soy ağacına göre bu çeĢitlerin tek bir genetik kaynaktan geldiklerini önermiĢlerdir (El-Shemy ve ark., 2000).
Nikolova ve arkadaĢları (2000) çalıĢmalarında; N, K, B ve S mineralleri bakımından yoksun olan topraklarda yetiĢtirilen soya fasülyelerinde ürünlerin azaldığını gözlerlerken, SDS-PAGE ile mineral eksikliklerinin protein ve polipeptid sentezindeki değiĢiklikleri gözlediklerini belirtmiĢlerdir. Protein ve polipeptid spektrumu ile de bu değiĢiklikleri gözledikleri nicel değiĢikliklerle doğrulamıĢlardır.
Soya fasülyesinin total tohum proteinleri ile ilgili genetik akrabalık çalıĢmalarına sınırlı sayıda rastlanmasına rağmen daha çok soya fasülyesinin protein kalitesi üzerine çalıĢmalar mevcuttur. Ancak soya fasülyesi ile aynı familyada olan (Fabaceae) birçok bitkinin genotiplerinin seçilmesinde SDS-PAGE baĢarılı bir Ģekilde kullanılmıĢtır. Suriye‟nin çeĢitli bölgelerinden toplanmıĢ bazı Vicia türlerinin genetik çeĢitliliğini değerlendirmek için 160 aksesyonun ( 30 Vicia faba, 15 V. narbonensis, 82 V. sativa, 25 V. ervilia 8 aksesyonda diğer Vicia türleri) tohum proteinleri SDS-PAGE tarafından analiz edilmiĢtir. Elde edilen dendrograma göre, V. faba ve V. villosa tüm türler arasındaki en uzak iki tür olmuĢtur (Mirali ve ark., 2007).
Yapılan bir çalıĢmada Lima fasülyesinin (Phaseolus lunatus) yabani türleri arasındaki filogenetik iliĢki tohum depo proteinleri kullanarak incelenmiĢ ve tohum depo protein elektroforezi sonucunda farklı protein bant örnekleri yabani türleri karakterize etmiĢtir (Maquet ve ark., 1999).
Baklagillerden 4 cins ve 11 türe ait 47 aksesyonunun tohum total proteinlerinin SDS–PAGE‟de yürütülmesi ile ayrımı gerçekleĢtirilmiĢtir. Onbir türün hepsinin protein bant örnekleri açıkça ayırt edilebilmiĢ ve ağırlıksız çift grup yöntemi ve aritmetik
ortalamalar (Unweighted Pair Group Method with Arithmetic Mean: UPGMA) kümeleme analizi kullanılarak filogenetik iliĢkiler tespit edilmiĢtir. Morfolojik karakterlere dayalı türlerin sınıflandırılması ile uyumlu olduğu belirlenmiĢtir. Ġlginç olarak morfolojik olarak Phaseolus cinsine yakın olan Glycine max (soya fasülyesi) morfolojik sınıflandırmasıyla tutarsızlık göstererek Cicer cinsine ait aksesyonlarla birlikte kümelenmiĢtir (Valizadeh, 2001).
Yine soya fasülyesi ile aynı familyadan olan 5 ülkeden elde edilen altmıĢ yedi bezelye genotipi; genetik sapma tespiti için tohum protein profilleri SDS-PAGE kullanılarak araĢtırılmıĢtır. Yirmi beĢ protein bant örneği gözlenmiĢ ve bunların arasından 20‟si polimorfik olarak belirlenmiĢtir. Protein profilleri ile agronomik özelliklere iliĢkin veriler birlikte değerlendirilmiĢ ve polimorfik bantların kuantatif özelliklerinin etkilediği belirtilmiĢtir (Ghafoor ve Arshad, 2008).
Nohut‟un (Cicer arietinum L.) 118 aksesyonunda toplam tohum protein, SDS-PAGE ile değerlendirilmiĢ ve genetik çeĢitliliği ve coğrafî iliĢkilerinin belirlenmesi sağlanmıĢtır. Aksesyonlar arasındaki tohum protein profillerindeki varyasyonların major bantlarda değil de küçük bantlarda olduğu görülmüĢtür (Nisar ve ark., 2007).
Farklı bölgelerden toplanan on sekiz mürdümük (Lathyrus sativus L.) aksesyonların tohum protein içerikleri ile tohum ağırlığı varyasyonları değerlendirilmiĢtir. ÇalıĢılan bölgelerdeki örneklerin tohum protein ağırlıkları ile genetik bağımsızlıkları arasında bir korelasyon belirlenmiĢtir. Protein analiz verilerine dayanarak çok değiĢkenli analizde (küme ve faktör analizi) farklı coğrafi bölgelerde ve aynı bölgenin aksesyonları arasında yüksek genetik çeĢitliliği ortaya çıkarmıĢtır (Sammour ve ark., 2007).
Hücrede pek çok iĢlevin yürütüldüğü en önemli makromolekül gruplarından biri olan proteinler büyük çeĢitliliğe ve farklı iĢlevlere karĢın temelde oldukça benzer yapılara sahiptirler. Proteinlerin üstlendikleri önemli görevler nedeniyle yapılarının aydınlatılması, sentezlerinin, iĢlevlerinin, regülasyonlarının ortaya konulması çok büyük önem taĢır (Temizkan ve Arda, 2004).
Bitki sistematikçileri tür ayırımında sadece morfolojik karakterleri kullandıkları için çeĢitli zorluklarla karĢılaĢabilmekte ve morfolojik olarak birbirine benzer iki türü ayırmada tereddüte düĢmektedirler. Kromozom sayıları da eĢitse bu ayırım oldukça zorlaĢmaktadır. Bu nedenle sistematik açıdan problemli taksonomik grupların (cins, tür, varyete vs.) ayırımında moleküler genetik çalıĢmalar kullanılmaya baĢlanmıĢtır. Problemli taksonomik grupların ayırımında kullanılabilen tekniklerden bazıları, Doğal
(native) Jel Elektroforezi, Doğal Olmayan (Denatüre) Jel Elektroforezi: SDS-PAGE, DüĢük Molekül Ağırlıklı Polipeptidlerin Elektroforezi, Ġzoelektrik Odaklama ve Ġki Boyutlu Elektroforez (2-D)‟dir. Yüklü moleküllerin elektriksel alanda ayrılmaları temeline dayanan elektroforez tekniği, proteinlerin analizinde ve ayrılmasında da geniĢ çapta kullanılır. Temelde protein tanımlama (molekül ağırlığını, oligomerik mi, monomerik mi olduğunu, miktarını, saflığını belirlemek vb.) ve saflaĢtırma amacıyla kullanılan bu yöntem doğal veya rekombinant bir proteinin sentezlenip sentezlenmediği; sentezleniyorsa iĢlevsel olup olmadığı hakkında da bilgi verir (Temizkan ve Arda, 2004).
Proteinler, içerdikleri aminoasitlere bağlı olarak, negatif veya pozitif net yüke sahiptirler. Eğer protein karıĢımı içeren bir çözeltiye elektrik akımı uygulanırsa, karıĢımdaki protein molekülleri, elektrik alanda, farklı hızla hareket ederek, moleküller birbirinden ayrılırlar (Hames ve Rickwood, 1990).
Elektrik alanda moleküllerin hareket hızları, molekülün; büyüklüğüne, biçimine, yüküne ve elektrik akımının Ģiddetine bağlı olduğu gibi moleküllerin hareket ettiği ortamın viskositesine, sıcaklığına ve iyon Ģiddetine de bağlı olarak değiĢir (Richards, 1998).
Eletroforezde ayrıĢtırılacak olan karıĢımdaki moleküllerin (numune), üzerinde hareket ettiği ortama destek (support) matriksi denir. Kağıt, selüloz asetat, niĢasta, agaroz ve poliakrilamid jel elektroforez ile ayrıĢtırmada destek matriksi olarak kullanılmaktadır. Poliakrilamid jel, proteinlerin ayrıĢtırılmasında en yaygın tercih edilen destek matriksidir (Alberts ve ark., 1989).
Poliakrilamid jelin tercih nedenleri; Birçok amaç için ayırma gücünün yeterli olması, kullanımının kolay ve zaman kazandırıcı olması, karĢılaĢtırma gerektiren çalıĢmalarda, birçok örneğin birlikte yan yana ayrıĢtırılabilmesi, poliakrilamid jel elektroforezi ile ayrıĢtırılmıĢ örneklere, gümüĢ (silver) gibi hassas boyama metotlarının geliĢtirilmesi, poliakrilamid jel elektroforezinde analizi yapılmıĢ örneğin nitroselüloz membrana transferini sağlayan (blot) metodun geliĢtirilmesidir (Brett ve ark., 1989).
Proteinler denatüre (SDS içeren) bir jelde molekül büyüklüğüne; doğal (native) bir jelde ise molekül biçimi, büyüklüğü ve yüküne göre ayrılırlar. Denatüre ortamda gerçekleĢtirilen elektroforez (SDS-PAGE) molekül ağırlığı belirlemelerinde kullanılır. Molekül ağırlığı bilenmeyen protein örneği, molekül ağırlığı bilinen standart proteinlerle birlikte, aynı jel üzerinde yan yana ceplere uygulanır ve ayrı hatlarda elektroforetik olarak ayrılır. Boyama sonrasında jelde gözlenen bantların
karĢılaĢtırılması ile proteinin molekül ağırlığı hakkında bir fikir edinilebilir. Ayrıca bu tip bir ayrıĢımın sonuçlarını matematiksel olarak değerlendirerek molekül ağırlığını belirlemek de olasıdır (Temizkan ve Arda, 2004).
Yapılan bir çalıĢmada Fabaceae türünün tohum depo proteinlerinin SDS-PAGE‟i üreme sistemini belirlemede kullanmıĢlardır. Protein PAGE örneklerinden, belirlenmiĢ olan polinasyon Ģekli hesaplanarak otogam ve allogam türler arasında protein değiĢikliği dikkatle gözlenmiĢtir. Sonuçlar, legumesin polinasyon mekaniği tipi hakkında elde mevcut bilgiler ile karĢılaĢtırılmıĢtır (Eggi ve Potokina, 1998).
Soya tohumlarının farklı kültürlerinde depo proteinlerinin genetik akrabalığını göstermek amacıyla SDS-PAGE yöntemini kullanmıĢlardır. SDS-PAGE numuneleri kullanılarak farklı kültürlerden örneklerde depo proteinlerinin karĢılaĢtırılmasından kazanılan bilgiler ile soy ağacı düzenlenmiĢtir (El-Shemy ve ark., 2000).
3. MATERYAL VE METOT
3.1. Materyal
ÇalıĢma materyalini Konya Sarayönü MYO araĢtırma arazisine ıslah amaçlı ekilen soya (Glycine max L.) genotipleri ve bu genotiplere ait olgun tohumlar oluĢturmuĢtur. AraĢtırmada kullanılan genotipler ve özelliklerine ait bilgiler Çizelge 3.1‟de verilmiĢtir.
Çizelge 3.1. Glycine max genotipleri hakkında bilgi Glycine max
genotipleri
MenĢei ÇeĢit ya da ÇeĢit adayının GeliĢtirildiği Kurum veya KuruluĢlar
Tescilli çeĢit YetiĢme Grubu
A3935 Amerika Asgrow Firması ÇeĢit 3.9
NE3399 Amerika Nebraska Eyaleti ÇeĢit 3.9
DEFĠENCE Amerika Amerika Tarım Bakanlığı ÇeĢit 4 ARISOY Türkiye Adana; Çukurova Üniversitesi Ziraat
Fakültesi
ÇeĢit 4
ATAKĠġĠ Türkiye Adana; Çukurova Üniversitesi Ziraat Fakültesi
ÇeĢit 3.7
ATAEM 7 Türkiye Antalya; Batı Akdeniz Tarımsal AraĢtırma Enstitüsü Müdürlüğü (BATEM)
ÇeĢit 4.1
NOVA Türkiye Antalya; May Tohumculuk ÇeĢit 3.8
BDS 27 Türkiye Konya; Bahri DağdaĢ Uluslararası Tarımsal AraĢtırma Enstitüsü Müdürlüğü (BDUTEM)
ÇeĢit adayı 3.9
BDS 25 Türkiye (BDUTEM) * ÇeĢit adayı 3.8
BDS 21 Türkiye (BDUTEM) * ÇeĢit adayı 3.8
BDS 11 Türkiye (BDUTEM) * ÇeĢit adayı 3.9
BDS 07 Türkiye (BDUTEM) * ÇeĢit adayı 4
3.2. Metod
3.2.1. Protein Ġzolasyonu
Protein profili analizleri, Saraswati ve Matoh (1993)‟a göre yapılmıĢtır. Her bir genotip için ayrı ayrı 40 mg tohum tartılmıĢtır. Havanda dövülerek toz haline getirilmiĢ ve 1,5 ml‟lik ependorf tüplerine konulmuĢtur. Üzerine ekstraksiyon tamponu (Çizelge 3.2) ilave edilmiĢtir. Tüpler 3 dk. benmaride kaynatılmıĢ ve 10.000 rpm‟de 10 dk. santrifüjlenmiĢtir. Süpernatant yükleme yapılıncaya kadar -20°C de saklanmıĢtır (Saraswati ve Matoh, 1993).
Çizelge 3.2. SDS-PAGE‟de kullanılan tampon ve çözeltiler
Tampon ve Çözeltinin Ġsmi Ġçeriği Miktarı Ekstraksiyon Tamponu 0.5 M Tris HCI (pH 6.8) %10 SDS 2-Merkaptoetanol Üre Saf su 125 μl 200 μl 50 μl 480 mg 275 μl
Boyama Çözeltisi I %100 TCA Metanol Saf su
25 ml 82.5 ml 142.5 ml
Boyama Çözeltisi II %2‟lik Comassie Brillant Blue G–250 10 N KOH %100 TCA 150 ml 12.5 ml 50 ml
%5‟lik Poliakrilamid Jel (Ön ayırma jeli) %30 akrilamid–Bisakrilamid 1 M Tris – HCI Saf su %10 SDS %10APS TEMED 1.7 ml 1.25 ml 6.8 ml 0.1 ml 0.1 ml 0.01 ml %13‟lük Poliakrilamid Jel (Ayırma jeli) %30 Akrilamid–Bisakrilamid 1.5M Tris - HCI Saf su %20 SDS %10 APS TEMED 21,6 ml. 12,5 ml 15,4 ml 0.25 ml 0.25 ml 0.025 ml 3.2.2. Jelin Hazırlanması
%13‟lük ayırma jeli Çizelge 3,2‟deki gibi hazırlanmıĢtır. Üzerine % 5 lik ön ayırma jeli hazırlanıp (Çizelge 3.2) dökülmüĢtür ve tarak yerleĢtirilmiĢtir. Jel donduktan sonra tarak çıkarılmıĢtır. Her bir kuyucuğa 15 μl örnek yüklenmiĢtir (Laemli, 1970).
3.2.3. Ġzole Edilen Proteinlerin Elektroforezi
Jele yüklenen örnekler, ön ayırma jelinde 16 mA, ayırma jelinde 30 mA de izleme boyası jelin sonuna gelene kadar yürütülmüĢtür (Laemli, 1970). Jel, boyama çözeltisi I‟ e (Çizelge 3.2) alınarak bir gece boyunca bekletilmiĢtir. Daha sonra boyama çözeltisi II‟ ye (Çizelge 3.2) alınmıĢtır (Demiralp ve ark., 2000). Saf su ile birkaç kez yıkandıktan sonra fotoğrafları çekilmiĢtir.
3.2.4. Jellerin Değerlendirilmesi
Glycine max genotiplerinin SDS-PAGE sonucu elde edilen jellerin görüntüleri Vilber Lourman görüntüleme cihazında alınmıĢ ve bilgisayara aktarılmıĢtır. Protein profil bantlarının varlığına (1) ve yokluğuna (0) göre genetik benzerlikleri Bio1D++
bilgisayar programı ile UPGMA kümeleme analizi kullanılarak Nei‟nin genetik benzerliğine göre hesaplanarak dendrogramı elde edilmiĢtir.
3.2.5. Agronomik Özelliklerin Belirlenmesi
Glycine max genotiplerine ait tohumlar Konya Sarayönü MYO araĢtırma arazisine 2009 yılı Mayıs ayının ilk haftasında “Tesadüf parselleri deneme deseni‟‟ne göre 3 tekerrürlü olarak ekilmiĢ, hasat öncesi verim üzerine etkili olan 9 morfolojik karakterin; bitki boyu (cm), bitki baĢına dal sayısı (adet), ilk dal yüksekliği (cm), ilk bakla yüksekliği (cm), bitki baĢına bakla sayısı (adet), bakla boyu (cm), baklada tohum sayısı (adet), yüz tohum ağırlığı (g), tohum verimi (kg/da) ölçümleri alınmıĢtır.
Bu çalıĢmada yapılan tüm ölçümler; Tarım ve KöyiĢleri Bakanlığı, Koruma ve Kontrol Genel Müdürlüğü, Tohumluk Tescil ve Sertifikasyon Merkezi Müdürlüğü‟nün 2002 yılında yayınlamıĢ olduğu “Tarımsal Değerleri Ölçme Denemeleri Teknik Talimatı–SOYA” kitapçığındaki talimatlara göre her parselden kenar tesirler çıkarıldıktan sonra geriye kalan alandan tesadüfen seçilen on bitkide belirlenmiĢ ve ortalamaları alınmıĢtır.
Bitki boyu; ana sapın kök tacından son olgun baklaya kadar olan uzunluk cetvel ile ölçülmüĢ cm cinsinden kaydedilmiĢtir. Bitki boyunun yüksek olması makineli hasat için önemlidir.
Bitki baĢına dal sayısı; çiçeklenen bitkilerde dallar sayılıp bitki baĢına düĢen dal sayısı adet olarak hesaplanmıĢtır.
Ġlk dal yüksekliği; toprak seviyesinden ilk dallanmanın baĢladığı yere kadar olan mesafe cetvel ile ölçülmüĢ ve cm cinsinden kaydedilmiĢtir.
Ġlk bakla yüksekliği; kök boğazı ile ilk baklanın bağlandığı nokta arasındaki uzunluk cetvel ile ölçülmüĢ ve cm cinsinden kaydedilmiĢtir.
Bitki baĢına bakla sayısı; bitki üzerinde bulunan tüm baklalar sayılmıĢtır ve ortalaması alınmıĢtır.
Bakla boyu; bakla sapı ile baklanın uç noktasına kadar olan mesafe cetvel ile ölçülmüĢ ve cm cinsinden kaydedilmiĢtir.
Baklada tohum sayısı; her bir bakla içerisindeki tohum adedidir.
Yüz tohum ağırlığı; her parselden 3 tekrarlamalı olarak alınan 100‟er adet tohum hassas terazide tartılmıĢ ve ortalaması alınarak g cinsinden kaydedilmiĢtir.
Tohum verimi; bitki baĢına tohum verimi hesaplanmıĢ, daha sonra bu oran dekardaki bitki sayısıyla çarpılıp tohum verimi (kg/da) hesaplanmıĢtır.
3.2.6. Agronomik Özelliklerin Değerlendirilmesi
Dokuz morfolojik karakterin 3 tekerrürlü ölçümleri değerlendirilmiĢ; ortalamaları alınarak istatistiksel analizi yapılmıĢtır. Euclidian uzaklığına göre NTSYS-pc version 2.1 istatistik paket programı ile bir dendrogram elde edilmiĢtir (Rohlf, 2000).
4. ARAġTIRMA SONUÇLARI VE TARTIġMA
4.1. SDS-PAGE ile elde edilen sonuçlar
Total protein profilleri, SDS-PAGE yöntemiyle %13.5‟luk (ġekil 4.1) ve % 13‟lük (ġekil 4.2) poliakrilamid jel kullanılarak analiz edilmiĢtir. Elde edilen her iki jel birlikte kombine edilerek değerlendirilmiĢ ve bantların varlığı (1) ve yokluğuna (0) göre skorlanmıĢtır (Çizelge 4.1). Tohum depo protein bant örneklerindeki varyasyon incelendiğinde, soya genotiplerindeki genetik çeĢitliliğin çok düĢük seviyede olduğu görülmüĢtür.
ġekil 4.1. SDS-PAGE yöntemiyle analiz edilen Glycine max genotiplerine ait protein profillerini gösteren %13.5‟luk jel elektroforegramı (M; Marker).
ġekil 4.2. SDS-PAGE yöntemiyle analiz edilen Glycine max genotiplerine ait protein profillerini gösteren %13‟lük jel elektroforegramı (M; Marker)
Bu çalıĢmada SDS-PAGE sonucunda elde edilen her iki elektroforograma göre 9,159 kDa – 148,904 kDa arasında değiĢen farklı büyüklüklerde toplamda 71 polipeptid band elde edilmiĢtir. Genotipler arasındaki protein profilleri temel olarak benzer fakat sadece bazı büyüklüklerde minor farklılıklar (ġekil 4.1‟de oklarla gösterilmiĢtir) dikkat çekmiĢtir. Protein profillerinde monomorfik major bantların çoğunlukta olmasına rağmen genotipler yakın akrabalık dereceleri ile de olsa ayrılmıĢlardır.
Çizelge 4.1. Glycine max genotiplerinin SDS-PAGE metoduyla elde edilen protein profillerinin skorlanması
A3935 BDS25 NE3399 BDS21 DEFĠANCE ARISOY BDS11 ATAKĠġĠ BDS27 NOVA ATAEM7 BDS07
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 0 1 0 1 0 0 0 1 0 1 0 0 1 0 1 0 1 1 1 0 0 1 0 0 1 0 0 0 0 0 0 0 1 0 1 1 1 1 1 1 1 1 1 1 0 1 0 0 1 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 1 0 0 1 0 1 0 0 0 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 1 1 1 1 1 1 0 0 0 0 0 1 1 0 0 0 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 1 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 0 0 1 0 0 0 0 0 0 0 1 0 1 1 0 1 1 1 1 1 1 1 0 1 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 1 1 1 1 0 1 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 0 0 1 0 1 1 1 0 1 1 0 0 1 1 0 1 0 0 0 1 0 0 1 1 0 0 1 0 1 1 1 0 0 0 0 0 0 1 1 0 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 0 1 1 0 0 1 0 1 0 0 0 0 1 1 1 1 0 1 0 1 1 1 1 0 0 0 0 1 1 1 1 0 0 0 0 1 0 1 1 0 0 1 0 1 1 1 0 1 1 0 0 1 1 0 1 0 0 0 1
ġekil 4.3. UPGMA kullanılarak Nei‟nin genetik benzerliğine göre elde edilen dendrogram.
On iki soya genotipinin tohum depo proteinlerinin kümeleme analizi UPGMA kullanılarak SDS-PAGE‟nin sonuçları üzerinde uygulanmıĢtır. Küme analizinin sonucunda Nei‟nin genetik benzerliğine göre bir dendogram elde edilmiĢtir (ġekil 4.3). Bu dendograma göre % 75 oranında birbirine benzeyen iki grup oluĢmuĢtur. Birinci grupta % 83-98 benzerlik oranlarında 10 genotip yer almıĢtır. BDS27 ve NOVA birbirine en yakın genotipler olarak (iki genotipi diğer genotiplerden ayıran ortak protein bandı ġekil 4.4‟de gösterilmiĢtir) belirlenirken ATAEM7 genotipi bunlara %96 benzer bulunmuĢtur. Birinci ana grupta yer alan diğer genotipler % 85-95 arasındaki değiĢen oranlarda birbirlerine protein profilleri bakımından genetik olarak oldukça yakındırlar. Genetik olarak % 25 uzaklıkta ikinci ana grubu BDS25 ve DEFĠANCE genotipleri oluĢturmuĢtur (iki genotipi diğer genotiplerden ayıran ortak protein bandı ġekil 4.4‟de gösterilmiĢtir). Bu iki genotip % 97 oranında benzerdir.
ġekil 4.4. BDS25 ve DEFĠANCE genotiplerini diğer genotiplerden ayıran ortak protein bandının (siyah oklarla) ve BDS27 ile NOVA genotiplerini diğer genotiplerden ayıran ortak protein bandının
(beyaz oklarla) elektroforegram üzerinde gösterilmesi (%13‟lük poliakrilamit jel).
Çukurova Üniversitesi Ziraat Fakültesinden elde edilen ARISOY ve ATAKĠġĠ genotipleri %93 benzerlik oranı ile birinci grubun ilk kümesinde toplanmıĢtır. Bu grubun diğer üyelerinden biri Amerika menĢeili (A3935), diğeri de (BDS07) Bahri DağdaĢ Tarımsal AraĢtırma Enstitüsünden elde edilmiĢ olmasına rağmen %93-86 gibi yüksek benzerlikte bir arada toplanmıĢlardır. Ġkinci grubun ikinci kümesinde ise Antalya örnekleri (ATAEM ve NOVA) birbirlerine oldukça yakın yer alırlarken, Amerika menĢeili (NE3399) bir genotip ve aynı ana babaya sahip olan (BDS21, BDS11) genotiplerinin çok yakın benzerlik oranları ile birlikte yer aldığı görülmüĢtür. Bu dendrogram sonuçlarına göre protein profilleri coğrafik dağılıma göre bir ayırım yapamadığı gibi aynı ana ve babaya ait 4 genotipide bir araya toplayamamıĢtır. Bu sonuç farklı genotipler oluĢturmak adına yapılan ıslah çalıĢmalarında önemli olabilir.
4.2. Agronomik özelliklerin değerlendirilmesi
Glycine max genotiplerinin bitki boyu (cm), bitki baĢına dal sayısı (adet), ilk dal yüksekliği (cm), ilk bakla yüksekliği (cm), bitki baĢına bakla sayısı (adet), bakla boyu (cm), baklada tohum sayısı (adet), yüz tohum ağırlığı (g), tohum verimi (kg/da) gibi verime etkili olan 9 agronomik özellik Çizelge 4.2‟ de gösterilmiĢtir.
Çizelge 4.2. * Glycine max genotiplerinden elde edilen agronomik karakterlerin sayısal verileri ÇeĢit ya da çeĢit adayının Adı Bitki Boyu (cm) Bitki BaĢına Dal sayısı (adet) Ġlk dal yüksekliği (cm) Ġlk bakla yüksekliği (cm) Bitki baĢına bakla sayısı (adet) Bakla boyu (cm) Baklada tohum sayısı (adet) Yüz tohum ağırlığı(g) Tohum verimi (kg/da) DEFĠANCE 52.2 3.6 9.7 15 16.7 4.1 2.7 13.3 107.7 BDS 07 75.8 2.9 12.5 18 29.2 4.4 2.8 15.5 188.8 BDS 27 61.9 3.1 10.1 17.2 18.9 4.3 2.6 15.2 129.4 NOVA 60.6 2.9 9.8 15.9 22.2 4.2 2.7 12.6 126.6 BDS 25 73.5 4 10 22.6 24.8 4.6 2.9 17 209.9 ATAEM 7 80,6 2.4 13.4 20.5 22.4 4.2 2.9 12.9 193.3 BDS 21 66.2 4.6 11.1 19 20 4.3 2.8 15.3 159.9 NE 3399 56.7 4.3 7.8 14.5 27.1 4.3 2.6 13.7 171.1 ARISOY 68.7 3.8 8.2 15.8 30.1 4.3 3 12.6 159.9 BDS 11 72.6 2.8 10.1 17.3 32.9 4.3 2.8 15.4 161 A 3935 60.7 3.4 9 16.1 22.7 4 2.7 12.8 137.6 ATAKĠġĠ 63 3.6 8 13.6 28.2 4.1 2.8 12 152.2
* Bu bilgiler Sayın Öğr. Gör. Ġrfan ÖZER‟in çalıĢmasından temin edilmiĢtir.
Genotiplerin gün uzunluğuna tepkilerinin farklı olduğu, erken olgunlaĢan genotiplere göre geç olgunlaĢan genotiplerin fotoperiyoda daha hassas olduğu ve bakla sayısının gün uzunluğu farklılıklarından etkilendiği belirtilmektedir (Whigham ve Minor, 1978).
Bu araĢtırmada incelediğimiz morfolojik karakterlerin hepsi; [bitki boyu (cm), bitki baĢına dal sayısı (adet), ilk dal yüksekliği (cm), ilk bakla yüksekliği (cm), bitki baĢına bakla sayısı (adet), bakla boyu (cm), baklada tohum sayısı (adet), yüz tohum ağırlığı (g), tohum verimi (kg/da)] çevre koĢullarından etkilenmektedir. Ayrıca gün uzunluğu bakımından yetiĢme gruplarına ayrılmıĢlardır. Genotiplerin olgunlaĢması için en fazla gün uzunluğu isteyenler 1. gruta yer alırken gün uzunluğu isteklerine göre gruplar sıralanmaktadır (Hongmin ve ark., 2008).
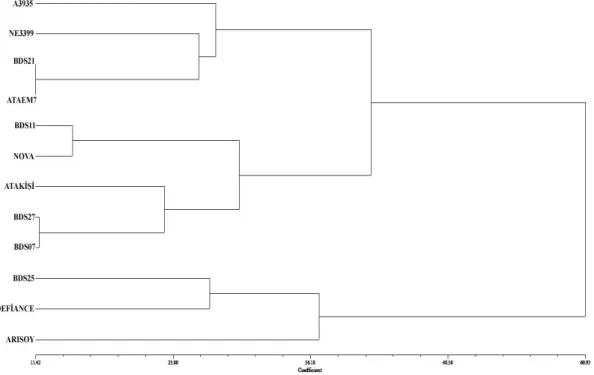
Morfolojik özelliklere dayanarak agronomik karakterin ölçümleri, euclidian uzaklığına göre hesaplanmıĢ ve bir dendrogram elde edilmiĢtir (ġekil 4.5). Bu
dendrograma göre 2 ana grup oluĢmuĢtur. Birinci grup kendi içinde 2 küçük küme meydana getirmiĢtir. A3935, NE3399, BDS21 ve ATAEM7 genotipleri birinci küçük kümeyi oluĢtururken, BDS11, NOVA, ATAKĠġĠ, BDS27 ve BDS07 genotipleri ise ikinci küçük kümede yer almıĢlardır. YaklaĢık % 60 benzer olan ikinci ana grupta BDS25, DEFĠANCE ve ARISOY yer almıĢtır.
ġekil 4.5. Agronomik karakterlere dayanarak Euclidian uzaklığına göre elde edilen dendrogram
Agronomik özelliklere göre elde edilen bu dendrogramda BDS21 ve ATAEM ile BDS27 ve BDS07 genotipleri morfolojik görünüĢlerine göre en yakın çeĢit ve çeĢit adayları olmuĢtur. ARISOY çeĢiti morfolojik olarak aynı yerden elde edilen ATAKĠġĠ‟den ayrılarak ikinci ana grupta yer almıĢtır. Amerika‟dan elde edilen iki genotip (A3935 ve NE3399) birinci ana grubun ikinci kümesinde morfolojik olarak yakın bulunmuĢlardır. Yeni çeĢit elde etmek amacıyla hibritleme sonucu oluĢan kardeĢ genotiplerin hepsi morfolojik olarak benzer bir küme içinde toplanmamıĢlardır. Her biri farklı bir grupta yer almıĢlardır. Dolayısıyla ıslah için farklı bir genotip elde etme amacına ulaĢıldığını göstermektedir.
4.3. Protein profilleri sonucu elde edilen dendrogram ile agronomik karakterlerin sayısal değerler sonucu elde edilen dendrogramın kombine değerlendirilmesi
Protein profillerine göre elde edilen dendogram ile agronomik özelliklere göre elde edilen dendogramı kıyaslarsak, protein profillerine göre soya genotipleri birbirine çok daha yakın akraba çıkmıĢlardır. Her iki dendrogramın, BDS25 ile DEFĠANCE genotiplerinin hem agronomik özelliklerine göre hem de protein profillerine göre diğer genotiplerden ayrılıp farklı bir grup oluĢturması bakımından uyumlu olduğu görülmüĢtür. Fakat diğer genotiplerin meydana getirdiği birinci ana grup içindeki küçük kümeleĢmeler her iki dendograma göre farklılık göstermektedir. Üstelik protein profillerine göre oldukça benzer olarak bir arada toplanan iki Antalya örneği (ATAEM7 ve NOVA ) ile iki Çukurova Üniversitesi Ziraat Fakültesi örneği (ARISOY ve ATAKĠġĠ), morfolojik görünüĢleri bakımından oldukça uzak gruplarda yer almıĢtır.
Morfolojik özellikler bakımından incelendiğinde genotiplerin coğrafîk dağılımı, aynı ana ve babaya sahip olmaları ya da elde edildikleri yer bakımından aynı olmalarına rağmen meydana getirdikleri kümelenmelerin oldukça farklılık göstermesini, fenotipik özelliklerin çevresel faktörlerden farklı derecelerde etkilenmesine bağlayabiliriz. Aynı zamanda genotiplerin sadece 9 agronomik karakterinin kullanımı ile elde edilen dendrogramının, tüm fenotipik özellikleri kapsayan total protein profillerine göre elde edilen dendrogramla uyuĢmamasını, incelenen morfolojik özellikler bakımından eksik olmasına bağlayabiliriz. Total protein profilleri çalıĢıldığında daha fazla morfolojik özellik incelenirse dendrogramlar arası örtüĢme daha fazla olacaktır.
Mujoo ve ark. (2003) soya fasülyesinin protein içeriğinin, soya peynirinin dayanıklılığına, dokusuna ve soya peyniri kalitesine etkisini incelemek amacıyla SDS-PAGE yöntemiyle depo proteinlerini karakterize etmiĢlerdir. AraĢtırmada, depo proteinlerini meydana getiren glisin (11S) ve konglisinin (7S) fraksiyonlarının yoğunluğunu belirleyerek soya peyniri üzerindeki etkileri incelenmiĢ ve SDS-PAGE ile elde edilen total protein profilleri sonucunda 97 kDa ile 6,5 kDa molekül ağırlıklarında soya proteinleri gözlenmiĢtir. ÇalıĢma sonucunda, 148,904 kDa ile 9,159 kDa arasında değiĢen molekül ağırlıklarında bulunan protein profilleri olduğu tespit edilmiĢtir. Barakat (2004)‟ın Glycine max kültürlerinin tohum depo protein büyüklükleri bu çalıĢma ile uyumludur ancak toplam 25 bant gözlemlemesi bakımından çeliĢkilidir. Barakat (2004), yaptığı çalıĢma ile kültürler arasında protein profilleri ile ayrım
yapamamıĢ olmasına rağmen bu protein profillerinin soya fasülyesi için genel bir biyokimyasal parmakizi olarak kullanılabileceğini belirtmiĢtir. Protein polimorfizminin düĢük seviyede olması bir tür yeĢil fasülyede (Hasan, 2001), Poa pratensis aksesyonlarında (Tamkoç ve Arslan 2010) ve birçok soya fasülyesinde de gözlenmiĢtir (Barakat 2004, Mujoo ve ark 2003).
Bonfitto ve ark. (1999), protein polimorfizminin düĢük olmasını tohum proteinlerinin doğal korunmuĢluğuna dayandırılabileceğini savunmuĢtur. Buna rağmen pek çok araĢtırmacı türlerin ve kültürlerin karekterizasyonu ve identifikasyonu için tohum depo protein çeĢitliliğini kullanmıĢlardır (Badr ve ark., 2000; Tamkoç ve Arslan, 2010; Ertuğrul ve ark., 2010; Arslan ve Ertuğrul, 2010).
Bu çalıĢmada, soya fasülyesi genotipleri arasında protein profilleri bakımından genetik çeĢitlilik düĢük olmasına rağmen protein polimorfizmi %52.11 (71 protein bandından 37‟si polimorfik bant) olarak tespit edilmiĢtir. Benzer Ģekilde Malik ve ark. (2009)‟da Glycine max hatlarındaki genetik varyasyonları belirlemek amacıyla inceledikleri tohum protein profillerinin % 50‟sinin polimorfik olduğunu bildirmiĢlerdir. Bunun aksine Barakat (2004) toplam 25 protein bandından 25‟nin de monomorfik olduğunu bulmuĢ SDS-PAGE ile soya fasülyesi kültürlerini ayıramamıĢtır. Aynı Ģekilde Alipoor ve ark. (2002), tohum proteinlerinin elektroforetik örneklerindeki genetik varyasyonu incelemiĢlerdir. Jel üzerinde molekül ağırlıklarına göre yürümüĢ olan protein profiline dayanarak toplam 30 protein bandından sadece 5‟inin aksesyonlar arasında çeĢitlendiği gözlemlenmiĢtir (Polimorfizm oranı %16,66). Ghafoor ve ark. (2003)‟da yaptıkları çalıĢmalarında Alipoor ve ark. (2002)‟larının sonuçlarına benzer sonuçları rapor etmiĢlerdir ve SDS-PAGE‟in farklılıklara dayanarak karekterizasyon ve değerlendirme sonucunda benzer bant profilleri sergilediği için tür içi varyasyona dayanan çeĢitli genotiplerin identifikasyonu için kullanılamayacağını belirtmiĢlerdir. Bu tekniği Vigna ssp.‟deki tür içi varyasyonlarından ziyade türler arası çalıĢmalarda kullanılabileceğini önermiĢlerdir (Ghafoor ve ark., 2002). Bu araĢtırıcıların aksine, bu çalıĢmada genetik çeĢitliliğin düĢük olmasına ve benzer bant profillerinin elde edilmesine rağmen soya aksesyonları arasında polimorfizm oranı %52.11 bulunmuĢtur ve %75–98 arasında değiĢen benzerlik oranları ile gruplaĢmalar meydana gelmiĢtir. Dolayısıyla genotiplerin ayırımı baĢarılmıĢtır. Benzer Ģekilde Ghafoor ve ark. (2002, 2003)‟nın bulgularının aksine birçok araĢtırmacı SDS-PAGE‟in hem tür içinde hem de türler arasında ayırt edebildiğini ve sınıflandırabildiğini savunmuĢlardır (Tamkoç ve Arslan, 2010; Arslan ve ark., 2010; Ertuğrul ve ark., 2010; Uysal ve ark., 2010).
SDS-PAGE ile yer fıstığındaki düĢük genetik çeĢitliği rapor eden Javaid ve ark. (2004) 2D elektroforezi önermiĢlerdir. Benzer bant örnekli genotiplerin detaylı agronomik ve 2D‟yi içeren biyokimyasal analizler ve DNA markırları ile çalıĢılabileceği önerilmiĢtir. (Celis ve Bravo, 1984; Beckstrom-Sternberg, 1989)
5. SONUÇLAR VE ÖNERĠLER
Günümüzde hem morfolojik hem de moleküler yaklaĢımlar türler arasında akrabalık iliĢkilerini çözmek amacıyla kullanılmaktadır. Ancak morfolojik yöntemlerden çok moleküler yöntemler tercih edilmektedir. Moleküler yöntemlerle elde edeceğimiz sonuçların daha kısa zamanda ve daha kesin veriler sağlaması bu tercihin önemini sağlayan etmenlerdir. Baklagiller familyasından olan soya fasülyesi dünyada en önemli endüstri bitkilerindendir. Bu önemi neticesinde birçok araĢtırıcı hem morfolojik hem de moleküler yöntemler kullanarak bu bitki hakkında araĢtırmalar yapmaktadırlar. Endüstri bitkisi olan soya fasülyesinin moleküler yöntemler kullanarak genotiplendirilmesi ve ülkemizde yakın akrabalıklarının tespit edilmesi ülkemiz ekonomisi için büyük önem taĢımaktadır.
Önemli bir protein ve yağ bitkisi olan soya fasülyesi, insan ve hayvan beslenmesinde önemli bir yere sahiptir. Ülkemizde çok yaygın olmayan soya fasülyesi tarımının en önemli problemlerinden birisi yeterli sayıda çeĢitin bulunmamasıdır. Gün uzunluğuna hassasiyeti ile bilinen soya fasülyesinin çevre ve toprak seçiciliği birçok bitkiye göre daha fazladır. Bu özellikleri soya fasülyesi ıslahını daha da önemli kılmaktadır.
Elde edilen sonuçlar, daha zahmetli, masraflı ve zaman alıcı morfolojik görünümlere dayanan klâsik ıslah çalıĢmalarından ziyade daha kolay, ucuz ve güvenilir tohum protein profillerindeki farklılıkların genotiplerin ayırımında kullanılabileceğini göstermiĢtir. Bu avantajlı tekniğin bitki ıslahçılarının istedikleri genotipleri daha kolay ve güvenilir yoldan seçmelerine olanak sağlayacağı kanısındayız. DNA‟nın ekspresyonu sonucu oluĢan proteinler, DNA‟da meydana gelen mutasyonları yansıtacağı için güvenilir sonuçlar elde edilebilir, fakat tohum depo proteinlerini kodlayan genler evrimsel süreçten çok az etkilenen oldukça korunmuĢ genler olduğu için tür içi polimorfizmleri düĢük olmaktadır. Dolayısıyla protein profillerinin de polimorfizmi düĢük olacaktır.
Bu çalıĢma materyalini oluĢturan Glycine max genotiplerinin bu kadar benzer protein bant profillerine sahip olmalarını ve bu kadar çok yakın akraba olmalarını; az sayıdaki soya genotipinin hibritlenmesi sonucu soya çeĢitlerinin geliĢtirilmesine bağlayabiliriz. Bu nedenle DNA markerları kullanılarak oldukça polimorfik sonuçlar elde edilebilir bu da genotiplerin ayırımını tartıĢmasız daha iyi sağlayacaktır.
KAYNAKLAR
Acarer, R., Akkaya, Ġ., Yılmaz, Ö. ve Turan Z.M., 2003, Bursa KoĢullarında Bazı Soya Fasülyesi (Glycine Max (L.) Merill) ÇeĢitlerinin Verim ve Verim Ögelerinin Belirlenmesi Üzerine Bir AraĢtırma. Türkiye V. Tarla Bitkileri Kongresi 13–17 Ekim Bursa, 271-275.
Akkaya, M.S., Shoemaker, C.R., Specht, J.E., Bhagwat, A.A. and Cregan, P.B., 1995, Integration of simple sequence repeat DNA markers into a soybean linkage map, Crop Science 35, 1439-1445.
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. and Watson, J.D., 1989, Molecular Biology of The Cell. 2 nd ed. Garland Publishing Inc. New York.
Alipour H., Rezai A., Meibodi S.A.M. and Taheri M., 2002, Evaluation of genetic variation in soybean lines using seed protein electrophoresis. Journal of Agricultural Science and Technology. 5 (4), 85-96.
Anonim, 2006, www.fao.org.
Arıoğlu, H.H., 1999, Yağ Bitkileri YetiĢtirme ve Islahı, Ç. Ü. Ziraat Fakültesi Genel Yayın No: 220, Ders Kitapları Yayın No: A–70, Adana, 204 s.
Arslan, E. and Ertuğrul, K., 2010, Genetic relationships of the genera Onobrychis, Hedysarum,and Sartoria using seed storage Proteins, Turkish Journal of Biolology, 34, 67-73.
Arslan, E., Ertuğrul, K., Dural, H. and Tamkoç, A., Seed Storage Protein Profile of Some Vetch Species (Vicia L.) Grown in Turkey Using SDS-PAGE, 45th Croatian and 5th International Symposium on Agriculture 15-19 February 2010, Opatija, Croatia.
Babaoğlu, M., 1998, Bitki Doku Kültürleri ve Geleceği, Tarımda Yeni Ufuklar Sempozyumu, Türk Ziraat Yüksek Müh. Bir. Vakfı, 142-148, Ankara.
Badr, A, El–Shazly H.H. and Abou El–Enain, M., 2000, Seed protein diversity and its implications on the relationships in the genus Lathyrus L. (Fabaceae). In: Proc. 1st Conf. Biol. (ICBS), 7–8 May. pp: 333-46. Faculty of Sci., Tanta Univ., Egypt
Bakoğlu, A. ve Ayçiçek, M., 2005, Elazığ ġartlarında Soya Fasülyesinin(Glycine max. L.) Tarımsal Özellikleri ve Tohum Verimi, F.Ü. Fen ve Mühendislik Bilimleri Dergisi, 17(1), 52-58.
Barakat, H., 2004, Genetic Fingerprinting and Relationships of Six Soybeans (Glycine max L.) Cultivars Based on Protein and DNA Polymorphism, International Journal of Agrıculture & Bıology 5, 877-883.
Beckstrom-Sternberg S.M., 1989, Two-dimensional gel electrophoresis as a taxonomic tool: evidence from the Centrospermae, Biochemical Systematics and Ecology, 17, 573-582. Brett, C.J., Curtis, A.S.G., Dow, J.A.T., Edwards, J.G., Lackie, J.M., Lawrence, A.J. and
Moores, G.R., 1989, The Dictionary of Cell Biology, Lackie, J.M. and Dow, J.A.T. (ed.), Academic Pres, London.
Bonfitto R, Galleschi L, Macchia M, Saviozzi F, Navari-Izzo F., 1999, Identification of melon cultivars by gel and capillary electrophoresis, Seed Science and Technology, 27, 779-783.
Cardy, B.J. and Beversdorf, W.P., 1984, Identification of soybean cultivars using isoenzyme electrophoresis, Seed Science and Technology, 12, 943-954.
Celis J.E. and Bravo, R., 1984, Two dimensional gel electrophoresis of proteins; methods and applications, Academic Press, New York.
Cereletti, P., 1979, The Legume Proteins Proc Congre PPI. 30 May – 2 June, Perugia, Italy, 31-57.
Davis, P.H., 1970, Flora of Turkey and the East Aegean Islands Vol.3, Edinburg University Pres.
Demiralp, H., Çelik S. and Köksel H., 2000, Effects of Oxidizing Agents and Defatting on the Electrophoretic Patterns of Flour Proteins During Dough Mixing, Eur. Food Research and Technology, 211, 322-325.
Eggi, E.E., Potokina, E.K., 1998, A Cross Pollinator or A Self Pollinator Electrophoretic Separation of Seed Polypeptides to Determine the Type of Pollination in Leguminous Species, Botanicheskii Zhurnal, 83 (1), 77-83.
El-Shemy, HA., Yamana, H., Saneoka, H. and Fujita, K., 2000, Phylogenetic comparative Analysis of storage structure in legume seeds, American Biotechnology Laboratory vol. 18, no.11.
Ertuğrul, K., Arslan, E. and Tugay, O., 2010, Characterization of Consolida S.F. Gray (Ranunculaceae) taxa in Turkey by seed storage protein electrophoresis, Turkish Journal of Biochemistry, 35 (2), 99-104.
Fehr, W.R., 1987, Breeding Methods for Cultivar Development. In B. E. Caldwell (Ed) Soybeans: Improvement, production, and uses. Agronomy, 16, 249-294.
Gazzoni, D. L., 1994, Tropical Soybean: Improvement and Production, Plant Production and Protection Series, No: 27, 1-17, Rome.
Ghafoor, A., Zahoor, A., Qureshi, A.S. and Bashir, M., 2002, Genetic relationship in Vigna mungo (L.) Hepper and V. radiata (L.) R. Wilczek based on morphological traits and SDS-PAGE, Euphytica.123, 367-2002.
Ghafoor, A., Gulbaaz, F.N., Afzal, M., Ashraf, M. and Arshad, M., 2003, Inter-relationship between SDS-PAGE markers and agronomic traits in Chickpea (Cicer Arietinum), Pakistan Journal of Botany 35 (4), 613-624.
Ghafoor, A. and Arshad, M., 2008, Seed protein profiling of Pisum sativum L., germplasm using Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis(SDS-PAGE) for investigation of biodiversity, Pakistan Journal of Botany, 40 (6), 2315–2321
Griffin, J.D. and Palmer, R.G., 1989, Genetic studies with two superoxide dismutase loci in soybean, Crop Science, 29, 968-971.
Hames, B.D. and Rickwood, D., 1990, Gel Electrophoresis of Proteins, Second Edition, Oxford University Press, Oxford New York Tokyo.
Hasan, H.Z., 2001, Biochemical and molecular genetic characterization of nine mung bean (Vigna radiate L.) cultivars, Bulletin of the Faculty of Science Assiut University, 30 (1), 37-51.
Hongmin, S., Leqiang A. and Xinzhong, T., 2008, Digital Design and Implementation of Soybean Growth Process Based on L-System, Deparment of computing, Engineering College, Northeast Agricultural University, Harbin 50030, China.
Ildis, 2001, Legumes of the World. International Legume Database &Information Service, The University of Reading, UK.
ĠĢler, N. ve ÇalıĢkan, M.E., 1998, GAP Bölgesi Ekolojik KoĢullarında Soyada (Glycine max (L.) Merr.) Verim ve Verime Etkili Bazı Özelliklerin Korelasyonu ve Path Analizi, Turkish Journal of Agriculture and Forestry, 22, 1-5.
Javaid, A., Ghafoor, A. and Anwar, R., 2004, Seed storage protein electrophoresis in groundnut for evaluating genetic diversity. Pakistan Journal of Botany, 30(1): 25-29.
Keim, P., Diers, B.W., Olson, T.C. and Shoemaker, R.C., 1990, RFLP mapping in soybean: association between marker loci and variation in quantitative traits, Genetics, 126, 735-742.
Keim, P., Schupp, J.M., Travis, S.E., Clayton, K., Zhu, T., Shi, L., Ferreira, A., Webb, D.M., 1997, A High- Density Soybean Genetic Map Based on AFLP Markers, Crop Science, 37, 537-543.
Kolsarıcı, Ö., Gür, A., BaĢalma, D., Kaya, M.D. ve ĠĢler, N., 2005, Yağlı Tohumlu Bitkiler Üretimi Raporu.
Laemli, U.K., 1970, Cleavage of structural proteins during the assembly of the head of the bacteriophage T4 Nature 277, 680-684.
Liu, Z., Steward, R., Luo, L. 2000, Drosophila Lis1 is required for neuroblast proliferation, dendritic elaboration and axonal transport, Nature Cell Biology, 2 (11), 776-783.
Malik, Faisal, Anwar, M., Qureshi, A.S., Khan M.R., Ashraf, M., Javed, A., 2009, Evaluation of genetic diversity in soybean (Glycine max) lines using seed protein electrophoresis, Australian Journal of Crop Science 3 (2), 107-112.
Maquet, A., Vekemans, X. and Baudoin, J.P., 1999, Phylogenetic study on wild allies of Lima bean, Phaseolus lunatus (Fabaceae), and implications on its origin, Plant Systematics and Evolution 218 , 43-54.
Markley, S.K., 1950, Soybeans and Soybean Products, Vol 1. New York. USA. P. 15-210. Mirali, N., El-Khouri, S. and Rizq, F., 2007 Genetic diversity and relationships in some Vicia
species as determined by SDS-PAGE of seed proteins, Biologia Plantarum 51 (4), 660-666.
Muehlbauer, B., Schwenk, M., Coram, W.M., Antonin, K.H., Etienne, P., Bieck, P.R. and Douglas, F.L., 1991, Magnesium-L-aspartate-HCl and magnesium-oxide: bioavailability in healthy volunteers, European Journal of Clinical Pharmacology 40, 437-438.
Mujoo, R., Trinh, D.T., Ng Perry, K.W., 2003, Characterization of storage proteins in different soybean varieties and their relationship to tofu yield and texture, Food Chemistry 82, 265-273.
Nikolova, A., Nedeva, D. and Tyankova, L., 2000, Electrophoretic Patterns of Proteins, Isolated From Soybean Seeds Grown Under Conditions of Some Mineral Deficiency and After Different Periods of Storage, Bulgarian Journal of Plant Physiology 26 (1-2), 27-38.
Nisar, M., Ghafoor, A., Khan, MR., Ahmad, H., Qureshi, A.S. and Ali, H., 2007, Genetic Diversity and Geographic Relationship Among Local and Exotic Chickpea Germplasm, Pakistan Journal of Botany, 39 (5), 1575-1581.
Önemli, F. ve AtakiĢi Ġ.K., 2001, Soya (Glycine max (L.) Merr.) ÇeĢit ve Hatları Üzerinde OlgunlaĢma Gün Sayısı Ġle Diğer Verim ve Kalite Unsurları Arasındaki Ġkili ĠliĢkilerin Belirlenmesi Üzerine Bir AraĢtırma. T.Ü. Ziraat Fakültesi. Türkiye 4. Tarla Bitkileri Kongresi, Cilt II Endüstri Bitkileri 355-359.