T.C.
SELÇUK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
SESELI L. (APIACEAE) CİNSİNİN MOLEKÜLER FİLOGENİSİ
Büşra TOSUN
YÜKSEK LİSANS TEZİ
Biyoloji Anabilim Dalını
Ocak-2015 KONYA Her Hakkı Saklıdır
TEZ KABUL VE ONAYI
Büşra TOSUN tarafından hazırlanan “Seseli L. (Apiaceae) Cinsinin Moleküler Filogenisi” adlı tez çalışması 05/02/2015 tarihinde aşağıdaki jüri tarafından oy birliği / oy çokluğu ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı’nda YÜKSEK LİSANS TEZİ olarak kabul edilmiştir.
Jüri Üyeleri İmza
Başkan
Prof. Dr. Ahmet DURAN ……….. Danışman
Prof. Dr. Ahmet DURAN ………..
Üye
Doç. Dr. Gökhan KARS ………..
Üye
Yrd. Doç. Dr. Ebru DOĞAN GÜNER ………..
Yukarıdaki sonucu onaylarım.
Prof. Dr. Aşır GENÇ FBE Müdürü
TEZ BİLDİRİMİ
Bu tezdeki bütün bilgilerin etik davranış ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalışmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all material and results that are not original to this work.
Büşra TOSUN
iv ÖZET
YÜKSEK LİSANS TEZİ
SESELI L. (APIACEAE) CİNSİNİN MOLEKÜLER FİLOGENİSİ
Büşra TOSUN
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı
Danışman: Prof. Dr. Ahmet DURAN
2015, 86
Jüri
Prof. Dr. Ahmet DURAN Doç. Dr. Gökhan KARS
Yrd. Doç. Dr. Ebru DOĞAN GÜNER
Bu çalışmada Türkiye’nin farklı illerinden toplanan Seseli L. (Apiaceae) cinsine ait 15 taksonun Basit İç Dizi Tekrar dizileri (ISSR) ve İç transkribe olan boşluk (ITS) bölgelerinin moleküler sistematik analizi yapıldı. ISSR ve ITS profillerinin belirlenmesi için her taksonun yaprak ve meyve örneklerinden DNA izolasyonu yapıldı. Taksonlara ait DNA’lar önce ISSR metodu için polimeraz zincir reaksiyonu (PCR) yapıldı ve daha sonra ITS bölgeleri için uygun ITS4 ve ITS5A primerleri kullanılarak PCR ile çoğaltıldı. ISSR yöntemi için elde edilen bantlar skorlandı ve NTSYS-pc paket programı ile filogenetik analiz yapıldı. ITS bölgelerinin baz dizileri elde edilip işlenerek MEGA programı ile filogenetik analizi yapıldı. Değerlendirme sonucu çalışılan türler arasındaki akrabalık ilişkilerini ortaya koyan Neighbor Joining (NJ), Unweighted Pair Group Method Using Arithmetic Average (UPGMA), Maksimum Parsimoni (MP), Maksimum Likelihood (ML) ağaçları elde edildi. ISSR ve ITS çalışmalarından sağlanan verilerin istatistiksel analizi sonucunda Türkiye Seseli taksonları arasındaki filogenetik ilişkiler açıklanmaya çalışıldı. ISSR ve ITS verilerine dayalı oluşturulan farklı dendogramların birbirini desteklediği görüldü. ISSR ve ITS verileri kullanılarak oluşturulan dendogramda Seseli taksonları üç klad altında toplanmıştır. Dış grup taksonları ise ayrı bir kladda yer almıştır.
v ABSTRACT
MS THESIS
Molecular phylogeny of the genus Seseli (Apiaceae)
Büşra TOSUN
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELCUK UNIVERSITY
THE DEGREE OF MASTER OF SCIENCE IN BIOLOGY
Advisor: Prof. Dr. Ahmet DURAN
2015, 86
Jury
Prof. Dr. Ahmet DURAN Assoc. Prof. Dr. Gökhan KARS Asst. Prof. Dr. Ebru DOĞAN GÜNER
In this present study, the regions of Inter Simple Sequence Repeats (ISSR) and nuclear DNA (nrDNA) Internal Transcribed Spacer (ITS) of 15 taxa of Seseli L. (Apiaceae) genus collected from different localities in Turkey, were made molecular systematic analysis. DNA was extracted from leaves and fruits for ISSR and ITS profiles. Firstly, polymerase chain reaction (PCR) were applied all taxa DNA’s for the ISSR methods. For the ITS regions, DNA’s were reproduced by PCR with using ITS4 and ITS5A primers. Bands were scored for the ISSR method, and NTSYS software programme were used for the phylogenetic analysis. Results exhibited that there is a relationships between studied species with respect to obtained Neighbor Joining (NJ), Unweighted Pair Group Method Using Arithmetic Average (UPGMA), Maksimum Parsimoni (MP), Maksimum Likelihood (ML) trees. After the statistical data analysis of ISSR and ITS, phylogenetic relationships were explanied among Turkish Seseli taxa. Dendograms constructed ISSR and ITS parameters supported each other. Seseli taxa cluster three clads in dendogram which was constituted with using ISSR and ITS parameters. Outgroup taxa is placed in the another clad.
vi ÖNSÖZ
Yüksek Lisans çalışmamın başından sonuna kadar her zaman destek veren ve yardımını esirgemeyen saygıdeğer danışmanım Prof. Dr. Ahmet DURAN’a teşekkür ederim.
Moleküler çalışmalarda önerilerinden faydalandığım ve Ziraat Fakültesi Biyoteknoloji Laboratuvarında çalışmama imkân sağlayan Doç. Dr. Erdoğan E. HAKKI’ya, tez çalışmam boyunca bana yardımcı olan, bilgi ve görüşlerinden faydalandığım Yrd. Doç. Dr. Meryem ŞEKER ve Araş. Gör. Özlem ÇETİN, Uzm. Mustafa Çelik ve arkadaşlarım Şerife ATİKER ve Yasemin GÜRBÜZ’e, tez çalışması kapsamında kullanmış olduğum materyalleri sağlamasından dolayı Gazi Üniversitesi öğretim üyesi Yrd. Doç. Dr. Ebru DOĞAN GÜNER’e teşekkür ederim.
14201009 numaralı projeyle, gerekli teçhizatı, kimyasalları ve malzemeleri temin etmemizi sağlayan BAP’a teşekkürlerimi sunarım.
Çalışma konusu olan bitki örneklerini inceleme imkânı sağlayan GAZI, KNYA, HUB herbaryumlarının yetkililerine teşekkür ederim.
Tez çalışmam boyunca benden maddi manevi desteklerini esirgemeyen aileme ayrıca teşekkür ederim.
Büşra TOSUN KONYA-2015
vii İÇİNDEKİLER ÖZET ... iv ABSTRACT ...v ÖNSÖZ ... vi İÇİNDEKİLER ... vii ÇİZELGELERİN LİSTESİ ... ix ŞEKİLLERİN LİSTESİ ... ix SİMGELER VE KISALTMALAR ... xi 1. GİRİŞ ...1 2. KAYNAK ARAŞTIRMASI ...6
2.1. Apiaceae familayasını Morfolojik Özellikleri ...7
2.2. Seseli Cinsi Üzerine Yapılan Moleküler, Morfolojik, Anatomik, Biyokimyasal ve Karyolojik Çalışmalar...8
2.2.1. Seseli Cinsinin Taksonomik Gelişimi ... 10
2.2.2. Selineae Tribusunun Özellikleri ... 15
2.2.3. Selineae Tribusunun Moleküler Çalışmalara Göre Taksonomik Yeri ... 17
3. Moleküler Markerlar ve Önemi ... 25
3.1. Moleküler Markırların Özellikleri ... 26
3.1.1. Moleküler Sistematikte Kullanılan Yöntemlerden Bazıları ... 26
3.2. ISSR (Inter Simple Sequence Repeats) Basit İç Dizi Tekrarları ... 26
3.3. ITS (Internal Transcribed Spacer) ve rDNA ... 27
3.2.1. ITS’nın Genel Özellikleri ... 28
3.2.2. ITS Bölgesinin Filogenetik Çalışmalarda Kullanılışı ... 28
3.2.3. Filogenetik Analiz ve Ağaç Oluşturma ... 29
4. MATERYAL VE YÖNTEM ... 32
4.1. Materyal ... 32
4.1.1. Çalışmada Kullanılan Bitki Örnekleri ... 32
4.1.2. Kullanılan Cam Malzeme Ve Plastik Malzemelerin Sterilizasyonu... 33
4.1.3. Moleküler Çalışmalarda Kullanılan Kimyasal Maddeler... 33
4.1.4. Moleküler Çalışmalarda Kullanılan Tampon ve Çözeltiler ... 33
4.1.5. Genomik DNA İzolasyonunun Yapılması ... 33
4.1.6. Seseli Cinsine Ait Örneklerin ISSR Primerleriyle PCR Amplifikasyonları . 36 4.1.7. Seseli Cinsine Ait Örneklerin ITS Primerleriyle PCR Amplifikasyonları .... 37
4.1.8. Agaroz Jel Elektroforezi ve Görüntüleme ... 37
4.1.9. Dizileme ve Dizi Analizi ... 38
viii
5. ARAŞTIRMA SONUÇLARI VE TARTIŞMA ... 39
5.1. ISSR Yöntemine Göre Elde Edilen Jel Görüntüleri ... 40
5.1.2. ISSR Verilerine Göre Oluşturulan Filogenetik Ağaç ... 42
5.2. ITS Yöntemine Göre Elde Edilen Jel Görüntüsü ... 44
5.2.1. ITS Verilerine Göre Oluşturulan Filogenetik Ağaç ... 44
6. SONUÇLAR VE ÖNERİLER ... 51
KAYNAKLAR ... 54
EKLER ... 62
EK-1. Çalışılan Taksonların ITS Dizileri ... 62
ix ÇİZELGELERİN LİSTESİ
Çizelge 1.1. Apiaceae familyasının dünya’da, Asya kıtası ve Türkiye’de geniş yayılışlı
cinslerinin yaklaşık tür sayıları………....4
Çizelge 1.2. Apiaceae familyası için en fazla tür çeşitliliğine sahip Asya ülkeleri……..5
Çizelge 1.3. Türkiye bölgelerinde Apiaceae familyasına ait cins ve tür sayısı………….5
Çizelge 2.1. Apioideae tribus ve subtribusları……….19
Çizelge 2.2. Selineae tribusu içindeki cinsler………..23
Çizelge 2.3. Selineae tribusu içindeki polifiletik cinsler……….24
Çizelge 4.1. Çalışılan türler ve toplandığı lokaliteler………..32
Çizelge 4.2. ISSR amplifikasyonunda kullanılan primerler………36
Çizelge 4.3. ITS yönteminde kullanılan primerler………..37
Çizelge 4.4. ITS yönteminde kullanılan PCR programı………..37
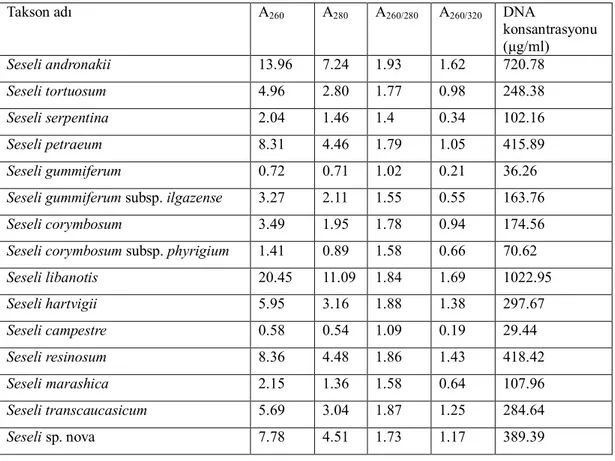
Çizelge 5.1. İncelenen örneklerin DNA yoğunlukları ve spektro değerleri………39
ŞEKİLLERİN LİSTESİ Şekil 2.1. Seseli cinsinin Türkiye’deki yayılışı………...14
Şekil 2.2. Selineae tribusunun filogenetik ağaçta gösterimi………..21
Şekil 3.1. Çekirdek ribozomal DNA’sının tekrarlı üniteleri………...28
Şekil 5.1. Jel görüntülerinde kullanılan markerın DNA baz uzunlukları………40
Şekil 5.2. İncelenen türlerin M13 primeriyle PCR amplifikasyonundan elde edilen sonuçların elektroforez jel görüntüsü .……… 40
Şekil 5.3. İncelenen türlerin M13 primeriyle PCR amplifikasyonundan elde edilen sonuçların elektroforez jel görüntüsü ………41
Şekil 5.4. İncelenen türlerin M13 primeriyle PCR amplifikasyonundan elde edilen sonuçların elektroforez jel görüntüsü……….41
Şekil 5.5. Türkiye Seseli taksonlarının ve dış grup olarak kullanılan taksonların ISSR primerleri ile yapılan PCR amplifikasyonlarının skorlanarak NTSYS programında, UPGMA analizi ile değerlendirilmesi sonucu elde edilen dendogram ………… 42
Şekil 5.6. İncelenen türlerin ITS4-5M primeriyle PCR amplifikasyonundan elde edilen sonuçların elektroforez jel görüntüsü……….44
x
Şekil 5.7. Türkiye Seseli taksonlarının ve dış grup olarak kullanılan taksonların ITS primerleri ile yapılan PCR amplifikasyonlarının MEGA programında, Neighbor Joining analizi ile değerlendirilmesi sonucu elde edilen dendogram …...45 Şekil 5.8. Türkiye Seseli taksonlarının ve dış grup olarak kullanılan taksonların ITS primerleri ile yapılan PCR amplifikasyonlarının MEGA programında, Maksimum parsimoni analizi ile değerlendirilmesi sonucu elde edilen dendogram………….47 Şekil 5.9. Türkiye Seseli taksonlarının ve dış grup olarak kullanılan taksonların ITS primerleri ile yapılan PCR amplifikasyonlarının MEGA programında, Maksimum Likelihood analizi ile değerlendirilmesi sonucu elde edilen dendogram………..48 Şekil 5.10. Türkiye Seseli taksonlarının ve dış grup olarak kullanılan taksonların ITS
primerleri ile yapılan PCR amplifikasyonlarının MEGA programında, UPGMA analizi ile değerlendirilmesi sonucu elde edilen dendogram……….……49 Şekil 5.11. Dünya Seseli taksonlarının ve dış grup olarak kullanılan taksonların ITS primerleri ile yapılan PCR amplifikasyonlarının skorlanarak MEGA programında, neighbor-joining analizi sonucu elde edilen dendogram………...50
xi
SİMGELER VE KISALTMALAR
Simgeler ve Kısaltmalar Açıklamalar
bç baz çifti
CTAB Setil Trimetil Amonyum Bromür
DNA Deoksiribo Nükleik Asit
dH2O Distile Su
dNTP Deoksiribo Nükleosid Trifosfat EDTA Etilen Diamin Tetra Asetik Asit
ETS Dış trankribe boşluklar
GAZI Gazi Üniversitesi Herbaryumu
HCl Hidroklorikasit
HUB Hacettepe Üniversitesi Herbaryumu
ISSR Inter Simple Sequence Repeat
ITS Internal Transcribed Spacer
KNYA Selçuk Üniversitesi Herbaryumu
NCBI National Center For Biotechnology Information
NTSYS-pc Numerical Taxonomy and Multivariate Analysis System MEGA Molecular Evolutionary Genetics Analysis
MgCl2 Magnezyum Klorür
ML Maksimum Likelihood
MP Maksimum Parsimoni
NaCl Sodyum Klorür
NAOH Sodyum Hidroksit
NJ Neighbor Joining
nrDNA Nüklear ribozomal DNA
PCR Polimeraz Zincir Reaksiyonu
RAPD Randomly Amplified Polymorphic DNA
SDS Sodyumdisülfit
subsp. Alttür
Taq Thermus aquaticus
xii
TBE Tris-Borikasit- EDTA
Tm Erime Sıcaklıkları
UPGMA Unweighted Pair Group Method Using Arithmetic Average
1. GİRİŞ
Türkiye coğrafi konumu, jeomorfolojik yapısı, farklı topografik yapılara ve toprak gruplarına sahip oluşu, değişik iklim tiplerini bir arada bulundurması, İran-Turan, Akdeniz ve Avrupa-Sibirya olmak üzere üç farklı bitki coğrafyasının birleştiği yerde bulunması, buzul dönemlerinde canlılar için sığınak olması ve bazı cinslerin gen merkezi olması nedeniyle oldukça zengin bir flora ve çok farklı vejetasyon tiplerine sahiptir (Davis ve Hedge, 1975).
Türkiye’de cins altı düzeyde toplam 11.466 doğal takson bulunmaktadır. Türkiye için endemik olan doğal takson sayısı 3.649 olup endemizm oranı % 31,82’dir (Güner ve ark., 2012).
Ülkemizin zengin bir floristik yapıya sahip olması her zaman botanikçilerin ilgisini çekmiştir. Farklı zamanlarda çok sayıda yabancı botanikçi ülkemiz bitki çeşitliliği üzerine araştırmalar yapmıştır. Bu çalışmalar çok eski tarihlere dayanmasına rağmen bitki toplama amacıyla yapılmış araştırmalar 16 yy. ortalarından itibaren başlamıştır. Fransız doğa bilimci Pierre Belon (1517-1564) ülkemize bitki toplama amaçlı gelen ilk araştırmacıdır ve İstanbul, Ege Adaları, İzmir ve Uludağ civarlarından bitki örnekleri toplamıştır (Sağıroğlu, 2005; Doğan, 2007).
Ülkemiz florası ile ilgili ilk önemli eser İsviçreli botanikçi Pierre Edmond Boissier’in 1865-1888 yılları arasında yayınlanan ve 6 ciltten oluşan Flora Orientalis adlı eseridir (Boissier, 1867-1888). Ülkemiz bitki çeşitliliği ile ilgili en önemli eser ise, Flora Orientalis‘ten tam bir asır sonra yazılan, editörlüğünü Peter Hadland Davis‘in yaptığı dokuz ciltlik Flora of Turkey and the East Aegean Islands adlı eseridir (Davis, 1965-1985). Türkiye florasının ilk dokuz cildinin yayınlanmasından sonra çok sayıda yeni takson ortaya çıkartılmış ve bu taksonların ek cilt olarak yayınlanmasıyla cilt sayısı 10’a çıkartılmıştır (Davis ve ark., 1988). Daha sonra yapılan çalışmalarda Türkiye Florasına ilave edilen yeni kayıt ve taksonlar için Türk botanikçiler tarafından ikinci bir ek cilt yayınlanmıştır (Güner ve ark., 2000). Ülkemiz florası ile ilgili diğer önemli eser ise; 2012 yılında tamamlanan, Türk araştırmacılar tarafından hazırlanmış olan ve Türkiye bitki zenginliğinin tamamını kapsayan Türkiye Bitkileri Listesi (Damarlı
Bitkiler) eseridir (Güner ve ark., 2012).
Ülkemizde yabancı botanikçiler tarafından yapılmış olan floristik çalışmalar kısa süreli ve genellikle ulaşımı kolay olan yerlerde yapılmıştır (Sağıroğlu, 2005). Floranın
tamamlanmasının ardından yapılan floristik çalışmalar ve yeni taksonların tanımlanması, Türkiye florasının tam anlamıyla bitirilemediği göstermiştir. Bu nedenle çok sayıda floristik çalışma yapılmış ve yapılmaya devam etmektedir. Aynı zamanda
Flora of Turkey eserinin yazımı sırasında sınırlı zaman ve materyal ile çalışıldığından
dolayı birçok cinsteki eksiklikler bu eserde belirtilmiş, ancak çözüm getirilememiştir (Davis, 1965-1985). Toplanan çok sayıda örnek teşhis edilirken karşılaşılan sorunlar bazı cinsler hatta familyalardaki problemlere dikkati çekmektedir. Özellikle bu taksonlardaki varyasyon sınırlarının ve yeni olabilecek taksonların tespiti için öncelikle cins düzeyinde revizyon çalışmaları ileri teknikler kullanılarak gerçekleştirilmektedir. Bu amaçla son yıllarda Türk araştırmacılar tarafından özellikle taksonomik açıdan problemli cinsler üzerinde revizyon çalışmalarının yapılmasına ağırlık verilmektedir. (Doğan Güner, 2006; Akçoşkun, 2010).
Apiaceae (Umbelliferae) familyası dünyada yaklaşık olarak 450 cins ve 3700 türe sahiptir ve çiçekli bitkilerin en önemli familyalarından biridir (Pimenov ve Leonov, 1993). Ekonomik açıdan da oldukça önemli bir familyadır. Apiaceae familyasının en iyi bilinen üyeleri arasında havuç (Daucus carota L.), maydanoz (Petroselinum crispum (Mill.) Nyman ex A.W. Hill), dereotu (Anethum graveolens L.), kişniş (Coriandrum
sativum L.), rezene (Foeniculum vulgare Mill.) ve kereviz (Apium graveolens L.)
sayılabilir. Zehirli olanlar ile geniş tıbbi kullanım alanına sahip olan türler Apiaceae familyasının ayırt edici kimyasal yapısını ortaya koymaktadır (Downie ve ark., 1998).
Apiaceae familyası üyeleri morfolojik olarak oldukça sabit karakterlere sahiptir; Theophrastus’un “doğal bitki familyası” olarak adlandırdığı familyalardan birisidir (Berenbaum, 2001). Genel olarak bazı belirgin karakterlere (iki merikarptan oluşan bir şizokarpik meyve vb.) sahip olması nedeniyle Apiaceae “çiçekli bitkilerin teşhisi yapılan ilk familyası” olmuştur (Constance, 1971). Üstelik Robert Morison’un 1672’de
Plantarum Umbelliferarum Distributio Nova eserinin yayınlaması ile birlikte Apiaceae
familyası, monografı yapılan ilk çiçekli bitki grubu olmuştur (Constance 1971, Hedge 1973).
Apiaceae familyasının sistematiği büyük ölçüde olgun meyvenin anatomik ve morfolojik özelliklerine dayanır (Lee ve Downie, 1999). Ayrıca Apiaceae familyası taksonlarının taban yaprakları da bu familyanın sistematiğinde önemli bir yere sahiptir. Bu familyanın birçok cinsinde çiçeklenme dönemi geç olduğundan olgun meyveli örnekler geç dönemdeki arazi çalışmaları sonucu toplanabilir. Araziden toplanan
örnekler teşhis anahtarında belirtilen karakterlere uygun olmadığında yanlış teşhis ihtimali artar. Bu nedenle Apiaceae familyası taksonları Türkiye ve dünyada en fazla probleme sahip bitkilerdendir (Güner, 2006).
20. yy ortalarından itibaren, DNA’nın yapısının açıklanmasından sonra, biyoloji bilimlerindeki gelişmeler, büyük bir hız kazanmıştır. Saiki ve ark., (1988) mevcut yöntemleri geliştirerek, DNA’nın aslına uygun bir şekilde in vitro olarak çoğaltma esasına dayalı bir teknik olan PCR‘ı (Polimeraz Zincir Reaksiyonu) geliştirmeleri ile çığır açmış, moleküler yöntemlerle bitki ve hayvanların daha sağlıklı bir şekilde filogenetik sınıflandırmaları yapılmaya başlanmıştır (Öztürk, 2011).
Günümüzde türlerin tanımlanmasında, kromozom haritalamalarında, gen kaynaklarının belirlenmesinde, evrimsel ilişkilerin ortaya konulmasında ve genetik varyasyonların araştırılmasında moleküler yöntemler kullanılmaktadır. Bu yöntemlerin polimorfizm oranlarının yüksek olması, pleotropik ve epistatik etki göstermeyip oldukça istikrarlı olmaları, çevre faktörlerinden etkilenmemeleri klasik yöntemlere göre büyük avantajlar sağlamaktadır (Çelik, 2013).
DNA baz dizinindeki polimorfizmin ortaya çıkarılması için RFLP, AFLP, SSR, RAPD ve ISSR gibi moleküler markör teknikleri sıkça kullanılmaktadır (Öztürk, 2011). Araştırmacılar, Apiaceae familyasının yüksek düzeydeki akrabalık ilişkilerinin çözülmesi için çalışmalar yapmışlardır. Bu çalışmalar 1990’lı yıllardan itibaren hız kazanmıştır. Apiaceae familyası içerisindeki yüksek düzeydeki filogenetik ilişkilerin çözülmesi, belli tribusların revizyonlarının ve cins komplekslerinin taslaklarının oluşturulması için gereklidir. Bu yüksek düzeydeki filogenetik ilişkilerin çözülmesi için modern sınıflandırma yöntemlerinin kullanılması kaçınılmaz olmuştur. Günümüzde Apiaceae familyası için yeni ve işlevsel bir sınıflandırma sistemi oluşturmak amacıyla farklı moleküler karakterler kullanılmaktadır. Kloroplast genleri (rbcL, matK) ve intron dizileri (rpoC1, rps16), çekirdek ribozom DNA’sı ITS dizileri bu karakterlerden bazılarıdır (Downie ve ark., 2000a).
Apiaceae familyası dünyada yaklaşık 450 cins ve 3700 tür içerir (Pimenov ve Leonov, 1993). Türlerin cinsler arasındaki dağılımı eşit değildir: cinslerin %41’itek tür ile temsil edilirken (monotipik), %26’sı sadece 2–3 türle temsil edilir. Türlerin % 60’ı ise her biri 20’den fazla tür içeren birkaç geniş cinste yer almaktadır. Polifiletik olan bu cinslerden bazılarının tür sayıları şöyledir; Ferula L. 177 ve Ligusticum L. 40-50 (Spalik ve ark., 2004).
Apiaceae familyasının Asya’da en fazla türle temsil edilen cinsleri sırasıyla şunlardır; Ferula 177, Bupleurum L. 155, Pimpinella L. 125, Heracleum L. 109, Seseli L. 101, Angelica L. 87, Bunium L. 43, Prangos Lindl. 43, Ferulago W.D.J.Koch 37,
Hymenidium Lindl. 35, Hydrocotyle L. 35, Chaerophyllum L. 34, Eryngium L. 34, Pternopetalum Franch. 32, Acronema Falc. ex Edgew. 23 ve Semenovia Regel & Herder
22 türe sahiptir (Çizelge 1.1.). Özellikle bazı büyük cinslerin dünya genelindeki tür sayıları birbirine yakındır. Bunu dünyadaki tür sayısı Asya’da yoğunlaşmış olan cinslerde görebiliriz ki bu cinslere örnek olarak Ferula, Seseli, Bupleurum, Pimpinella,
Bunium, Ferulago, Prangos ve Asya’ya özgü olan Hymenidium, Pternopetalum, Acronema ve Semenovia cinsleri verilebilir (Pimenov ve Leonov, 2004; Güner ve ark.,
2000; Özhatay ve Kültür, 2006; Özhatay ve ark., 2008-2009, 2009, 2011).
Çizelge 1.1. Apiaceae familyasının dünya’da, Asya kıtası ve Türkiye’de geniş yayılışlı cinslerinin yaklaşık tür sayıları (Güner ve ark., 2012)
Apiaceae familyası için en fazla tür içeren Asya ülkeleri Çin, Türkiye (Anadolu bölümü), İran, Rusya (Asya bölümü) ve Kazakistan’dır. Familyaya ait en fazla tür sayısı Çin Florası için verilmiştir, 108 cinste 677 tür vardır. Türkiye coğrafik açıdan daha
Cins adı Türkiye Asya Dünya
Ferula Bupleurum Pimpinella Heracleum Seseli Angelica Bunium Prangos Ferulago Hymenidium Chaerophyllum Eryngium Pternopetalum Elaeosticta Acronema Semenovia 22 47 25 17 13 3 12 14 34 - 17 25 - 1 - - 177 155 125 109 101 87 43 42 37 35 34 33 32 26 23 22 180-185 185-195 170-180 120-125 125-140 110-115 45-50 43 47 35 45 250-260 32 26 23 22
küçük olmasına rağmen, tür sayısı bakımından ikinci sırada yer alır ve 109 cinse ait ~455 tür ile temsil edilir (Çizelge 1.2.). Türkiye’de yayılış gösteren türlerin yaklaşık %33 endemiktir. Bu durum Türkiye’nin Asya’da ve büyük olasılıkla da dünyada, Apiaceae familyası için en yüksek tür çeşitliliğine sahip ülke olduğunu gösterir (Pimenov ve Leonov, 2004; Davis ve ark., 1988; Güner ve ark., 2000; Özhatay ve Kültür, 2006; Özhatay ve ark., 2008-2009; Özhatay ve ark., 2009; Özhatay ve ark., 2011).
Çizelge 1.2. Apiaceae familyası için en fazla tür çeşitliliğine sahip Asya ülkeleri
Türkiye’nin yanı sıra bir bütün olarak Güneybatı Asya da Apiaceae tür çeşitliliği bakımından oldukça zengindir. İran, Gürcistan, Suriye, Azerbaycan, Irak, Ermenistan ve Lübnan Apiaceae familyasına ait tür sayısı yüksek ülkelerdir. Buna karşılık Umman, Kuveyt, Birleşik Arap Emirlikleri, Bahreyn ve Katar tür çeşitliliği düşük olan ülkelerdir (Pimenov ve Leonov, 2004).
Türkiye’de yayılış gösteren Apiaceae familyasına ait taksonların coğrafi bölgelere dağılımı düzenli değildir. Beş coğrafik bölge içerisinden Güneybatı ve Doğu Anadolu en yüksek tür çeşitliliğine sahiptir (Çizelge 1.3.). Doğu Anadolu’da 80 cinste 242 tür vardır ve bu türlerden 15’i endemiktir (Pimenov ve Leonov, 2004).
Çizelge 1.3. Türkiye bölgelerinde Apiaceae familyasına ait cins ve tür sayısı Ülke Cins Sayısı Tür Sayısı
Çin Türkiye (Asya böl.) İran Rusya (Asya böl.) Kazakistan 108 109 111 105 78 677 455 350 278 236
Bölge Cins Sayısı Tür Sayısı Kuzey Anadolu Orta Anadolu Batı Anadolu Güneybatı Anadolu Doğu Anadolu 74 80 64 82 80 185 221 132 251 242
Çin ve Türkiye’den sonra en yüksek tür çeşitliliği İran’da görülür. İran’ın doğusunda bulunan Orta Asya ülkeleri de yüksek düzeyde tür çeşitliliğine sahiptir. Orta Asya ve Kazakistan’dan yaklaşık olarak 430 tür bilinmektedir. Tianshan ve Pamiro-Alai dağlarında 19 endemik cins bulunmaktadır. Endemik cinslerin fazlalığı, oldukça kurak düzlüklerle çevrili yüksek dağ sistemlerinin sağlamış olduğu izolasyonun yanı sıra bölgenin sınır yerleşiminin Orta Asya sınırına yakınlığıyla da açıklanabilir. Bu bölgelerde, Apiaceae familyası için bazı alışılmadık karakterler de gözlenebilir. İkincil durumda çalı özelliği gösteren Schrenkia kultiassivii ve bazı Seseli cinsine ait türlerdeki odunlaşmış toprakaltı gövde bu karakterlere örnek olarak verilebilir (Pimenov ve Leonov, 2004).
Bu çalışmanın konusu olan Seseli cinsinde yetersiz toplamalar ve eksik materyalle çalışıldığı için karşılaşılan sorunlar ile içerdiği problemler bu cinsin revizyonunun yapılmasının gerekliliğini ortaya koymuş ve 2006 yılında cinsin revizyonu tamamlanmıştır (Doğan Güner, 2006).
Seseli L. cinsi ile ilgili yaptığımız kapsamlı literatür araştırmalarından elde
edilen bilgiler doğrultusunda bu cinsin taksonomik problemlerinin yapılan revizyon çalışmasıyla büyük ölçüde giderildiği ancak modern taksonomiye göre moleküler yöntemlerle de desteklenmesinin önceki çalışmaları güçlendireceği öngörülmüştür.
2. KAYNAK ARAŞTIRMASI
Apiaceae Lindl. (Umbelliferae Juss.) familyası dünya genelinde çöller, bataklıklar, orman altı ve açıklıkları, subalpin tundralar, stepler ve açık yerler gibi çok çeşitli habitatlarda yayılış gösteren kozmopolit bir familyadır (Pimenov ve Leonov 1993; Berenbaum 2001). Bu familya üyelerinin çoğu şemsiye biçimindeki çiçek durumu, bir karpofora asılı duran tek tohumlu iki karpelden (merikarp) oluşan özelleşmiş meyveleri ve çok sayıda küçük epigin çiçekleri ile kolaylıkla ayırt edilebilir (Downie ve ark., 1998). Ayrıca familya n=4–12 (genellikle 11 ya da 8) arasında sıralanan temel kromozom sayısıyla karakteristiktir (Plunkett ve ark., 1996a).
Apiaceae taksonlarının doğal kumarinler için iyi birer kaynak olduğu bilinmektedir (Doğanca ve ark., 1979). Kumarinler, uçucu yağlar ve seskiterpenler gibi yararlı sekonder metabolitleri içermeleri nedeniyle familya üyeleri sıklıkla baharat ve ilaç olarak kullanılırlar. Asya ülkelerinde kullanılan doğal ilaçların birçoğu Apiaceae üyelerinden elde edilen özütlerden oluşur. Örneğin Angelica türlerinin kurutulmuş
kökleri Japonya, Kore ve Çin’de ilaç olarak kullanılmaktadır. Bu bitkilerden sindirim rahatsızlıkların, baş ağrılarının, deri hastalıklarının tedavisinde ve bağışıklık sisteminin güçlendirilmesinde yararlanılmaktadır. Peucedanum ve Bupleurum da geleneksel tıpta kullanılan bitkilerdendir. Uzak Doğu’da P. japonicum türü diüretik, laksatif ve sedatif olarak kullanılırken B. falcatum türünün ise diare, amenore ve humma gibi hastalıkların tedavisinde kullanılmaktadır (Lee ve Rasmussen, 2000).
2.1. Apiaceae familayasını Morfolojik Özellikleri
Apiaceae familyası üyeleri genellikle internodlarda içi dolu ve kuvvetli gövdeli otsu bitkiler. Nadiren çalımsı bazen ağacımsı ya da ağaç formunda. Sukulent ya da değil. Yapraklar tabanda rozet şeklinde; gövdede almaçlı dizilmiş; tabanda yaprak kını bulunur; basit ya da bileşik bazen peltat, bileşik olduğunda ternat, pinnat, bipinnat ya da çok pinnat bazen de palmat; yaprak kenarı düz, parçalı, dikenli; stipul yok; yaprak büyüklüğü değişken; damarlanma pinnat, palmat ya da paralel. Birleşik veya basit umbel, nadiren simöz çiçek durumlu. Brakte var ya da yok. Çiçekler genellikle brakteollü; brakteoller küçük. Bitkiler genellikle hermafrodit, andromonoik, poligam ya da dioik. Kaliks indirgenmiş; indirgenmemişse serbest ya da birleşik, kesinlikle kaliks tüpü oluşmaz, sepal 5, çok küçük. Korolla serbest; petal 5; beyaz, sarı, pembe ya da eflatun; nadiren petalsiz. Stamen 5, perianttan bağımsız, sepallerle karşılıklı, fertil, tomurcukta içe dönük. Ovaryum alt durumlu (1-)2 bölmeli. Stilus 2, tabanı genişlemiş ve stilopodyum oluşur. Plesantasyon axillar ya da apikal. Ovül her bölmede 1 ya da 2. Meyve kuru, 2 merikarplı şizokarp. Her merikarp 1 tohumlu; merikarpların iç yüzleri birbirine bakar; arada birbirine bağlayan karpofor bulunur; dış yüz ise 5 birincil sırtlı, nadiren 4 ikincil sırt bulunur. Sırtlar arasında vitta (yağ kanalları) bulunur. Tohumlar yağlı endospermli (Heywood, 1978; Pimenov, 2004). Uçucu yağ var ya da yok. Reçineli ya da reçinesiz. Polenleri böcekler yoluyla yayılır (entomofil) (Lawrance, 1989). Çok soğuk bölgelerden tropikal bölgelere kadar yayılır. Tropik bölgelerin yüksek kesimlerinde özellikle kuzeyde ılıman bölgelerde yayılır (Doğan Güner, 2006).
2.2. Seseli Cinsi Üzerine Yapılan Moleküler, Morfolojik, Anatomik, Biyokimyasal ve Karyolojik Çalışmalar
Bu familya üzerinde morfoloji ve anatomi çalışmalarından, sitoloji ve bitki kimyası alanlarına kadar çok geniş bir yelpazede araştırmalar yapılmaktadır (Güner ve ark., 2000; İşcan ve ark., 2002).
Apiaceae familyası üyeleri dünyada ekonomik öneme sahip bitki gruplarındandır. Özellikle besin kaynağı olarak sebze ve hayvan yemi olarak kullanılırlar. Park ve bahçelerde süs bitkisi olarak kullanılan türleri mevcuttur. İçerdikleri alkoloidler ve reçineler nedeniyle tıpta (özellikle barsak rahatsızlıkları) ve kozmetikte yaygın kullanım alanlarına sahiptirler (Doğan Güner, 2006).
Diğer Apiaceae familyası üyeleri gibi Seseli cinsinin ekonomik değerinin de olduğu bilinmektedir. Genel anlamda cinsin taban yaprakları kurutularak hayvancılıkta besin olarak kullanılmaktadır. Avrupa’da bahçelerde ve parklarda süs bitkisi (S. gummiferum subsp. gummiferum) olarak kullanılmaktadır. Bütün Apiaceae cinslerinde olduğu gibi Seseli türlerinde bulunan kimyasallardan tıpta yararlanılmaktadır. Pakistan’da S. libanotis’in kimyasal çalışması yapılmış ve tohumlarından ekstrakte edilen uçucu yağların patojenik bakteriler (Staphylococcus
aureus, Echerichia coli, Shigella dysentry, Vibrio cholera ve Salmonella typhi)
üzerinde çok etkili olduğu bulunmuştur (Meena ve ark., 1989; Hu ve ark., 1990).
Apiaceae familyası üyeleri cinsleri çok farklı yapıda meyve örnekleri bulunduğundan araştırmacılar tarafında oldukça ilgi görmüş ve anatomik çalışmalarda sıklıkla kullanılmıştır (Pimenov ve Sdobnina, 1975; Pimenov ve ark., 1982; Tosun, 2002) ve yapısındaki eterik yağ moleküllerinden dolayı kimyasal analiz (Tosun, 2002; Glowniak, 1991) çalışmaları da mevcuttur.
Seseli türlerinin uçucu yağları ile yapılmış birkaç çalışmadan biri de, Seseli elatum L. toprak üstü kısmının uçucu yağı ile olan çalışmadır. Bu uçucu yağın,
analiz neticesinde seskiterpen hidrokarbonlarca çok zengin olduğu görülmüştür (Coassini Lokar, 1988).
Seseli ismi Yunanca’dan gelmektedir ve güçlü kokulu bitki anlamındadır (Doğan
Güner, 2006).
Diğer bir çalışmada Seseli campestre Besser'in, meyva ve herbası üzerinde gerçekleştirilmiştir. Clevenger apareyi kullanılarak yapılan distilasyonda uçucu yağ
miktarı meyvalarında % 1.5, toprak üstü kısmında ise % 1.0 olarak tespit edilmiştir (Tosun ve Özkal, 2003).
Bir diğer çalışmada İspanya'da endemik bir tür olan Seseli farreynii Molero ve Pujadas'ın taze yapraklarının dietileterli ekstresinin, GC-MS analizi sonucunda seskiterpenik bileşiklerin yanında monoterpenler, tetradesilasetat, tetrakozanol, hekzakozanol, oktakozanol gibi bileşiklere de rastlanmıştır (Muckensturm, 1997). Kinonik bileşiklerin izolasyonuna ait sadece bir çalışmaya rastlanmış olup, bu da İspanya'da endemik bir tür olan Seseli farreynii Molero ve Pujadas üzerinde yapılmış ilk kimyasal araştırmadır (Tosun ve Özkal, 2003).
Seseli elatum'a ait alt türlerden elde edilen izopimpinellinin yılanlara karşı
öldürücü bir zehir olduğu, ayrıca parazitik solucanlara karşı etkili olup, diüretik bir özelliğe sahip olduğu da literatürde kayıtlıdır (Coassini ve ark., 1988).
Biyokimyasal çalışmaların çoğu kumarinler üzerinde yoğunlaşırken, diğer birkaç farklı biyokimyasal bileşik üzerinde yapılmış çalışmalara da rastlanmıştır. Bu bileşikler terpenik, fenilpropanoit yapıdaki bazı bileşikler ile uçucu yağları kapsamaktadır (Tosun ve Özkal, 2003).
Doğan Güner’in (2006) revizyon çalışmasında 4 türün kromozom sayısı belirlenmiştir; Seseli gummiferum subsp. gummiferum (2n=20), S. gummiferum subsp. corymbosum (2n=20), S. resinosum (2n=20), S. peucedadanoides (2n=22). Literatür çalışmalarında ise Seseli cinsinin kromozom sayısı n= 9, n=10, n=11 olarak tespit edilmiştir. Aynı zamanda ülkemizde de yayılış gösteren Seseli resinosum Freyn & Sint. ve Seseli tortuosum L. türlerinin kromozom sayısı n= 11 olarak rapor edilmiştir (Grabiele ve ark., 2010).
Dünyanın farklı yerlerinde bilim dünyasına tanıtılan bazı Seseli türlerinde sayılan kromozom sayıları Seseli farrenyi J.Molero & J.Pujadas (2n = 18), Seseli
galloprovinciale Reduron (2n = 22), Seseli globiferum Vis. (2n = 22), Seseli longifolium subsp. intermedium Reduron (2n = 18), Seseli rhodopeum Velen. (2n =
42), Seseli rigidum Waldst. & Kit. (2n= 20), Seseli webbii Coss. (2n = 22) olarak sayılmıştır (Baltisberger ve Kocyan, 2010).
Ayrıca Seseli resinosum türünün embriyonik gelişimi bir doktora çalışmasına da konu olmuştur (Özveren, 2011).
Dünyada Seseli cinsi üzerine yapılan moleküler çalışmalar genellikle ITS üzerine yoğunlaşmış ve bu dizilerin bir kısmı NCBI ‘da (National Center for Biotechnology Information) da yayınlanmıştır.
Seseli webbii Coss. (GenBank: AY179037.1), Seseli peucedanoides
Koso-Pol.(GenBank: AY179034.1), Seseli hippomarathrum Pall. (GenBank: AY179033.1),
Seseli longifolium subsp. intermedium Reduron (GenBank: AY179032.1), Seseli tortuosum (GenBank: AY179031.1), Seseli galloprovinciale Reduron (GenBank:
AY179025.1), Seseli praecox Gamisans (GenBank: AY179024.1), Seseli gummiferum (GenBank: AY179023.1) türlerinin ITS1, 5.8 ribozomal RNA, ITS2 bölgeleri gen bankasında (NCBI) yayınlanmıştır.
2.2.1. Seseli Cinsinin Taksonomik Gelişimi
Seseli cinsi 1753 yılında Linnaeus’un Species Plantarum adlı eserinde 9 tür ile
temsil edilmektedir (Linnaeus, 1957). Bu türler S. pimpinelloides L., S. montanum L., S.
glaucum L., S. annuum L., S. ammoides L., S. tortuosum L., S. carviflora L., S. pyreneacum L. ve S. laxifragum L.’dur. Bunlardan S. tortuosum Türkiye’de de
yetişmektedir. Seseli tortuosum türü aynı zamanda cinsin lektotipidir (Pimenov and Leonov, 1993). Bu cinsin özellikleri ilk olarak 1754’de Linnaeus’un Genera Plantarum adlı eserinde verilmistir (Linnaeus, 1960).
Seseli ile ilgili birçok bilim adamının çalışmaları mevcuttur. Cins ile ilgili en
önemli çalışmalardan biri 1824’te W. Koch tarafından yapılmıştır (Koch, 1824). Bu çalışmada Koch, o zamana kadar tanımlanan Seseli türlerini toplu halde vermiş ve türleri brakte sayısı az ya da yok (S. elatum, S. montanum, S. glaucum, S. gracile, S.
annuum, S. varium, S. leucospermum, S. hippomarathrum, S. tortuosum, S. peucedanifolium, S. petraeum, S. cuneifolium, S. gummiferum, S. buchtornense) ve
braktesi çok sayıda olanlar (S. libanotis) olmak üzere 2 gruba ayırmıştır. Yine aynı yayında S. libanotis’i tanımlamıştır (sinonim: Athamantha libanotis).
1848 yılında Greiner ve Godron Flore De France’ı yazmışlar ve Seseli cinsini 2 seksiyona ayırmışlardır (Grenier and Gordon, 1848).
Seksiyon 1: Euseseli DC. S. tortuosum S. bocconii Guss. S. elatum L. S. montanum L. S. coloratum Ehrh. S. carvifolium
Seksiyon 2: Libanotis Crantz
S. libanotis
S. sibthorpii Godron & Gren.
Grenier ve Godron’un seksiyon olarak değerlendirdiği Libanotis, 1897’de Drude tarafından altcins olarak değerlendirilmiştir (Heywood, 1968).
1753, öncesi ve sonrasından günümüze kadar yapılan birçok çalışmada Seseli cinsinin sinonimleri, altcinsleri veya seksiyonları arasında görülen Libanotis cinsi farklı floralarda farklı kategorilerde ve farklı otör isimleriyle kullanılmaktadır. Örneğin Flora of U.S.S.R.’da Libanotis L. (Schischkin, 1973), Flora Europaea (Heywood, 1968) ve Flora Iranica’da (Rechinger, 1987) Libanotis J.Hill. ve Flora of Turkey’de ise Libanotis Hall. olarak görülmektedir. Yine Greiner ve Godron’un 1848 yılında yaptığı çalışmada
Libanotis seksiyonunun otörü Crantz’dır.
1950 yılında Flora of U.S.S.R’ın 16. cildine Seseli cinsini Shishkin yazmıştır (Schischkin, 1950). Bu çalışmada cinsin alt familyası, ordosu ve tribusu belirtilmiş ve cins 6 seksiyona ayrılmıştır. Bu seksiyonlardan Euseseli DC. 10 seride toplanmıştır. Aşağıda seksiyon ve serilerde sadece Türkiye’de bulunan türler verilmiştir:
Ordo: Umbelliflorae Bartl. Familya: Umbelliferae Moris. Alt familya: Apioideae Drude Tribus: Ammineae Koch Cins: Seseli L.
Seksiyon 1. Euseseli DC. Seri1. Stricta Schischk.
Seri 2. Varia Schischk.: S. grandivittatum (Somm.et Lev.) Schischk. Seri 3. Peucedanoidea Schischk.: S. peucedanoides (M.B.) K.-Pol. Seri 4. Glabrata Schischk.
Seri 5. Petraea Schischk.: S. petraeum Bieb.
S. gummiferum Pall.
Seri 6. Tortuosa Schischk.: S. campestre Bess.
S. andronakii
Seri 7. Eriocarpa Schischk. Seri 8. Fasciculata Schischk. Seri 9. Tenuisecta Schischk. Seri 10. Flavidae Schischk.
Seksiyon 2. Hippomarathroidea DC.
Seksiyon 3. Lomatopodium (Fisch et Mey.) Schischk. Seksiyon 4. Macrostylopodium Schischk.
Seksiyon 5. Erioscias Schischk.
Seksiyon 6. Pseudosilaus Schischk.: S. foliosum (Somm. et Lev.) Manden
Seseli ve diğer Apiaceae cinsleri üzerinde en yoğun araştırmalar M.G. Pimenov
tarafından yapılmıştır. 1975 yılında Pimenov ve L.I. Sdobnina Seseli cinsini 8 seksiyona ayırmıştır (Pimenov and Sdobnina, 1975).
Seksiyon 1. Condensatae M. Pimen. et Sdobn.
Seksiyon 2. Pseudolibanotis (Schischk.) M. Pimen. et Sdobn. Seksiyon 3. Libanotis (Hill.) Gren. et Godr.
Seksiyon 4. Sclerorhiza M. Pimen. et Sdobn. Seksiyon 5. Seseli L.
Seksiyon 6. Macrostylopodium Schischk. Seksiyon 7. Hippomarathroides DC.
Seksiyon 8. Lomatopodium (Fisch et Mey.) Schischk. Seksiyon 9. Pseudosilaus Schischk.
1985’te Pimenov ve Constance’ın yaptığı bir çalışma da cins üstü kategorileri düzenlemişlerdir (Pimenov and Constance, 1985). Buna göre:
Tribus: Seseleae Koch Subtribus: Seselinae Tausch
Genus: Seseli L., Sp. Pl. 1: 259 (1753).
Seseli cinsi üzerine daha önce belirtilenlerin dışında da sistematik çalışmalar
yapılmıştır (Schichkin, 1950;Pimenov, 1997; Hardvig and Strit, 1987). Sistematik çalışmaların dışında anatomik (Pimonov ve ark., 1982; Pimenov and Sdobnina, 1975;
Tosun, 2002). ve kimyasal analiz (Tosun, 2002; Glowniak, 1991) çalışmaları da mevcuttur. Fakat cins ile ilgili moleküler çalışma yoktur.
Dünyada 125-140 tür ve tür altı seviyede takson ile temsil edilen Seseli cinsi Avrupa (Kuzey Batı, Batı, Orta, Güney Batı, Güney, Güney Doğu, Doğu), Asya (Güney Batı, Kuzey, orta ve merkez, Batı, Güney Doğu, Güney, Kafkasya), Afrika (Kuzey, Kuzey Batı, Tropikal bölgeler), Kuzey Amerika ve Avustralya’da yayılış göstermektedir. (Davis,1972; Heywood, 1978;Pimenov and Leonov, 2004; Pimenov and Leonov, 1993;Heywood, 1978; Rechinger, 1987;Schischkin, 1950; Pignatti, 1982). Cins Türkiye’de 13 tür ve tür altı kategori ile ile temsil edilmektedir (Doğan Güner ve Duman, 2013).
Seseli cinsinin Türkiye’deki yayılışına bakıldığında özellikle Doğu Karadeniz,
Batı Karadeniz ve Antalya çevresinin tür sayısı açısından daha yoğun olduğu görülür. Taksonların fitocoğrafik bölgelere dağılımı; 8’i Avrupa-Sibirya, 3’ü Doğu-Akdeniz, 3’ü İran-Turan elementi ve 1’i geniş yayılışlı olarak değerlendirilmiştir (Doğan Güner, 2006).
Çok yıllıktır. Genelde tabanda ölü yaprakların saplarından oluşan lifli bir yapı bulunur. Bazı türlerde ise ölü yaprakların sapları kalıcıdır ama lifli yapı oluşturmaz. Bazen de hem lifli yapı hem de yaprak sapı seklinde bulunur. Gövde tek ya da çok sayıda olabilir; dallanma tabandan ya da ortadan baslar; gövde boyu 5-175 cm, bazı türlerde 3 m’ye kadar uzayabilir; gövde genellikle belirgin sırtlı nadiren düzdür; genelde yeşil nadiren kırmızımsı-mordur; tüylü ya da tüysüz olabilir. Yapraklar 1-4 pinnattır; tüylü ya da tüysüz olabilir; taban yaprakları gövde yapraklarından uzundur, daha fazla parçalanma gösterir; gövde yaprakları tabandan yukarı doğru sayıca azalır ve boyları küçülür, yaprak kını nadiren belirgin olarak bulunur. Çiçek durumu bileşik umbeldir; bazı türlerde ortadaki umbel yan umbellere göre daha gelişmiş olup ana umbel yapısı gösterir, ana umbelin ışın sayısı yan umbellere göre fazla ve ışın boyuda daha uzundur. Işınlar skabrid nadiren tüysüzdür. Braktenin varlığı türlere göre değişir, var ise tüylüdür. Brakteol bütün türlerde mevcuttur, serbest ya da tabanda birleşik, yoğun tüylüdür. Sepal yok ya da 5 tanedir, çok küçüktürler. Petaller 5, genellikle tomurcukta açık ya da koyu mor, nadiren beyaz ya da sarı, çiçek açtığında beyaz, orta damar mor çevresi beyaz ya da nadiren açık sarıdır; sırtta skabrid ya da tüysüzdür. Stamenler petallerle almaçlı dizilmiştir ve uzundur; beyaz ya da mor renklidir. Anterler versatil bağlı, sarı ya da mordur. Meyve oblong, oblong-ovoid, ovoid, linear-oblong, oblong-eliptik, eliptik-ovat
şekilli; 2-6,5 mm boyundadır; tüylü ya da tüysüzdür; belirgin 5 sırtlı, nadiren düzdür (Doğan Güner, 2006). Bu çalışmada kullanılan taksonların lokaliteleri aşağıdaki haritada işaretlenmiştir (Şekil 2.1).
Şekil 2.1. Seseli cinsinin Türkiye’deki yayılışı. : S. libanotis, : S. gummiferum subsp.
ilgazense, : S. corymbosum subsp. corymbosum, : S. transcaucasicum, : S. petraeum, : S. gummiferum subsp. gummiferum, : S. corymbosum subsp. phyrigium, : S.
resinosum, : S. tortuosum, : S. campestre, : S. marashica, : S. andronakii, : S.
serpentina, : S. hartvigii, : Seseli sp. nov.
Türkiye Florasında Artvin, Borçka’da yetiştiği belirtilen Seseli foliosum türü son yıllarda yapılan flora çalışmalarında ve Türkiye Seseli cinsinin revizyonu çalışmasında toplanamamıştır. Ancak Grossheim’in haritasında Gürcistan sınırından (Sovyet) toplandığı belirtilen bu türün ülkemizde olmadığı tespit edilmiştir. Yapılan ayrıntılı çalışmalar sonucunda Seseli grandivittatum türü, Trinia cinsine aktarılmıştır (Davis ve ark., 1988). S. peucedanoides (M.Bieb.) Koso-Pol. ve S. foliosum (Sommier & Levier) türleri Gasparrina cinsine aktarılmıştır (Pimenov and Kljuykov, 2010). Pimenov tarafından 2010 yılında yayınlanan Seseli paphlagonicum türü, Seseli gummiferum türünün sinonimi yapılmıştır (Doğan Güner ve Duman, 2013). Seseli phrygium Pimenov & Kljuykov türü, S. corymbosum Boiss. & Heldr. ex Boiss. subsp. phrygium (Pimenov ve Kljuykov) E.Doğan & H.Duman alttür seviyesine indirgenmiştir. Bunlara ilaveten S.
marashica E.Doğan & H.Duman ve S. serpentina B.L.Burtt ex H.Duman & E.Doğan
türleri bilim dünyası için yeni taksonlar olarak tanımlanmıştır (Doğan Güner ve Duman, 2013). Doğan Güner ve Duman (2013), Türkiye’deki Seseli cinsini de 13 takson (tür ve
alttür) altında toplamıştır. Üzerinde yayın çalışmaları devam eden bir yeni alt tür (Seseli gummiferum subsp. ilgazensis Ö.Çetin & A.Duran, M.Öztürk ( incelemede) tespit edilmiştir. Ayrıca 2013 yılında Tuz Gölünden toplanan bir Seseli örneğinin de tür düzeyinde yeni bir taksonu temsil ettiği düşünülmektedir. Bu örnekler de çalışmaya dâhil edilerek Türkiye’deki tüm Seseli taksonlarını kapsayan moleküler bir filogeni oluşturulması hedeflenmiştir.
2.2.2. Selineae Tribusunun Özellikleri
Drude Apiaceae familyasını Hydrocotyloideae, Saniculoideae ve Apioideae olmak üzere üç alt familyaya ayırmıştır ve bu durum familyanın sınıflandırma sistemine hâkim olarak kalmıştır (Ay, 2008).
Alt familyalardan en büyüğü ve ekonomik olarak öneme sahip olan Apioideae, diğer iki alt familyadan bileşik umbellere sahip oluşu, iyi gelişmiş vittalar (salgı kanalları) ve serbest karpofor ile ayırt edilebilir (Downie ve ark., 1998; Ay, 2008). Bileşik umbellanın yanı sıra çatallı bir karpoforla birleşmiş tek tohumlu iki merikarp ve stilopodiumdan çıkan stiluslar gibi morfolojik özellikler altfamilyanın monofiletik yapısını yansıtır ve Apioideae’yı Saniculoideae ve Hydrocotyloideae alt familyalarından sınırlar. Moleküler veriler bu tür morfolojik karakterlerin Apioideae alt familyasının monofiletik yapısını ortaya çıkardığını ve Apioideae’nın Saniculoideae alt familyasına kardeş bir grup olduğunu kanıtlamaktadır (Spalik ve Downie, 2001; Ay, 2008).
Ancak moleküler alandaki gelişmelerden sonra yapılan filogenetik çalışmalarla (Plunkett ve ark., 1996, 1997, 2004; Downie ve Katz-Downie 1999; Plunkett, 2001; Valiejo-Roman ve ark., 2002; Chandler ve Plunkett ve ark., 2004; Çelik, 2013). Drude’nin Hyrocotyloideae alt familyasının polifiletik olduğunun anlaşılması üzerine bu alt familyada değişiklik yapılmıştır (Plunkett ve ark., 2004). Yapılan değişikliklere göre
Hyrocotyloideae alt familyasına ait cinslerden Trachymene Rudge ve alt familyanın tip
cinsi Hydrocotyle L. Araliaceae familyasına aktarılmıştır. Geri kalan Hyrocotyloideae alt familyasına ait taksonlar yeni tanımlanan Azorelloideae Plunkett & Lowery ve
Mackinlayoideae Plunkett & Lowery alt familyalarına aktarılmıştır (Plunkett ve ark.,
2004; Liu ve ark., 2006, 2009, 2012; Nicolas ve Plunkett, 2009). Hyrocotyloideae alt familyasında yapılan bu değişikliklerden sonra Apiaceae familyasına ait taksonlar
Apioideae, Saniculoideae, Azorelloideae ve Mackinlayoideae olmak üzere dört alt
familyada toplanmıştır.
Tribus diğer taksonomik kategoriler gibi soyut bir kavramdır. Yer olarak familya ile cins arasında bulunur. Kelime anlamı oymak, kabile anlamlarına gelmektedir. Böyle bir kategoriye ihtiyaç duyulmasının sebebi ise bazı familyalardaki filogenetik karmaşıklığı çözümlemektedir. Diğer familyalarda da bulunmakla birlikte Apiaceae familyasının sistematiğinde oldukça önemli bir yere sahiptir. Apiaceae familyası meyve karakterleri bakımından çok çeşitlilik göstermektedir. Çeşitlilik daha fazla sistematik kategoriyi beraberinde getirmiştir.
Ayrıca filogenetik ağaçlarda bazı nodlara klad adı verilir. Kladlar, tribus ya da cinsler gibi taksonomik bir kategori değildir. Modern filogenetik sistematik, kladistik analize dayanmaktadır. Kladistiğe dayalı filogenetik ağaca kladogram denir. Bu ağaç çift yönlü çatallardan oluşur. Her dallanma noktası iki türün ortak bir atadan ayrılışını ifade eder. Bir kladogramdaki her bir evrimsel dala klad denir. Bir klad atasal bir tür ve bundan türeyen taksonlardan meydana gelir. Klad moleküler anlamda belli bir bilgiyi ifade eden bir gruptur. Diğer bir özelliği ise çalışmayı yapan araştırıcının belirlediği bir bilgidir. Bazen tribus içindeki cinslerin klada dâhil olduğunu da görmek mümkündür (Simpson, 2012).
Böyle cins, familya ya da daha yüksek bir taksondaki tür grubuna tek soydan anlamında ‘monofiletik’ denir. Birden fazla ortak soydan meydana geliyorsa ‘parafiletik’, eğer bir gruptaki türler, ortak bir ata içermiyorsa ‘polifiletik’ adını alır. Sistematikçiler bir kladogramda dallanma dizisine tüm benzerlik durumlarının ortak bir atayı işaret etmesine göre belirler. Ancak bu oldukça karmaşık bir uğraştır. Tüm bunların yanı sıra sistematikçiler moleküler verilerden de filogeniyi anlayabilirler. DNA dizisindeki nükleotidler kalıtsaldır ve moleküler düzeyde türlerin evrimsel dağılımı, bunların genom farklılıklarının birikimi ile paraleldir. İki türün ortak bir atadan dallanma hali ne kadar yakınsa, DNA ve aminoasit dizileri de o kadar benzer olmalıdır (Simpson, 2012).
Moleküler sistematik; Filogenetik ilişkilerin anlaşılması ve çok farklı türler arasındaki akrabalık derecelerinin izlenmesiyle soy ağacının en ince uçlarının anlaşılmasına yardımcı olur.
Selineae tribusunun cinsleri ile yapılmış çalışmalar incelenecek olursa Seseli ile yapılmış revizyon çalışmasında tribusun yeri aşağıdaki şekilde gösterilir;
Alem: Plantae Altalem: Tracheobionta Üstbölüm: Spermatophyta Sınıf: Dicotyledonae Altsınıf: Aralianae Takım: Apiales Familya: Apiaceae Altfamilya: Apioideae Oymak: Seseleae Altoymak: Seselinae Cins: Seseli (Doğan, 2006).
Yapılan kaynak taramaları sonucunda Türkiye Florası’nda Selineae tribusuna rastlanmamıştır (Davis, 1965-1985). İran Florası’na göre Selineae tribusu yoktur. Fakat bu tribus içindeki cinsler Apieae tribusu içerisinde yer alır. İran Florası’na göre Apieae tribusun özellikleri; çiçekler hermafrodit ya da tek eşeyli, petaller beyaz, pembe ya da sarı, stilopodyum konik ya da gömülmüş durumda, meyve oval ve uzamış, yanları hafif kanatlı, merikarp sırtları hafif belirgin ya da belirgin olmayan kanatlı, komisürler beyazımsı-krem, ± düz (Rechinger, 1987).
Rus Florası’nda ise yine Selineae tribusu bulunmamaktadır. Bunun yerine Selineae içerisindeki cinsler Ammineae (Koch, 1824) tribusuna dâhil olarak gösterilmektedir. Buna göre Ammineae tribusunun özellikleri; Çiçekler iki eşeyli nadiren tek eşeyli, kaliks dişli ve iyi gelişmiş ya da yok, petaller içe doğru kavisli ve lobları mevcuttur genellikle oval veya obkordattır, stilopodyum konik ya da yastık şeklinde, meyve yanlardan hafifçe basık; silindirik, oval ya da oblong-silindirik olabilir, valekul kanalları ise 1-4 arasında değişir, enine kesitleri dairesel ya da beşgen şeklinde ve beyaz renkli, perikarp pürüzsüz ya da tüylü, enine kıvrımlı, genellikle komissüre doğru kristaller bulunmaz (Schischkin, 1950).
2.2.3. Selineae Tribusunun Moleküler Çalışmalara Göre Taksonomik Yeri
Bitkilerde üç temel tip DNA dizisi, üç ana DNA kaynağından köken alır; nüklear DNA(nDNA), kloroplast DNA (cpDNA) ve mitokondriyal DNA (mtDNA). Nüklear DNA atalardan yeni nesillere hiç şüphesiz eşeyli veya eşeysiz üremeyle çekirdek
bölünmesi yoluyla aktarılmaktadır. Diğer taraftan kloroplast ve mitokondri, çekirdekten bağımsız olarak replike olur, bölünür ve yeni nesillere farklı bir şekilde aktarılır. Örneğin Angiospermlerde bu organeller genellikle (bazı istisnalarla beraber) dişi gamet yoluyla aktarılır, dolayısıyla yumurtayla aktarılırken sperm hücreleriyle aktarılmaz. (Kozalaklılarda, ilginç bir şekilde kloroplast DNA, dişi gamet yoluyla değil erkek gamet yoluyla aktarılır). Kloroplast DNA’ sından elde edilen dizileme verilerinin kullanımının hem uzak hem de yakın akrabalıkları belirlemede faydalı olduğu kanıtlanmıştır. Bütün organel ve prokaryotik DNA’ larda olduğu gibi, kloroplast DNA’sı da halkasaldır. İlginç şekilde çoğu Angiospermlerde, ters tekrar olarak adlandırılan, ayna görüntüsü şeklinde kloroplast DNA’sı bulunmaktadır. Kloroplast DNA’nın kodlayan genlerine ilave olarak genler arası bölgeler olarak bilinen genler arasındaki dizilerde filogenetik analizlerde kullanılabilir. Genler arası bölgeler, kodlayan genlere kıyasla özellikle daha büyük varyasyonlar gösterir; bu nedenle genler arası bölgeler, tür ve tür altı taksonlar gibi daha düşük düzeydeki taksonomik analizler için faydalıdır (Simpson, 2012).
Apiaceae familyasının gen merkezi Anadolu kabul edilir. Türlerin dağılımı homojen olmamakla birlikte monofiletik ve polifiletik olarak değişiklik gösterir (Davis ve Hedge, 1975).
Polifiletik olduğu kabul edilen cinslerden bazılarının tür sayıları şöyledir; Ferula 177, Ligusticum 40-50, Seseli 100-120 ve Peucedanum 100-120. Çok türle temsil edilen ve oldukça geniş yayılışlı olan bu cinslerin teşhisi zorlaşmış ve bu cinslerin tanımlanması yetersiz kalmıştır (Spalik ve ark., 2004).
Altıncı Apiales sempozyumuna göre, çalışmalardaki ITS sekansları temel taksonların tamamını kapsayacak kadar kapsamlı değildir. Moleküler veri seti ile taksonomi merkezli veri filogenetik sonuçlar arasında uyumluluk açısından farklı bir moleküler veri setleri (cpDNA genleri ve intron sekansları, cpDNA sınırlanmış alanlar, ITS sekans) kullanılmıştır (Plunkett ve Downie 1999; Downie ve ark., 2000; Downie ve ark., 2001).
Aşağıdaki 10 tribusun monofiletik olduğu teyit edilmiştir. Aciphylleae, Bupleureae, Careae, Coriandreae, Echinophoreae, Oenantheae, Pleurospermeae, Pyramidoptereae, Scandiceae (alt tribusları içinde Daucinae, Ferulinae, Scandicinae, Torilidinae ve Glaucosciadium kladı) ve Smyrnieae. Buna ek olarak önceki sonuçlar ve şimdiki filogenetik analizler sadece ITS sekansları destek alınarak 4 monofiletik tribus belirlenmiştir. 1- Apieae, 2- Selineae 3- Tordylieae 4- Pimpinelleae.
Downie ve arkadaşlarının yaptığı çalışmaya göre majör klada dâhil 12 cinsten,
Arcuatopterus, Chamaesium, Conium, Diplolophium, Erigenia monofiletik; Acronema, Cachrys, Conioselinum, Komarovia, Opopanax, Physospermopsis, Sinodielsia ise
polifiletik olduğu düşünülmektedir (Downie, 2010).
Yapılan diğer çalışmalarda ise Selineae tribusunun “Arracaciea” kladına dâhil olduğunu kabul etmişlerdir. Selineae tribusu ile birlikte Careae, Echinophoreae ve Pyramidoptereae tribusları ve Apium, Heracleum ve Pimpinella kladları toplu olarak Apioid süperkladının sadece bir kısmını oluşturmaktadır (Plunkett ve Downie 1999; Downie ve ark., 2000; Downie ve ark., 2001).
Apioid süperkladının monofiletik olduğu hem sekans sonuçları hem de
restriksiyon verileri ile ispatlanmıştır (Downie ve ark., 2000). Ayrıca monofiletik çalışmalar sadece yapısal plastid genomlarının sekansları da çıkarılarak aynı sonuçlar desteklenmiştir (Plunkett ve Downie 1999; Downie ve ark., 2000; Downie ve ark., 2001).
Downie ve ark. (2001), Üçüncü Uluslar Arası Apiales Sempozyumu’nda Apioideae alt familyası ile ilgili yayınlanmış olan moleküler sistematik çalışmaları özetleyerek bu alt familya ile ilgili revize edilmiş sınıflandırmayı sunmuşlardır. Bu sınıflandırma, kloroplast genleri (rbcL, matK) ve intron dizileri (rpoC1, rps16,rpl16), çekirdek ribozom DNA’sı ITS gibi farklı moleküler çalışmalardan elde edilen taksonomik sonuçlara dayandırılmıştır. Bu sınıflandırma sonrasında 10 tribus ve 3 subtribus belirlenmiştir (Çizelge 2.1.) (Downie ve ark., 2001; Çelik, 2013).
Çizelge 2.1. Apioideae tribus ve subtribusları (Downie ve ark., 2001). Altfamilya: Apioideae Drude
Tribus: Aciphylleae M.F.Watson & S.R.Downie Tribus: Bupleureae Spreng.
Tribus: Careae Baill.
Tribus: Echinophoreae Benth.
Tribus: Heteromorpheae M.F.Watson & S.R.Downie Tribus: Oenantheae Dumort.
Tribus: Pleurospermeae M.F.Watson & S.R.Downie Tribus: Pyramidoptereae Boiss.
Tribus: Scandiceae Spreng. Subtribus: Daucinae Dumort.
Subtribus: Scandicinae Tausch. Subtribus: Torilidinae Dumort. Tribus: Smyrniae Spreng.
Downie ve ark., yaptığı bu çalışmaya göre Selineae tribusu içine dâhil olan
Seseli ve diğer bir çok cins Angelica kladı içinde gösterilmiştir. Downie ve arkadaşları
kladları 7 ayrı grup yaparak ayırmışlardır;
Klad: Angelica Klad: Apium Klad: Arraracia Klad: Conioselinum Klad: Heracleum Klad: Komarovia
Klad: Pimpinella (Downie, 2010).
Downie’nin Selineae tribusunu kapsayan çalışmasında önceki tribus Pyramidoptereae ve sonraki tribus Tordyliinae yer almıştır. Bu çalışmada kloroplast genleri baz alınarak böyle bir filogenetik ağaç çıkarılmıştır. Downie ve arkadaşları tribus ve subtribus seviyesinde konumlama yapmak için öncelikle ITS verilerini baz almaya çalışmışlardır. En az 94 cinsle temsil edilen Selineae tribusu Apioideae tribusu içinde en büyük klad olma özelliğine sahiptir. Bu çalışmaya göre çıkan filogenetik ağaç aşağıdaki gibidir (Şekil 2.2.) (Downie, 2010).
Şekil 2.2. Selineae tribusunun filogenetik ağaçta gösterimi (Downie, 2010).
III. Apiales sempozyumu 1999 yılında Amerika’da yapılmıştır. Downie ve ark. (2001), Apioideae alt familyası ile ilgili moleküler sistematik çalışmaları özetleyerek bu alt familya ile ilgili revize edilmiş sınıflandırmayı sunmuşlardır. Bu sınıflandırma, kloroplast DNA’sına ait kloroplast genleri (rbcL, matK); intron dizileri (rpoC1, rps16,
rpl16) ve çekirdek ribozomal DNA’sında ITS sekansları ile kloroplast genomundaki
yeniden yapılandırılmadan elde edilen veriler alt familya içerisindeki büyük kladlarların aydınlatılmasında kullanılmıştır. Fikir birliğine varılan 12 büyük kökenden 6 tanesi tribus seviyesinde öngörülmüştür. Bunlar Bupleureae, Heteromorpheae, Oenantheae, Pleurospermeae, Smyrnieae ve Scandiceae’dir. Heteromorpheae ve Bupleureae temel
kladları oluşturmaktadır, diğer kalan tribuslar arasındaki ilişkiler net değildir. Scandicinae tribusu, Daucinae ve Torilidinae alt tribuslarını içermektedir. İkinci olarak geleneksel anlamda iki taksonu Caucalideae içerisinde yer almaktadır (Downie ve ark., 2001).
III. Apiales sempozyumunda birçok moleküler veri birleştirilmiş ve Selineae tribusu ve buna ek olarak 22 cinsin soy ağacındaki yeri tam olarak belirlenememiştir. Daha sonraki ITS tabanlı çalışmalar ise daha olumlu sonuçlar vermiştir (Downie ve ark., 1999).
Selineae tribusuna dâhil olan pek çok cins Drude (1897-1898)’nin yapmış olduğu klasik sınıflandırma sistemi içinde Peucedaneae tribusunun Angeliceae ve Ferulinae alt tribusunda veya Pimenov ve Leonov (1993)’un sınıflandırmasında Peucedaneae ve Angeliceae tribuslarında düşünülmüştür. Downie ve ark. (2001) moleküler verileri esas alarak yapmış olduğu yeni sınıflandırma çerçevesinde bu taksonların hepsi Angelica ve Arraracia isimli kladlar içerisinde iki gayri resmi grupta yer almıştır. Arraracia kladının filogenetik pozisyonu henüz net olmamasına rağmen, daha önce yapılmış birçok çalışmada Angelica kladı içinde ortaya çıkmıştır (Plunkett ve ark., 1996, Downie ve ark., 1998). Downie ve ark. (2001), Angelica kladının oldukça güçlü (%97-98 bootstrap değerleri) desteklenmesinden dolayı bu kladı tribus olarak önermişlerdir (Spalik ve ark., 2004).
Kuzey Amerika Apioideae altfamilyasına 19 cinse ait 137 ve Angelica kladına ait 22 dış grubun, Kuzey Amerika endemik taksonlarındaki monofilisini değerlendirmek ve yeri belli olmayan Cymopterus grup içindeki yerini belirlemek için ITS sekanslarıyla filogenetik analizleri yapılmıştır. Sonuçlara göre çok yıllık, endemik Kuzey Amerika Apioid umbelliferlerini içeren zayıf bir şekilde desteklenen monofiletik bir grup olduğu, bunun yanısıra Angelica kladı ve meso-Amerikan Arracacia kladının birçok olası kardeş klad bağlantısının ikisini içermektedir (Downie ve ark., 2010).
V. Apiales sempozyumu 2005 yılında Avusturya (Viyana) da yapılmış olup daha fazla filogenetik çalışmalar ile Oenantheae tribusunun yerinin daha iyi belirlenmesi için buna yönelik çalışmalar şeklinde gelişmiştir (Downie ve ark., 2010). 17 cinsle tanınan Oenantheae tribusunda bu çalışma sayesinde, 4 cinsin monofiletik olmadığı ortaya çıkmıştır. Bu cinsler Berula, Bifora, Cryptotaenia ve Sium (Downie ve ark., 2010).
Son yıllarda yapılan çalışmalardan biri de İran Apiaceae familyasının ITS sekanslarıyla desteklenerek yapılan filogenetik analizlerdir. Buna göre Johrenia ve
Johreniopsis monofiletik olmadıkları ispatlanarak Selineae tribusu içine aktarılmışlardır. Buna ek olarak Ferulopsis Kitag., Trinia Hoffm. ve Vicatia DC. cinsi de Selineae tribusu içine aktarılmıştır. Önceki çalışmalarda göz önüne alınarak Selineae tribusu “monofiletik değildir” sonucuna varılmıştır (Ajani ve ark., 2008).
Downie’nin major kladları tanıttığı çalışmasında Selineae tribusu olarak gösterdiği cinsler aşağıdaki şekilde gösterilmiştir (Çizelge 2.2). *Pachypleurum
mutellinoides ise Selineae tribusu içerisinde tür kategorisinde değerlendirilmiştir
(Downie, 2010).
Çizelge 2.2. Selineae tribusu içindeki cinsler (Downie ve ark., 2010).
Aethusa Dimorphosciadium Katapsuxis Oreocome Saposhnikovia
Ammoselinum Dystaenia Kedarnatha Oreoselinum Seseli
Angelica Endressia Kitagawia Ormosolenia Spermolepis
Apiastrum Exoacantha Ledebouriella *Pachypleurum
mutellinoides Stenocoelium
Carlesia Ferulopsis. Libanotis Paraligusticum Thecocarpus
Cervaria Glehnia Ligusticopsis Peucedanum Thysselinum
Chymsydia Imperatoria Ligusticum Phlojodicarpus Tommasinia
Cnidiocarpa Johrenia Lomatocarpa Pilopleura Trinia
Cnidium Kadenia Macrosciadium Pteroselinum Vicatia
Cortia Kailashia Magadania Rumia Xanthogalum
Cortiella Karatavia Oligocladus Sajanella Xanthoselinum
Dichoropetalum
Yapılan son çalışmalardan biri olan altıncı Uluslararası Apiales Sempozyumu 2008 yılında Rusya (Moskova) da düzenlenmiştir. Downie ve ark., Apiaceae familyasının Apioideae alt familyasının ITS tabanlı sınıflandırmasını ele almışlardır.
Bu çalışmalara göre, şu anda ve muhtemelen yakın gelecekte nrDNA ve ITS sekanslarının çalışılması Apioideae familyası için en kapsamlı verileri oluşturacaktır. Bu sekansların ortaya çıkarılması için kullanılacak DNA eski herbaryum materyallerinden bile kolayca elde edilebilir. Downie ve arkadaşları çalışmaları sonucu türlerin kendi arasındaki az miktarda polimorfizmin filogenetik ilişkiyi tahmin etmeye engel teşkil etmediğini söylemişlerdir. Apioidea alt familyasına ait filogenetik analizler için 1240 ITS sekansı gen bankalarında bulunan ve henüz yayınlanmamış sekansların da kullanılmasıyla monofiletik gruplar belirlenmiştir (Downie ve ark., 2008). Ayrıca bu monofiletik grupları güvenli kılmak için kloroplast sekansı da test edilmiş ve çalışmaya eklenmiştir. Yapılan çalışmada 5.8 S gen bölgesi kullanılmıştır. Bu 1240 ITS sekansı 292 cins ve 959 tür ile yapılmıştır ve Apioidea familyasının coğrafi yayılışını