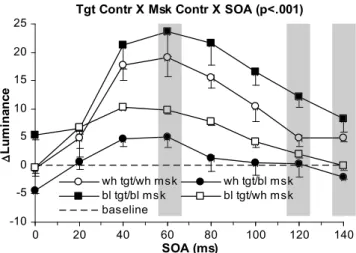
Metacontrast masking and stimulus contrast polarity
Tam metin
Şekil


Benzer Belgeler
The overall aim of the project was to develop a joint networking and advanced research programme on critical issues of planning, management and urban heritage to strengthen
Tüm uygulama gruplarının kontrol grubuna kıyasla cGMP miktarında artış sağladığı belirlenirken 100 µM GA ve 100 µM IAA uygulamaları uygulama süresinin artmasıyla
Visits to the homes of the disabled children and their families enable monitoring the interaction between the family members, 23 evaluation of the family in general,
Nancy Andreasen'in yaratýcý beyin ile ilgili, Schactel'in belleðin sýnýrlarý ile ilgili kitaplarý, psikiyatri topluluðunun yakýndan tanýdýðý, ülkemizin yüz aký, onurlu
Sa¤l›k çal›flanlar› aç›s›ndan; bilgi güvenli¤inin sa¤lan- mas› için kimlik belirleme yöntemleri olarak kullan›c› ad› ve flifre yayg›n
Onun için en kolay yol bu olmuştur; çünkü güçlü bir kız değildir, mücadele edememiştir.. Modern edebiyatın temel izleklerinden biri olan “ölüm”, bu
Bu metaforların analizi sonucunda, öğrencilerin teknoloji kavramına ilişkin algılarının; “yararlı bir şey olarak teknoloji”, “hem yararlı hem de zararlı bir
Psikiyatrik hasta grubu ile psikiyatrik hastalığı olmayan sağlıklı kontrol grubunun yaşadıkları travma şiddetini karşılaştırarak, travmanın benlik saygısı
