ICAM-1 PROTEĠNĠNE KARġI DNA APTAMERLERĠNĠN GELĠġTĠRĠLMESĠ
Deniz YĠĞĠT Yüksek Lisans Tezi Biyomühendislik Anabilim Dalı Dr. Öğr. Üyesi Abdullah Tahir BAYRAÇ
T.C.
KARAMANOĞLU MEHMETBEY ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
ICAM-1 PROTEĠNĠNE KARġI DNA APTAMERLERĠNĠN GELĠġTĠRĠLMESĠ
YÜKSEK LĠSANS TEZĠ Deniz YĠĞĠT
Biyomühendislik Anabilim Dalı
Dr. Öğr. Üyesi Abdullah Tahir BAYRAÇ
ii
TEZ BĠLDĠRĠMĠ
Yazım kurallarına uygun olarak hazırlanan bu tezin yazılmasında bilimsel ahlak kurallarına uyulduğunu, baĢkalarının eserlerinden yararlanılması durumunda bilimsel normlara uygun olarak atıfta bulunulduğunu, tezin içerdiği yenilik ve sonuçların baĢka bir yerden alınmadığını, kullanılan verilerde herhangi bir tahrifat yapılmadığını, tezin herhangi bir kısmının bu üniversite veya baĢka bir üniversitedeki baĢka bir tez çalıĢması olarak sunulmadığını beyan ederim.
iii ÖZET
Yüksek Lisans Tezi
ICAM-1 PROTEĠNĠNE KARġI DNA APTAMERLERĠN GELĠġTĠRĠLMESĠ Deniz YĠĞĠT
Karamanoğlu Mehmetbey Üniversitesi Fen Bilimleri Enstitüsü
Biyomühendislik Ana Bilim Dalı
DanıĢman: Dr. Öğr. Üyesi Abdullah Tahir BAYRAÇ Temmuz, 2019, 93
ICAM-1, endotelyal ve epitel hücreleri, vasküler düz kas hücreleri ve bazı lökosit alt kümeleri gibi çok sayıda hücre tipinin hücre yüzeyinde ifade edilen, insanlarda ICAM-1 geni tarafından kodlanan ve immünoglobulin süper ailesine ait bir transmembran glikoproteinidir. ICAM-1, endotel hücrelerinde ve bazı lenfositler ve monositler üzerinde yapısal olarak düĢük seviyelerde ifade edilmesine rağmen, sitokinlerin ve reaktif oksijen türlerinin varlığında ekspresyonu önemli ölçüde artmaktadır. ICAM-1, hayatta kalma, göç, ekstravazasyon, ve metastaz gibi kansere bağlı biyolojik süreçleri mümkün kılar. ICAM-1 çeĢitli kanser hücrelerinde ifade edilir ve ifade edilen ICAM-1‟in göğüs, akciğer gibi kanser türleri için potansiyel bir biyobelirteç olduğu düĢünülmektedir. Kanser büyük bir halk sağlığı sorunudur ve metastatik kanser, kanser ölümlerinin büyük bir çoğunluğunu oluĢturur. Erken tanı ve uygun metastatik kanser tedavisi, sağkalım oranını büyük ölçüde uzatabilir ve hastalar için yaĢam kalitesini artırabilir. Hücre yüzey proteinleri hücrelerin ilk eriĢilebilir bölümleri olduğundan, nispeten kolaylıkla tespit edilebilir. Bu nedenle, hücre yüzeyi biyobelirteçleri kanserin aĢamasının erken ve kolay saptanması, farklılaĢması ve belirlenmesi için uygundur. Aptamerler yüksek afinite ve seçicilik özelliklerine sahiptir. Kimyasal bileĢikler, proteinler, hücreler gibi geniĢ hedef yelpazesine yüksek bir etkinlik ve özgünlük ile bağlanabilen tek zincirli kısa oligonükleotit dizileridir. Yapılan çalıĢmalarda moleküler tanı ve tedavilerde kullanımlarında yüksek potansiyele sahip oldukları belirtilmiĢtir. Bu çalıĢmada da ICAM-1 aracılığıyla metastazı engellemek için, Protein-SELEX yöntemini kullanarak ICAM-1‟a yüksek etkinlik ve özgünlükte bağlanabilen aptamer dizileri geliĢtirilmiĢtir.10 turluk protein-SELEX ve sonrasında zenginleĢen dizileri belirlemek için yeni nesil dizileme yapılmıĢtır. En çok tekrar eden aptamerlerin arasında üç aday aptamer (BD-10-1, D-10-2 ve D10-3) seçilmiĢ ve Kd değerleri
hesaplanmıĢtır. Bu aday aptamerler, CHO-ICAM-1 hücresine yüksek özgüllük göstermiĢtir fakat negatif seçilim için kullanılan MIA PaCa-2 hücresineyüksek bir bağlanma afinitesi göstememiĢtir.
iv ABSTRACT
MsThesis
DEVELOPMENT OF DNA APTAMERS AGAĠNST ICAM-1 PROTEĠN Deniz YĠĞĠT
Karamanoğlu Mehmetbey University Graduate School of Natural andAppliedSciences
Department of Bioengineering
Supervisor: Assist. Prof. Dr. Abdullah Tahir BAYRAÇ July, 2019, 93
ICAM-1 is a transmembrane glycoprotein of immunoglobulin superfamily that is coded by ICAM-1 gene in humans and expressed on cell membranes of various cell types such as endothelial and epithelial cells, vascular smooth muscle cells, and some leukocyte subgroups. Although ICAM-1 is expressed in low levels on endothelial cells, some lymphocytes and monocytes, the expression levels increase significantly in presence of cytokines and reactive oxygen species. ICAM-1 enables cancer related biological processes like survival, migration, invasion and extravasation. ICAM-1 is expressed in various cancer cells and it is thought to be a potential biomarker for lungs and breast cancers. Cancer is an important public health problem and metastatic cancer leads to majority of cancer deaths. Early diagnosis and appropriate metastatic cancer therapies could prolong survival on a large scale and improve life quality. Cell membrane proteins are considered to be targeted easier since they are on the first part that could be reached. For that reason, cell membrane biomarkers are suitable for early and simple diagnosis of cancer, stages of cancer, and malignant transformation. Aptamers have properties of high affinity and selectivity for their targets. They are single stranded oligonucleotide sequences that can bind to large scale of targets involving chemical compounds, proteins and cells, effectively and specifically. In recent studies, it was shown that they have a very high potential in use of molecular diagnoses and therapies. In this study, highly effective and specific aptamer sequences for ICAM-1 were generated with protein-SELEX method in order to prevent metastasis. Next generation sequencing was utilized to determine enriched sequences at the end of a 10 cycle protein-SELEX. Three aptamer candidates (BD-10-1, D-10-2 and D10-3) were selected from the most repeated sequences and Kd values were calculated. The aptamer candidates show high affinity to CHO-ICAM-1 cells but
low affinity to negative control cell line MIA PaCa-2.
v ÖNSÖZ
Yüksek Lisans eğitimim boyunca bana yol gösteren, çalıĢmamın her aĢamasında tecrübesini, yardımlarını esirgemeyen, maddi ve manevi destek olan değerli hocam ve danıĢmanım sayın Dr.Öğr. Üyesi Abdullah Tahir Bayraç‟a,
Motivasyonuve desteğiyle yanımda olansayın hocam Dr.Öğr. Üyesi Ceren Bayraç‟a, Tüm laboratuvar çalıĢmalarım boyunca yardımcı olansayın hocam ArĢ.Gör. Berke Bilgenur ġener‟e,
Desteğini, yardımlarını hiçbir zaman esirgemeyen, hep yanımda olan canım dostum Mukaddes KeskinateĢ‟e,
Hayatım boyunca beni maddi ve manevi olarak destekleyen, akademik kariyer yapmamı en az benim kadar isteyen aileme; her zaman varlığını hissettiren canım babam Mehmet Yiğit‟e, bana hep Ģevkatle sevgiyle yaklaĢan canım annem ġenay Yiğit‟e ve sadece kardeĢim değil en iyi arkadaĢım olan canım kardeĢim Gökhan Yiğit‟e çok teĢekkür ederim.
Bu çalıĢma, 215E157 (AptaSELECT) numaralı araĢtırma projesi kapsamında TÜBĠTAK tarafından desteklenmiĢtir. Desteklerinden dolayı TÜBĠTAK‟a teĢekkür ederim.
Deniz YĠĞĠT Karaman-2019
vi ĠÇĠNDEKĠLER Sayfa ÖZET... iii ABSTRACT ... iv ÖNSÖZ ... v ĠÇĠNDEKĠLER ... vi ÇĠZELGELER DĠZĠNĠ ... viii ġEKĠLLER DĠZĠNĠ ... ix SĠMGELER VE KISALTMALAR DĠZĠNĠ ... xi 1. GĠRĠġ ... 1
2. KURAMSAL TEMELLER VE KAYNAK ARAġTIRMASI ... 3
2.1. Kanser ... 3
2.1.1. Kanser nedenleri ... 4
2.1.2. Kanser belirtileri ... 5
2.1.4. Kanser tedavi yöntemleri ... 5
2.1.5. Kanser türleri ... 6 2.1.6. Kanser Ġstatistikleri ... 7 2.2. Metastaz ... 7 2.2.1. Metastaz belirteçleri... 10 2.3. ICAM-1 ... 11 2.3.1. ICAM-1‟ın Yapısı ... 13
2.3.2. ICAM-1‟ın Kanserdeki Rolü... 15
2.3.3. ICAM-1 Ġfadesine Etki Eden Sinyal Yolakları ... 17
2.4. Aptamer ... 20
2.4.1. Aptamer Tabanlı Hedefli Kanser Tedavileri ... 27
2.5. SELEX ... 28
2.5.1 Hedef Seçimi ve Kütüphane Tasarımı ... 29
2.5.2 Seleksiyon ve Amplifikasyon ... 29
2.5.3. Dizileme ve Karakterizasyon ... 31
2.5.4 SELEX‟in GeliĢim Süreci ve ÇeĢitleri ... 31
3. MATERYAL VE METOT ... 39
vii
3.2. Metot ... 40
3.2.1. DNA Kütüphanesi ve Primer Tasarımı ... 40
3.2.2. DNA Kütüphanesi PZR ġartlarının Optimizasyonu... 40
3.2.3. Proteinin Ġmmobilizasyonu ... 41
3.2.4. SELEX ... 42
3.2.5. Polimeraz Zincir Reaksiyonu ... 44
3.2.6. Dizileme (Aday Aptamerlerin Sekanslanması) ... 45
3.2.6 Biyoinformatik ... 51
3.2.7. Hücre kültürü... 51
3.2.8. Flow Sitometri ... 52
3.2.9. Bağlanma Afinitesinin Belirlenmesi ... 52
4. BULGULAR ve TARTIġMA ... 54
4.1. Protein Ġmmobilizasyonu ... 54
4.2. PZR Optimizasyonunu ... 54
4.3. SELEX Turları... 55
4.4. Dizileme (Aday Aptamerlerin Sekanslanması) ve Biyoinformatik Analizleri 57 4.5. Flow Sitometri ... 63
4.6. Bağlanma Afinitesinin Belirlenmesi ... 66
5. SONUÇLAR ... 69
KAYNAKLAR ... 70
viii
ÇĠZELGELER DĠZĠNĠ
Çizelge Sayfa
Çizelge 2- 1 Kansere neden olan risk faktörleri ... 5
Çizelge 2- 2 ICAM-1‟ın topolojisi ve molekül yapısı ... 14
Çizelge 2- 3 Aptamer- antikor arasındaki temel farklılıklar ... 27
Çizelge 2- 4 Hücre-SELEX ile belirlenen aptamerler ... 35
Çizelge 2-5 Aptamerlerin tanımlanması için SELEX'te en yaygın kullanılan hedef proteinler (Stoltenburg ve ark., 2007) ... 38
Çizelge 3-1 PZR koĢulları ... 41
Çizelge 3- 2 SELEX turlarına ait bilgiler ... 44
Çizelge 3- 3 Kullanılan PZR ısı değiĢim Ģartları ... 45
Çizelge 3- 4 Kullanılan PZR içerik ve miktarları ... 45
Çizelge 3- 5 SELEX 7,9 ve 10‟un Ġllumina primerler ile PZR yapılması ve elde edilen PZR ürünlerinin Ampure XP küreleri ile temizlenmesi için kullanılan koĢullar ... 48
Çizelge 3- 6 Ġllumina primerler ile yapılan PZR ve Ampure XP küreleri ile temizlenme 48 Çizelge 3- 7 SELEX 7,9 ve 10‟un Ġndeks primerler ile PZR yapılması ve elde edilen PZR ürünlerinin Ampure XP küreleri ile temizlenmesi için kullanılan koĢullar ... 49
Çizelge 3- 8 Ġndeks primer dizisi ... 50
Çizelge 4-1 SELEX turlarının optimum döngü sayıları ... 57
Çizelge 4-2 Ġllümina PZR ürünlerinin AmpureXP küreleri ile temizlenmesi sonrası kalite ölçümleri ... 59
Çizelge 4-3 Dizileme sonrası okunan dizi sayıları ... 59
Çizelge 4-4 Seçilen aday aptamerler ... 62
Çizelge 4-5 Seçilen aptamerlerin floresans yoğunlukları ... 66
ix
ġEKĠLLER DĠZĠNĠ
ġekil Sayfa
ġekil 2- 1Kanser hücresinin özellikleri ... 4
ġekil 2-2 Kanser tedavi evriminin basamakları ... 6
ġekil 2-3 Metastazın aĢamaları ... 8
ġekil 2-4 ICAM-1 molekül yapısı ... 15
ġekil 2- 5 Metastazda MUC-1 ve ICAM-1 ĠliĢkisi ... 18
ġekil 2- 6 Metastazda LFA-1 ve ICAM-1 ĠliĢkisi ... 20
ġekil 2- 7 Aptamerin hedefe bağlanması ... 23
ġekil 2- 8 Aptamerlerin tanımlanması için SELEX'te kullanılan hedefler... 29
ġekil 2-9 Protein SELEX ... 37
ġekil 3- 1 PZR ürünlerinin tek zincirli hale getirilmesi için kullanılan teknik ... 43
ġekil 3- 2 Protein(ICAM-1)-SELEX döngüsü ... 44
ġekil 3- 3 Dizlilemede kullanılan illümina primerler ... 46
ġekil4-1 Protein immobilizasyonu ... 54
ġekil4-2 Döngü optimizasyonu jel görüntüsü ... 55
ġekil4-3 Agaroz jel elektroforezlerde kullanılan ladder ... 55
ġekil4-4 ICAM-1 SELEX 1 ve SELEX 2 için döngü optimizasyonu jel görüntüleri ... 56
ġekil4-5 ICAM-1 SELEX 3 ve SELEX 4 için döngü optimizasyonu jel görüntüleri... 56
ġekil4-6 ICAM-1 SELEX 5 ve SELEX 6 için döngü optimizasyonu jel görüntüleri ... 56
ġekil4-7 ICAM-1 SELEX 7 ve SELEX 8 için döngü optimizasyonu jel görüntüleri ... 56
ġekil4-8 ICAM-1 SELEX 9 ve SELEX 10 için döngü optimizasyonu jel görüntüleri... 57
ġekil4-9 SELEX 7, 9 ve 10‟un amplikon PZR jel görüntüleri ... 58
ġekil4-10 SELEX 7, 9 ve 10‟un amplikon PZR ürünlerinin AmpureXP küreleri ile temizlenmesi sonrası jel görüntüsü ... 58
ġekil4-11 Primer bölgeleri kesilen dizilerin MAFFT diagramı ... 60
ġekil4-12 Seçilen overrepresented dizilerin MAFFT programındaki filogenetik ağacı ... 60
ġekil4-13 Aday aptamerlerin 37°C'de oluĢturdukları ġekiller ... 61
ġekil4-14 Sentezlenen aday aptamerlerin 37°C ve 4°C'deki ikincil yapıları (B10-1, D-10-2 ve D-10-3) ... 62
ġekil4-15 15 MIA PaCa-2, CHO-ICAM-1, kontrol aptameri ve aday aptamerlerin FL1-H histogramı ... 64
x
ġekil4-16 CHO-ICAM-1 ile inkübe edilmiĢ kontrol aptameri ve BD-10-1 aptamerinin
FL1-H histogramı ... 64
ġekil4-17 CHO-ICAM-1 ile inkübe edilmiĢ kontrol aptameri ve D-10-2 aptamerinin FL1-H histogramı ... 65
ġekil4-18 CHO-ICAM-1 ile inkübe edilmiĢ kontrol aptameri ve D-10-3 aptamerinin FL1-H histogramı ... 65
ġekil4-19 Seçilen aptamerlerin floresans yoğunlukları grafiği ... 66
ġekil4-20 BD-10-1 aptamerinin Kd grafiği... 67
ġekil4-21 D-10-2 aptamerinin Kd grafiği ... 68
xi
SĠMGELER VE KISALTMALAR DĠZĠNĠ
Kısaltmalar Açıklama
SELEX Systematic Evolution of Ligands by Exponential Enrichment (Üssel ZenginleĢme ile Ligandların Sistematik Evrimi)
ICAM-1 Intercellular adhesion molecule-1 (Hücre Ġçi Adezyon Molekülü-1) ssDNA single-stranded deoxiribo nucleic acid (Tek zincirli deoksiribo nükleik asit) dsDNA double-stranded deoxiribo nucleic acid
(çift zincirli deoksiribo nükleik asit) MUC-1 Musin 1
CA15-3 Cancer Antigen 15-3 IL ‐ 2R interlökin ‐ 2 reseptörü
IGFBP ‐ 3 insülin benzeri büyüme faktörü bağlanma proteini ‐ 3 EGFR Epidermal Büyüme Faktörü Reseptörü
MMP Matriks Metalloproteinaz uPA Plazminojen Aktivatörleri TIL Tümör infiltre edici lenfositler ECM Ekstra selüler matriks
ADC Antikor-ilaç konjugatları ApDC Aptamer-ilaç konjugatları CE-SELEX Kapiler elektroforez SELEX GO-SELEX Grafen oksit SELEX
1 1. GĠRĠġ
ICAM-1 (Intercellular Adhesion Molecule 1, hücreler arası adezyon molekülü-1) ya da diğer bilinen adıyla CD54 (Cluster of Differentiation 54) insanlarda Icam-1 geni tarafından kodlanan bir proteindir. ICAM-1, antikorlar ve T-hücre reseptör proteinlerini de içeren immünglobulin süperailesinin bir üyesidir ayrıcakeratinositler ve lökositler arasındaki etkileĢime aracılık eden bir hücre yüzeyi reseptörüdür. Endotel hücreleri ve lökositler tarafından normal koĢullarda bazal seviyelerde ifade edilir. Ancak TNF-α, IFN-γ, IL-1 ve LPS gibi enflamatuar sitokinler tarafından ekspresyonu arttırılır(Weiner ve ark. 2010).Göğüs kanserinde metastaz olan hastalardan alınan numunelerde ICAM-1 proteinin ekspresyonu yüksek düzeylerde görülmektedir. ICAM-1 tutunmaya bağlı hücre-hücre etkileĢimlerinde rol alır ve hücrelerin ekstraselüler matriks boyunca hareketini ve ilerlemesini kolaylaĢtırır. ICAM-1 bağıĢıklık sisteminde önemli bir role sahiptir ve aktive edilmiĢ lökositler, 1 vasıtasıyla endotele yapıĢırlar. ICAM-1çeĢitli kanser türlerinde tümör prognozunda ve metastazda rol alır; kolorektal, pankreas, akciğer ve göğüs kanseri gibi birçok kanser türünde yüksek miktarda ekspre edilir. Literatürde kanserin ilerlemesi ve ICAM-1 seviyesinin yükselmesi ile iliĢkili birçok araĢtırma bulunmaktadır; Usami ve ark. ICAM-1 ekspresyonunun kanserde önemli bir faktör olduğunu göstererek göğüs kanserinde saldırgan tümör fenotipi ile ICAM-1 ifadesinin iliĢkisi olduğunu bulmuĢtur. Maurer ve ark. koleraktal kanserde ICAM-1 ekspresyonunun arttığını belirtmiĢtir. Çoğu kanser türünde yüksek miktarda ifade edilen ICAM-1 iyi bir diagnoz ve prognoz belirteci olarak kullanılabilme kapasitesine sahiptir.
Aptamerler üç boyutlu yapı özellikleri sayesinde hedef moleküle yüksek bir afinite ve özgüllükle bağlanabilen oligonükleotitlerdir. Aptamerlerin hedef molekülleri küçük bir iyon olabildiği gibi protein gibi büyük bir molekül de olabilir. Aptamerleri seçmek ve çoğaltmak için SELEX (Systematic Evolution of Ligands by Exponential Enrichment (Üssel ZenginleĢtirmeyle Ligandların Sistemik Evrimi)) tekniği kullanılmaktadır. Bu teknik 1014-1015 molekül içeren bir sentetik oligonükleotit havuzundan hedefe bağlanabilen dizilerin zenginleĢtirerek seçilmesinden oluĢur (Tuerk ve Gold, 1990; Bock ve ark., 1992).
2
SELEX iĢlemi sırasında oligonükleotit havuzu hedef ile inkübe edilir ve bağlanmayan diziler yıkanarak uzaklaĢtırılır. Bağlanan diziler hedeften ayrılır ve uçlarındaki primer bağlanma bölgeleri kullanılarak polimeraz zincir reaksiyonu (PZR) ile çoğaltılırlar. ZenginleĢtirilen bu ikincil DNA havuzu ssDNA‟ya (tek zincirli DNA) çevrilir ve SELEX‟in ikinci turunda kullanılmak için hazır hale getirilir. Bu ayırma ve zenginleĢtirme tekniği hedefe göre 8-20 tur aralığında değiĢim göstermektedir. Hedefe karĢı pozitif bir seçilim uygulanırken amaca göre bağlanılması istenmeyen hedeflere karĢı negatif bir seçilim de uygulanabilmektedir. Böylece oligonükleotit havuz hedefe karĢı zenginleĢtirilirken istenmeyen moleküllere bağlanması engellenebilir. SELEX ile zenginleĢtirilen DNA havuzları hedefe belli bir afinite ile bağlandığında seçilim sona erdirilir ve dizileme iĢlemi yapılır. Dizileme sonucunda en çok rastlanan diziler aday aptamer olarak seçilir ve hedefle oluĢturdukları kompleksin çözüĢüm katsayıları (Kd)
hesaplanır. DüĢük çözüĢüm katsayısına sahip diziler aptamer olarak seçilir. Bu seçilen aptamerler ilerleyen çalıĢmalarda kullanılmak için hazırdır (Bayraç ve ark., 2011). Bu çalıĢmanın amacı da ICAM-1 aracılığıyla metastazı engellemek için, SELEX adı verilen in vitro seçme yöntemini kullanarak ICAM-1‟a yüksek etkinlik ve özgünlükte bağlanabilen aptamer dizileri elde etmektir.
3
2. KURAMSAL TEMELLER VE KAYNAK ARAġTIRMASI
2.1. Kanser
Kanser, bir hücre ya da hücre grubunun, organizmanın sahip olduğu kontrol mekanizmalarının etkisinden çıkıp kontrolsüz bir Ģekilde çoğalmasıdır(Fidler ve ark., 2001). Kanser çeĢitleri; köken aldığı hücrenin yapısı, ilerleme süreci, baĢka dokulara yaptığı metastazlar ve moleküler seviyedeki farklılıkları açısından kiĢiden kiĢiye değiĢiklik göstermektedir (Segal ve ark., 2001). Normal yaĢam koĢullarında hücreler belirli bir döngü içerisinde olup, ölen hücrelerin yerini yeni hücreler almakta ve bu döngügenlerin kontrolünde gerçekleĢmektedir (Ferlay ve ark., 2015). Genler hem hücrelerin bölünüp çoğalmasını sağlamakta hem de kontrolsüz hücre bölünmesini engellemektedir. Çevresel faktörler baĢta olmak üzere çeĢitli nedenlerle hücre DNA‟sında değiĢiklikler meydana gelmekte bu değiĢikliklerde normal döngüyü bozarak kanser geliĢimine neden olabilmektedir.Normal koĢullarda hücre döngüsü; DNA‟nın sentezlendiği S evresi, mitoz bölünmenin gerçekleĢtiği M evresi, duraklama evreleri olan G1 ve G2 evrelerinden oluĢmaktadır. Hücrelerin stabil halde olduğu G0 evresi ise, uyarı sonrasındadöngünün baĢlayacağı potansiyel evredir. Bu evrelerdemeydana gelen bir uyarıhücreler arasındaki döngüyü bozarak kontrolsüz bir hücre çoğalmasına neden olabilir (Li ve ark., 2014; Robertson ve ark., 1990).Kanser hücrelerinin ġekil 2-1‟de de görüldüğü gibi büyüme baskılayıcılardan kaçma, yeni damar oluĢumunun desteklenmesi,apoptozisi baskılama, anjiyogenez, genetik kararsızlıklar, mutasyonlar ve metastaz gibi normal hücreden farklı özellikleri vardır (Hanahan ve Weinberg, 2011). Kanser hücrelerini etkileyen genetik ve epigenetik modifikasyonlar, 6 önemli fenotipik özelliğin ortaya çıkmasına neden olur;
1-Kanser hücreleri mutasyonlar ve epigenetik değiĢimler sonucunda aktifleĢen proto-onkogenler oluĢtururlar. TGF-α gibi kendi kendilerine büyüme faktörlerisekresyonunu gerçekleĢtirebilirler ve böylece dıĢarıdan herhangi bir sinyale ihtiyaç duymadan, büyümeye devam edebilirler.
2-Kanser hücreleri anti-büyüme sinyallerine karĢı duyarsızlardır.
3-Kanser hücreleri apoptozdan kaçabilirler. Bu özelliğe genellikle p53 gibi tümör baskılayıcı genlerin sinyal yolaklarının mutasyonu sonucunda sahip olurlar.
4
4-Kanser hücrelerinde telomeraz aktivitesi oldukça yüksektir. 5-Artan oksijen ve besin ihtiyacından dolayı anjiyogenez baĢlar.
6-Primer tümör hücreleri bulundukları dokudan ayrılarak, farklı bir bölgeye göç edip sekonder tümör oluĢtururlar. Bu sürece metastaz denir (Fidler, 2001).
ġekil 2- 1Kanser hücresinin özellikleri (Hanahan ve Weinberg, 2011)
2.1.1. Kanser nedenleri
Kanserlerin %5‟i kalıtımsal, %90-95‟i vücut hücrelerinde sonradan oluĢan bozulmalardan kaynaklanmaktadır. Kanser insidansı farklı sebeplere bağlı olarak değiĢmektedir (Scully ve ark., 2012). Kansere neden olan etkenler için yaĢ, cinsiyet, genetik faktörler, beslenme alıĢkanlıkları, çalıĢma ortamları, hormonlar ve prekanseröz hastalıklar ve lezyonların varlığı gibi farklı sebepler gösterilebilir. Genetik hasarlar kalıtımsal olabileceği gibi, çevresel faktörlerden de kaynaklanabilir (Vogel ve ark., 2010). Kanser, son yıllarda insidansı ve mortalitesi artan bir hastalıktır ve birçok ülkede kardiyovasküler hastalıklardan sonra ikinci sırada yeralmaktadır.Çizelge 2-1‟de kansere neden olan risk faktörleri türlerine göre gösterilmiĢtir. Sigara ve alkol kullanımı, enfeksiyon, uzun süre güneĢ altında kalma, kimyasal maddeler, hava kirliliği,
5
radyasyon, kötü beslenme alıĢkanlığı ve genetik faktörler kansere neden olabilmektedir (Fidler ve ark., 2001).
Kanser türü Risk faktörü
Akciğer kanseri sigara, asbestos,hava kirliliği Göğüs kanseri radyasyon, genetik değiĢiklikler
Kalın bağırsak kanseri polipler, genetik değiĢiklikler, diyet, sigara, iltihaplı barsak hastalıkları
Prostat kanseri diyet
prostatta iyi huylu değiĢiklikler
Karaciğer kanseri karaciğer hepatit virüsleri (HBV,HCV) Pankreas kanseri pankreas sigara diyabet
Çizelge 2- 1 Kansere neden olan risk faktörleri
2.1.2. Kanser belirtileri
Kanser belirtisi olabilecek bazı genel belirtiler ve semptomlar Ģunlardır;
Yorgunluk
Nedensiz kilo kaybı
Ciltde meydana gelen ĢiĢlikler
Cilt renginin sararması, koyulaĢması ya da kızarması
ĠyileĢmeyen yaralar
Bağırsak veya mesane alıĢkanlıklarındaki değiĢiklikler
Sürekli öksürük hali ve nefes darlığı
Yutma güçlüğü
Enfeksiyon olmadığı halde görülen yüksek ateĢ
Ses kısıklığı
Kas veya eklem ağrısı(Ferlay ve ark., 2015). 2.1.4. Kanser tedavi yöntemleri
Kanser tedavisinde mortaliteyi azaltmak ve sağ kalımı artırmak için çeĢitli tedavi yöntemleri kullanılmaktadır. Bunlar: cerrahi iĢlemler, kemoterapi,radyoterapi,immunoterapi, gen tedavisi ve anjiyogenez inhibitörleri olarak
6
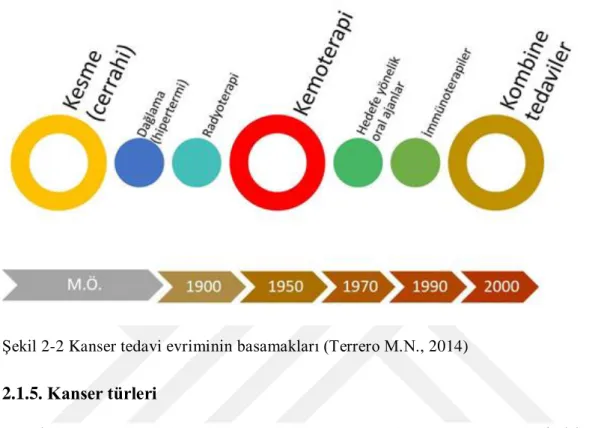
sayılabilir (Terrero M.N., 2014). Kanser tedavi evriminin basamakları ġekil 2-2‟de gösterilmiĢtir.
ġekil 2-2 Kanser tedavi evriminin basamakları (Terrero M.N., 2014)
2.1.5. Kanser türleri
Karsinom: En yaygın kanser türü karsinomdur (Strell ve ark., 2010). Epitel hücreleri tarafından oluĢturulurlar ve oluĢturuldukları epitel hücre türüne göre isimlendirilirler: Adenokarsinom,bazal hücreli karsinom, skuamöz hücreli karsinom ve transizyonel hücreli karsinom(Ferlay ve ark., 2015).
Sarkom: Sarkomlar kan ve lenf damarlarında, kaslarda ve yağlarda, kemik ve yumuĢak dokularda meydana gelen kanser türüdür (Siegel ve ark., 2014).
Lösemi: Kemik iliğinin kanı oluĢturan dokusunda meydana gelen kanser türüdür. Lösemide katı tümörler oluĢmamaktadır. Kanda ve kemik iliğinde anormal derecede beyaz kan hücreleri oluĢmaktadır. Normal kan hücrelerinin sayılarının azalması, dokulara oksijen taĢıması, kanamanın kontrol edilmesi ve enfeksiyonlarla mücadele edilmesi gibi hayati fonksiyonları zorlaĢtırmaktadır (Terrero M.N., 2014).
7
Lenfoma: Lenfoma, lenfositlerde köken alan bir kanser türüdür. Lenfoma kanserinde, lenf nodlarında, lenf damarlarında ve vücudun diğer organlarında lenfosit sayısında anormal derecede artıĢ meydana gelir (Siegel ve ark., 2014).
Multipl Myelom: Multipl myelom, plazma hücrelerinde görülür ve anormal derecede plazma hücrelesi kemik iliğinde oluĢmaktır. Myelom hücreleri olarak adlandırılan bu hücreler kemiklerde tümöre neden olmaktadır (Siegel ve ark., 2014).
Germ Hücreli Tümörler: Germ hücreli tümörler, sperm veya yumurta hücrelerinde görülmektedir. Bu tümörler, vücudun hemen hemen her yerinde oluĢabilmektedir(Fidler ve ark., 2001).
2.1.6.Kanser Ġstatistikleri
Dünya çapında kanser görülme sıklığı bölgelere göre değiĢmektedir. Dünya da her yıl 13 milyon, Türkiye‟de ise her yıl yeni kanser vakası 150 bindir. Dünya sağlık örgütünün (WHO) 2018 verilerine görekanser 18,1 milyon yeni vakayave 9,6 milyon kiĢinin ölümüne neden olmuĢtur ayrıca2020 yılında tüm dünyada kanserli vaka sayısının %30 artarak yaklaĢık 17 milyona ulaĢacağı tahmin edilmektedir. En yaygın kanser türleri; akciğer (2,09 milyon), göğüs (2,09 milyon) kolorektal (1,80 milyon), prostat (1,28 milyon), deri (1,04 milyon) ve mide (1,03 milyon) kanseri olarak belirlenmiĢtir.
2.2. Metastaz
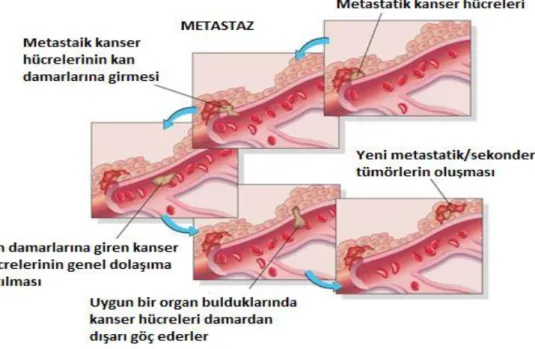
Metastaz, kanser hücrelerinin köken aldıkları primer tümör bölgesinden kan damarları aracılığıyla farklı doku veya organlara ulaĢması ve sekonder kanser geliĢimine neden olması durumudur (Ferlay ve ark., 2015). Metastaz, kanser hücrelerinin primer tümörden çevre dokulara ve uzak organlara yayılmasını tanımlamak için kullanılan genel bir terimdir ve kanser morbidite ve mortalitesinin baĢlıca nedenidir (Witte ve ark., 1998). Kanser ölümlerinin yaklaĢık %90'ından metastazın sorumlu olduğu tahmin edilmektedir (Garvin ve Dabrosin, 2003). Metastaz, birçok basamaktan oluĢmaktadır;
Anjiyogenez,
Metastatik hücrelerin primer tümörden ayrılması,
Ġnvazyon,
8
Kan damarları aracılığıyla metastatik hücrelerin hedef organa ulaĢması,
Ekstravazasyon.
Tümör hücrelerinin çeĢitli bölgelerinde mutasyonlar veya kromozomda değiĢimler meydana gelir. Bu değiĢimler onkogenleri, tümör baskılayıcı genleri ve proteaz inhibitörleri gibi metastazla iliĢkili genleri etkileyebilir (Weigelt, 2005).
ġekil 2-3 Metastazın aĢamaları (Weigelt, 2005)
Metastaz; tümör hücrelerinin köken aldıkları primer dokudan farklı bir dokuya göç etmesi ve yeni dokuda sekonder tümörlerin oluĢma sürecidir. Tümör anjiogenezitümör hücresinin büyümesi için gerekli olan besin ve enerji ihtiyacını sağlamakta ve aynı zamanda dolaĢım sistemi ile etkileĢimini arttırmaktadır(Garvin ve Dabrosin, 2003). Metastaz hücrelerarası iletiĢim kaybınada sebep olmaktadır.Tümör hücrelerinde hücrelerarası adhezyon moleküllerinin sayısında (kaderinler, integrinler, selektinler, ICAM) farklılıklar görülmektedir.
Tutunma özelliklerinikaybeden metastatik tümör hücreleri lenf ve kan damarları yoluyla diğer dokulara yayılmaktadır. Lenf kanallarındaki tümör hücreleri bölgesel lenf nodlarına ulaĢır.Bölgesel lenf nodlarına tutunur ve burada çoğalmaya baĢlar. Bölgesel lenf nodunda kitümör hücreleri büyüdükten veya çoğaldıktan sonra tümör hücreleri baĢka lenf nodlarınada yayılma gösterebilir. Venöz duvarları tümör hücrelerinin
9
invazyonuna karĢı arter duvarlarından daha duyarlıdır (Chen ve ark., 2010). Ayrıca lenf dolaĢımının venöz dolaĢıma katılmasından dolayımetastaz süreci venöz yolla gerçekleĢmektedir. Metastatik tümör hücreleri introvazasyon sırasında bağıĢıklık sisteminden kendilerini korumak için lökosit ve trombosit pıhtıları ile anormal antijenlerini saklamaktadırlar (Welch ve ark., 2000).
Bir hastalığın moleküler özelliklerini anlamak, teĢhis, tedavi ve hastalığın doğasını ve mekanizmasını aydınlatmak için temeldir. Artan kamu bilinci ve istatistiksel veriler nedeniyle, kanser moleküler karakterizasyon için önemli bir hedeftir. BaĢarılı kanser tedavisi, her ikisi de uygun moleküler problar ve biyobelirteçler gerektiren erken tanı ve doğru evrelemeye dayanmaktadır. Metastaz taramasında kullanılan testler;
Karaciğer metastazında karın ultrasonografisi ve karaciğer kan testleri, Akciğer metastazında akciğer filmi,
Kemik metastazında kemik sintigrafisi , Beyin metastazında beyin MR‟ı ve
Tüm vücut taraması için PET-CT çekilmektedir .
Kanserle iliĢkili gen mutasyonları ve değiĢiklikler genellikle proteomik seviyelerde değiĢikliklere neden olur ve bu değiĢikliklerin bazıları hücre yüzeyinde meydana gelir. Hücre yüzey proteinleri hücrelerin ilk eriĢilebilir bölümleri olduğundan, nispeten kolaylıkla tespit edilebilir. Bu nedenle, hücre yüzeyi biyobelirteçleri kanserin aĢamasının erken ve kolay saptanması, farklılaĢması ve belirlenmesi için uygundur. Ayrıca, hem kanseri tespit etmek hem de tedaviye yanıtı izlemek için in vivo görüntüleme teknikleriyle birlikte kullanılabilirler.
Tümör belirteçleri, kanser tanısı için ve tedavi sonrası nüks tespiti için kullanılırlar. Tümör belirtecinin seviyesinin artması, kanser hücrelerinin varlığını göstermekle birlikte tek baĢına yeterli değildir. Bu sebeple tümör belirteçleri diğer tetkik ve tanı yöntemleriyle birlikte kullanılır (Lee ve ark., 2015). Prognostik belirteç gereksinimlerini karĢılamak için potansiyel belirteç, uzun bir takip süresi olan geniĢ hasta kohortlarında geriye dönük olarak test edilmelidir. Sonuç olarak, bulgular bağımsız bir araĢtırmacı grup tarafından doğrulanmalı test edilen belirtecin prognostik önemini doğrulamalıdır (Granato ve ark., 2006)
10 2.2.1. Metastaz belirteçleri
Metastaz belirteçleri, tümör hücrelerinin köken aldıkları bölgeden ayrılıp vücuda, dokulara, hücrelere yayılma olup olmadığını saptamak için önemli bir teĢhis yöntemidir. Metastaz belirteçleri tümör büyüklüğü ve kötü prognozla iliĢkilidir ayrıca tek olarak kullanıldığı gibi diğer belirteçlerle beraberde kullanılabilmektedir (Tirkkonen ve ark., 1997). En yaygın kullanılan metastaz belirteçleri arasında MCAM, CA 15-3, ErbB2-HER2/neu ve MUC-1 sayılabilir.
2.2.1.1 MCAM
CD146 veya MUC18 olarak da bilinen immünoglobulin süper ailesinin bir üyesi olan melanom hücre adezyon molekülü (MCAM) çeĢitli tümörlerde anormal Ģekilde ifade edilen ve tümör metastazı ile yakından iliĢkili bir hücre adezyon molekülüdür (Valentin ve ark., 2012).MCAM ekspresyonunun diğer yumurtalık epitelyal dokularının diğer patolojik tipleriyle karĢılaĢtırıldığında metastatik yumurtalık kanserlerinde daha yüksek olduğu gösterilmiĢtir (Rakha ve ark., 2008). MCAM sadece yumurtalık kanseri prognozunu belirlemek için moleküler bir iĢaretleyici olarak değil, metastatik yumurtalık kanserinde terapötik bir hedef olarak da kullanılmaktadır. MCAM, melanom, prostat kanseri, hepatoselüler karsinom ve ovaryum kanseri gibi birçok kanser türünde kötü prognoz ile iliĢkilendirilmiĢtir. Ayrıca, MCAM ekspresyonu yüksek dereceli tümörler, östrojen reseptörü (ER) negatif ve progesteron reseptörü (PR) negatif tümörleri ve üçlü negatif alt tip (ER- / PR- / insan epidermal büyüme faktörü reseptörü 2 (HER2) ile iliĢkilendirilmiĢtir (Wu ve ark., 2012).
2.2.1.2. CA 15-3
CA 15-3 (kanser antijeni 15-3), hücrelerin birbirine tutunması, bağıĢıklık ve metastazdan sorumlu bir proteindir. CA 15-3muc-1geninin ürünüdür. Normal göğüs dokusuna göre, kanserli göğüs dokusunda MUC-1 daha yüksek konsantrasyonlarda bulunur (Annecke ve ark., 2008). Bu sebeple göğüs kanserinde en çok kullanılan serum belirtecidir (Hwa ve ark., 2008). Serumdaki konsantrasyonu >30 U/mL ise yüksek olarak değerlendirilir. CA 15-3 seviyesin tümör kütle büyüklüğü ile iliĢkilidir dolayısıyla, CA 15-3 seviyesinin göğüs kanseri olan hastalarda yüksek bulunması çoğunlukla hastalığın kötü gidiĢatının bir göstergesidir. CA15-3, bölgesel nükslerin ve
11
uzak metastazların saptanmasında kullanılmaktadır. Bölgesel nüks tespit edilen göğüs kanseri olan hastaların CA 15-3 seviyesi %7, diğer organlara metastaz yapan göğüs kanserli hastaların ise CA 15-3 seviyesi %95-96 daha yüksek görülmektedir (Granato, 2006).
2.2.1.3. ErbB2-HER2/neu
HER2 olarak da bilinen erbb2 geni 17q21 kromozomunda lokalize olan bir onkoproteindir. Normal hücrelerde düĢük düzeyde ifade edilir. ErbB2, büyümeyi hızlandıran bir proto-onkogendir ve kanser hücrelerinin yaklaĢık% 20'sinde aĢırı miktarda ifade edilir (Eletxigerra ve ark., 2015). HER2-pozitif kanserler daha hızlı büyür, daha agresiftir ve kötü prognoz ile iliĢkilidir (Slamon ve ark., 1999). Bu haliyle, HER2'nin hedefli inhibisyonu, göğüs, yumurtalık, mide, mesane ve akciğer karsinoması dahil olmak üzere birçok kanserinin tedavisi için en çok tercih edilen metastaz belirteçlerinden biridir (Thiel ve ark., 2012; Stuhlmiller ve ark., 2015).
2.2.1.4. MUC-1
MUC-1 musin ailesine ait bir glikoproteindir. Musin ailesi proteinleri epitel yüzeyi korur ve kaygan hale getirir. MUC-1 göğüs, yumurtalık, pankreas, prostat, gastrik, kolorektal gibi kanser türünde yüksek miktarda ifade edilmektedir. Son zamanlarda metastatik kanser belirteci olarak kullanılmaya baĢlanmıĢtır. MUC-1 ifadesi ne kadar yüksek ise kanser o kadar ölümcül olmaktadır (Kasimanickam ve ark., 2014).Kullanılan bu belirteçlerin yanısıra normal hücrelerde bazal seviyede ifade edilen ICAM-1‟ın, kanser hücrelerinde seviyesi artmakta ve kötü prognozla iliĢkilendirilmektedir.Bu sebeple ICAM-1‟ın yeni bir metastaz belirteci olarak kullanılabileceği düĢünülmektedir(Granato ve ark., 2006).
2.3. ICAM-1
ICAM-1 (Intercellular Adhesion Molecule 1, Hücreler arası adezyon molekülü-1) ya da diğer bilinen adıyla CD54 (Cluster of Differentiation 54) endotelyal ve epitel hücreleri, vasküler düz kas hücreleri ve bazı lökosit alt kümeleri gibi çok sayıda hücre tipinin hücre yüzeyinde ifade edilen, insanlarda icam-1geni tarafından kodlanan ve immünoglobulin süper ailesine ait bir transmembran glikoproteindir. ICAM-1,
12
lökositlerin ve endotel hücrelerinin zarlarında düĢük konsantrasyonlarda sürekli olarak bulunan hücreler arası yapıĢma molekülüdür (Schröder ve ark., 2011). ICAM-1, hücre yüzeyi üzerinde; vasküler endotel hücreleri, timik ve mukozal epitel hücreleri, dermal fibroblastlar ve düĢük derecede periferik kan lökositleri üzerinde bulunur ve enfeksiyon veya yaralanma bölgelerindeki lökositlerin endotel hücrelerine yapıĢmasını arttırır(Eletxigerra ve ark., 2015). ICAM-1, soğuk algınlığı insan rinovirüsleri (HRV'ler), coxsackievirus A21 ve malarial parazit Plasmodium falciparum gibi çeĢitli patojenler tarafından da kullanılır. Endotel hücrelerde ICAM-1 ekspresyonu, tümör nekroz faktörü (TNF) -a, interlökin (IL) -1β ve interferon (IFN) -γ gibi enflamatuar sitokinlerle artabilir (O'Hanlon ve ark., 2002). ICAM-1, CD11a/CD18 veya CD11b/CD18 gibi integrinlere bağlanır. Enflamasyon bölgelerinde, aktive edilmiĢ lökositler önce endotel hücrelerinde açığa çıkan selektinler ile zayıf bir Ģekilde etkileĢime girer, daha sonra ICAM-1 yoluyla endoteliye sıkı sıkıya yapıĢır, ardından hücrelerin damar duvarı içinden geçiĢi sağlanır(Veitonmäki ve ark., 2013). Endotelyal ICAM-1'in önemli bir rol oynayabileceği metastaz sırasında dolaĢımdaki tümör hücrelerinin ekstravazasyonu için benzer bir mekanizma önerilmiĢtir (Roland ve ark., 2007).
ICAM-1‟in hayati rol oynadığı immünolojik olaylar 3 farklı durum; bağlantı oluĢumu, yeniden düzenleme ve kararlı immünolojik komplekslerin oluĢumuile karakterizedir. Ġlk adımdaki bağlantı oluĢumu esnasında, antijen sunan hücreler ve T hücreleri arasındaki yakın temas; ICAM-1 ve LFA-1‟in yardımları ile sağlanır. ICAM-1‟in ligandına bağlanması sırasında T reseptörleri için yüzeyde bulunan CD45-CD43 glikoproteinlerinden kaynaklanan sterik engel aĢılarak; Antigen Sunan Hücreler (APC)‟ler yüzeyindeki MHC-peptidleri ile etkileĢim sağlanır(Eletxigerra ve ark., 2015). TCR ve MHC-peptid kompleksleri arasındaki düĢük affinite olayı ve iki hücre arasındaki etkileĢim bu Ģekilde kalabilir. Eğer TCR ve MHC-peptid kompleksibirbirlerine yüksek affinite gösterirlerse, sinyal yolağı tetiklenir, bu yolağın tetiklenmesi ilgili moleküllerin yeniden düzenlenmesine yol açar. TCR ve MHC- peptid kompleksleri içeriye, merkeze doğru hareketlenirken dıĢarıya doğru LFA-1/ICAM-1 moleküllerince etraflarında sıkı bir halka oluĢturulur. Bu basamak ile kararlı immünolojik kompleksler oluĢturulmuĢ olur(Veitonmäki ve ark., 2013). Yeniden düzenleme ve halka oluĢumu sürecinde ICAM-1 polimerleĢerek LFA-1‟e daha yüksek
13
afinite ile bağlanma gösterir. Buna ek olarak halka etrafına toplanmıĢ TCR-MHC peptid kompleksleri hücreler ve sinyal yolakları ile optimal iletiĢimi sağlar. ICAM-1‟in ayrıca MHC-1 ve MHC-2‟ye seçili antijen sunumu esnasında yardımcı uyaran olarak görev aldığı bildirilmiĢtir (Schröder ve ark., 2011).
Hücresel adezyon molekülleri, tümör ilerlemesi ve metastazında rol oynar. DeğiĢik hücrelerde yapısal olarak bulunmakla birlikte, iltihabi dokularda IFNγ, IL-1 ve TNFα gibi para-inflamatuvar sitokinlerin uyarısıyla hücre yüzeyinde yoğunluğu artmaktadır (Roland ve ark., 2007). Uyarı sonrası, ICAM-1'in hücre yüzeyinde belirmesi, 2 ile 4 saat arasında baĢlar ve 12-16 saat arasında plato çizer. Bu durum ortamda sitokin bulunması durumundaise, 24 ile 72 saate kadar devam eder. ICAM-1 molekülleri eozinofillerin, T lenfositlerin ve nötrofillerin göçünde rol alırlar (Rosette ve ark., 2005). Ayrıca ICAM-1, β2 integrinlerden, LFA-1 ve Mac-1'in karĢı hücrede bağlandığı liganddır. Bu moleküller monoklonal antikorlarla bloke edilecek olursa, bu hücrelerin endotele yapıĢamadığı ve inflamasyon bölgesinde birikemediği görülmüĢtür (Schröder ve ark., 2011).ICAM-1, spesifik olarak lökosit efektör fonksiyonlarında, antijen sunan hücrelerin antijen sunumu sırasında T lenfositlere yapıĢmasında, mikrobiyal patojenezde ve sinyal iletim olaylarında görev almaktadır. CD11a/CD18 veya CD11b/CD18 tipindeki integrinlere bağlanır ve ayrıca Rhinovirus tarafından da reseptör olarakkullanılır (Fakhari ve ark., 2011).ICAM-1, hücre türüne bağlı olarak, antijen sunumu sırasındaki hücre hücre etkileĢimlerinde ve dıĢardan gelen sinyal iletimlerinde rol almaktadır. ICAM-1, hücre zarı protein ekspresyonunda artıĢa, reaktif oksijen türü üretimine ve hücre çoğalmasına neden olan fosforilasyon yoluyla spesifik kinazları aktive etmek için kullanılmaktır (Heer ve ark., 2006).ICAM-1 fibroblastlar, lökositler, keratinositler, endotelyal ve epitelyal hücrelerde düĢük bir bazal seviyesinde ifade edilir, fakat çeĢitli inflamatuar mediatörlere yanıt olarak yukarı regüle edilir (Schröder ve ark., 2011; Veitonmäki ve ark., 2013).
2.3.1. ICAM-1’ın Yapısı
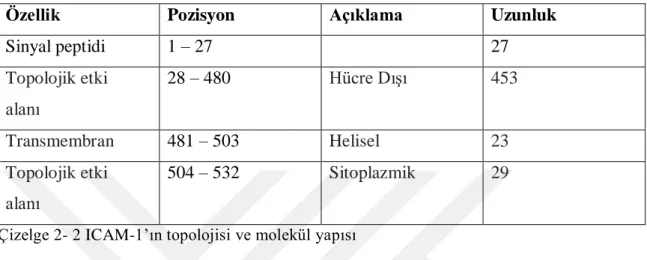
ICAM-1, 532 amino asit içeren tek bir polipeptit zinciridir. ICAM-1, beĢ immünoglobulin (Ig) benzeri alan içeren büyük bir hücre dıĢı bölgeden, bir hidrofobik transmembran bölgeden ve kısa bir sitoplazmik kuyruktan oluĢmaktadır (Roland ve ark., 2007; Veitonmäki ve ark., 2013). Çizelge 2-2 veġekil 2-4‟te ICAM-1‟ın topolojisi
14
ve molekül yapısı gösterilmiĢtir. Farklı glikosilasyon modellerinden dolayı, moleküler ağırlığı 80 ile 114 kDa arasında değiĢir. LFA-1 ve Mac1 integrinleri, fibrinojen, Muc1 dahil olmak üzere çeĢitli ICAM-1 ligandları tanımlanmıĢtır (Rahn ve ark., 2005; Mace ve ark., 2010).
Özellik Pozisyon Açıklama Uzunluk
Sinyal peptidi 1 – 27 27 Topolojik etki alanı 28 – 480 Hücre DıĢı 453 Transmembran 481 – 503 Helisel 23 Topolojik etki alanı 504 – 532 Sitoplazmik 29
Çizelge 2- 2 ICAM-1‟ın topolojisi ve molekül yapısı
ICAM-1 hücre yapıĢması, migrasyon ve agregasyon gibi çok sayıda hücresel yanıtla iliĢkili önemli sinyalleme özelliklerine sahip bir adhezyon molekülüdür. ICAM-1, endotelyal ve epitelyal hücreler, düz kas hücreleri, perisitler, fibroblastlar ve keratinositler dâhil olmak üzere çok çeĢitli hücrelerin yüzeyinde yapısal olarak ifade edilir. Genel olarak, ICAM'ler hücre zarını kapsar ve sadece kısa bir sitoplazmik kuyruk içerirler (Klausz ve ark., 2014; Ghislin, 2012).
15 ġekil 2-4 ICAM-1 molekül yapısı (Ghislin, 2012)
2.3.2. ICAM-1’ın Kanserdeki Rolü
Adezyon molekülleri, hayatta kalma, göç, ekstravazasyon, ve metastaz gibi kansere bağlı biyolojik süreçleri mümkün kılar, böylece kanserin ilerlemesinde önemli bir rol oynar. ICAM-1 birçok kanser türünde ifade edilir ve kolorektal, renal hücreli karsinom ve pankreas dahil olmak üzere çeĢitli kanserlerde tümör prognozunda önemli bir rol oynar (Veitonmäki ve ark., 2013). ICAM'lerin fonksiyonundaki ve ekspresyonundaki değiĢiklikler normal hücre-hücre etkileĢimlerinin bozulmasına neden olur ve malign transformasyona ve tümör ilerlemesine neden olabilir(Veitonmäki ve ark., 2013). ICAM-1, endotel hücrelerinde ve bazı lenfositler ve monositler üzerinde yapısal olarak düĢük seviyelerde ifade edilmesine rağmen, sitokinlerin ve reaktif oksijen türlerinin varlığında ekspresyonu önemli ölçüde artmaktadır (Ghislin, 2012). ICAM-1 çeĢitli kanser hücrelerinde ifade edilir ve ifade edilen ICAM-1‟in göğüs, akciğer gibi kanser türleri için potansiyel bir biyobelirteç olduğu düĢünülmektedir. Klasik bir kanser biyobelirteci olmaktan daha önemlisi ICAM-1 seviyeleri, hastalığın evresi, saldırganlığı, ilerlemesi ve metastatik potansiyeli hakkında bilgi verebilir(Roland ve ark., 2007).ICAM-1 bir yapıĢma molekülüdür aynı zamanda bir metastatik fenotipi artırabilen bir sinyal yanıtını da ortaya çıkarabilir.ICAM-1 proteini, bir yandan metastazı kolaylaĢtıran önemli bir adezyon molekülüdür diğer yandan, T hücresi aracılı eliminasyon için gerekli olan immünolojik sinapsı desteklemektedir. (Schröder ve ark.,
16
2011). ICAM-1'ın inhibisyonunun, akciğer kanseri hücrelerinin in vitro invazyonunu bloke ettiği gösterilmiĢtir. Antisens oligonükleotitleri kullanarak melanoma hücrelerinde ICAM-1 inhibisyonu raporunda, akciğer metastazlarında azalma gözlenmiĢtir (Guo ve ark., 2014). Çoklu korelatif çalıĢmalar göğüs kanseri hastalarında çözünebilir serum ICAM-1 fraksiyonunda bir artıĢ olduğunu göstermiĢtir, ancak bu çözünebilir formun endotelyal veya epitelyal hücrelerden mi kaynaklandığı açık değildir. Bu nedenle, göğüs kanseri ilerlemesinde ICAM-1 tutulumu için doğrudan bir kanıt henüz mevcut değildir (Veitonmäki ve ark., 2013).
Tümör invazyonu ve tümör hücresi yayılması, hücre dıĢı matris ile tümör hücrelerinin yapıĢkan temaslarının oluĢumu ve gevĢemesi arasında sıkı bir Ģekilde düzenlenmiĢ etkileĢimi içerir(Ghislin, 2012). ICAM-1'ın matriks yoluyla invazyona katkıda bulunduğu mekanizma, hücre içi sinyal yollarına bağlanabilir. Müsin 1'in (MUC-1) bilinen tek doğrudan ligandı olan ICAM-1'ın, MUC-1'ın inoksanozile biçimini ifade eden göğüs kanseri hücrelerinde kalsiyum aracılı bir sinyal baĢlatabildiği gösterilmiĢtir. ICAM-1 ekspresyonunun göğüs kanserinde daha agresif tümör fenotipi ile iliĢkili olduğu bulunmuĢtur (Schröder C., 2011).MUC-1, göğüs tümörü oluĢumu ve metastatik yayılmanın yerleĢik bir belirtecidir (Kasimanickam ve ark., 2014).ICAM-1 pozitif tümörlü hastalarda lenf nodu veya karaciğer metastazı insidansı, ICAM-1-negatif tümörleri olanlara göre anlamlı olarak daha yüksek gözlenmiĢtir. Tümör infiltre edici lenfositlerin (TIL) infiltrasyonu, ICAM-1 pozitif tümörlerde, ICAM-1 negatif tümörlerden daha sık görülmüĢtür. ICAM-1 negatif tümörleri olan hastaların prognozu, ICAM-1 pozitif tümörleri olanlardan anlamlı olarak daha zayıftır. Sonuç olarak, bu bulgular ICAM-1 ekspresyonunun metastaz ile yakından iliĢkili olduğunu ve göğüs kanseri olan hastalarda prognozun yararlı bir göstergesi olabileceğini düĢündürmektedir(Schröder ve ark., 2011).Göğüs kanseri dokularında ICAM-1 proteinin ifadelenmesinin arttığı bildirilmiĢ olup, metastaz olan hastalardan alınan numunelerde en yüksek düzeydedir (Klausz ve ark., 2014). Epitelyal tümör oluĢumunun yanı sıra bir dizi enflamatuar ve immün yanıtla iliĢkilidir. Yayınlanan kanıtlar, ICAM-1 ekspresyonunun upregülasyonunun veya inhibisyonunun, özellikle göğüs kanseri baĢta olmak üzere, insan tümör progresyonuna katkıda bulunup bulunmadığı konusunda tartıĢmalıdır. Bazı kolorektal ve göğüs kanseri dokularının immünohistokimyasal analizi, ICAM-1'ın düĢük ekspresyon seviyelerini göstermektedir. Bu, ICAM-1'ın bir
17
immüno-gözetim mekanizması aracılığıyla tümör baskılamasında rol oynayabileceği ihtimaline yol açmaktadır. Diğer taraftan, bu proteinin artmıĢ ekspresyonu, mide, pankreas ve göğüs kanseri dokularında bildirilmiĢtir. Her ne kadar birçok kanser türünde yüksek oranda ifade edilse de, ICAM-1'ı prognoz belirteci olarak kullanmak, teĢhis belirteci olarak kullanılmasından daha avantajlıdır (Schröder ve ark., 2011). 2.3.3. ICAM-1 Ġfadesine Etki Eden Sinyal Yolakları
ICAM-1 hücre-hücre ve hücre-ECM (Eksra selüler matriks) etkiliĢiminde önemli rol oynar(Roland ve ark., 2007). ICAM-1 MUC-1, LFA-1 (CD11a/CD18) ve Mac-1 (CD11b/CD18)‟e bağlanarak immünositlerin saldırısına karĢı kanser hücrelerini korumaktadır (Ghislin, 2012).
2.3.3.1. MUC-1
MUC-1 musin ailesine ait bir glikoproteindir. Musin ailesi proteinleri epitel yüzeyi korur ve kaygan hale getirir. MUC-1,göğüs, yumurtalık, pankreas, prostat, gastrik, kolorektal kanser gibi kanser türlerinde fazla ifade edilmektedir (O'Hanlon ve ark., 2002).Son zamanlarda metastatik kanserlerin belirteçleri olarak kullanılmaya baĢlanmıĢtır. MUC-1 ifadasi ne kadar yüksekse kanser o kadar ölümcül olmaktadır. Kanserin adaptasyonu ve MUC-1‟in değiĢimi metastatik ilerlemede önemlidir (Kasimanickam ve ark., 2014). Kanser hücrelerinin kan damarlarının duvarlarına tutunması için selektin-glikoprotein etkileĢimine ihtiyaç vardır. MUC-1 bir E-selektin ve ICAM-1 ligandıdır. Göğüs kanserinin metastatik yolağında önemli bir bağlantıdır. MUC-1‟in az glikozillenmiĢ halinin selektin bağlayıcı kısımlara sahip olduğu bulunmuĢ. Literatürde MUC-1‟in göğüs kanseri (ZR-75-1 ve MCF7) hücrelerinde ifade edildiği gösterilmiĢtir. Yüksek metastatik ZR-75-1 hücreleri az glikozillenmiĢ MUC-1 formunu daha fazla üretirken, az metastatik MCF 7 hücreleri bu proteini daha az ürettiği belirtilmiĢtir. MetastatikZR-75-1 hücrelerinde E-selektin etkileĢimi daha fazladır. ġekil 2-5‟te gösterildiği gibi sağlam bir tutunma olmaktadır. Bu da MUC-1:ICAM-1 etkileĢimine hücrelerin yüzeye yapıĢmasını sağlamıĢtır (Ghislin ve ark., 2012). MUC-1 tüm kanser hücrelerinde ifade edilir ve normaldeki glikozillenme seviyesinin altında glikozillenir. Ekstraselüler bölgenin çekirdek yapısı serin ve threonin açısından zengin olan 20 özgün aminoasit dizisinin 25-150 tekrarlı biriminden oluĢmaktadır. Bu da bu
18
proteini diğer zar proteinlerinden 5-10 kat daha büyük hale getirmektedir. Normalde bu aminoasitler fazla miktarda O-glikozillenmiĢ olurlar. Ancak kanser hücrelerinde bulunan mutant MUC-1‟lerde daha kısa oligosakkaritler üretmektedirler. Bu da MUC-1 proteinin E-selektin‟e daha verimli bağlanmasını sağlamaktadır (O'Hanlon ve ark., 2002). MUC-1‟in kısa glikozillenmesi diğer taraftan da bu proteinin merkez kısmının daha ulaĢılabilir olmasına neden olmuĢtur ve MUC-1 ifadesini tanımak için geliĢtirilen probların bağlanmasına kolaylık sağlamıĢtır. Böylece ICAM-1, MUC-1 proteinin kısa glikozillenmiĢ mutant türünün çekirdek/merkez kısmına daha kolay bağlanabilmektedir. Bu nedenlede, CTC (kanda dolaĢan tümör hücreleri, circulating tumor cells) hücreleri kan damarlarının yüzeyine daha kolay yuvarlanıp tutunabilmek için kısa glikozillenmiĢ MUC-1 ifade etmektedirler(Ghislin ve ark., 2012). CTC hücrelerinin damarda var olan hidrodinamik kayma gerilimine (hydrodynamic shear stres) rağmen damar duvarına tutunmaları hücre yüzeylerinde ifade ettikleri selektin ailesi tutunma molekülleri sayesinde olur. Bu durumun benzerini inflamasyon sırasında lökositler (beyaz kan hücreleri) de yapmaktadır. Ancak beyaz kan hücrelerinde MUC-1 proteini yoktur ve ICAM-1 ligandı olarak lökosit beta-2 integrin (LFA-1, Mac-1) ifade etmektedirler (Kasimanickam ve ark., 2014).
ġekil 2- 5 Metastazda MUC-1 ve ICAM-1 ĠliĢkisi (Kasimanickam ve ark., 2014)
ICAM-1, lökosit transendotelyal göçü ve antijen sunumunda rol alır. ġekil 2-5‟de de görüldüğü gibi transendotelyal göç dört kısımda gerçekleĢir. Ġlk transendotelyal göç adımı, selektin bağlanması ile baĢlar. Selektinler, diğer hücrenin sialik asit içeren
19
karbohidratı ile etkileĢime girer. E ve P selektinler, endotel hücrelerinde bulunurlar, lökositlerin ve P-selektin glikoprotein ligand-1 üzerindeki sialy Lewis x ile iliĢkili karbohidratlara bağlıdırlar. L selektin ise dolaĢımdaki lökositlerin üzerinde olup, endotel hücrelerindeki sialy Lewis x ile bağlıdırlar. Lökositlerin endotel hücrelerine bağlanmasıyla, hücreler yavaĢlar ve yuvarlanmaya baĢlar. Kemokinler, ikinci transendotelyal göç basamağında görev alırlar. Lökosit yüzeyinde aktive olan integrinler, böylelikle inflamasyonlu endotel alanında bulunan adezyon moleküllerine sıkıca bağlanabilirler. Bu bağlanma endotelin tek katlı hücreleri içerisine transendotelyal göç fırsatı olan alanları bulabilecek Ģekilde yayılmasına ve yavaĢça göç etmesine imkân verir. Lökositler, endotele sıkıca tutunduklarında mikrovili benzeri kıvrımlar (podozomlar) oluĢtururlar bu da endotel hücre yüzeyinin derinine inmeye imkân sağlar.
2.3.3.2 LFA-1
LFA-1 birçok hücresel süreçte önemli roller oynadığından, fonksiyonlarının bozukluğu ciddi otoimmün ve enflamatuar hastalıklara neden olabilir ve miyelom, malign lenfoma ve akut ve kronik lösemiler dahil olmak üzere birçok kanserde rol oynar(Fakhari ve ark., 2011). Böylece LFA-1 çoklu otoimmün ve enflamatuar hastalıkların ve kanserin tedavisi için terapötik bir hedef haline gelmiĢtir. LFA-1, T hücreleri tarafından üretilir, ICAM-1 için bir reseptör görevi görür. ġekil 2-6LFA-1 ve ICAM-1 iliĢkisini göstermektedir. LFA-1‟in iki zinciri vardır; CD11 ve CD18. Beyaz kan hücrelerinin damara tutunması ve transendotel göçünde görevlidir (Kasimanickam ve ark., 2014).Transendotel göç sırasında lökositlerin endotel katmandan dıĢarı çıkabilmesi için endotel hücreler arasındaki bağlantı molekülleri JAM-A‟ların arasındaki homofilik etkileĢim LFA-1 tarafından bozulur. ICAM-1, LFA-1 (CD11a/CD18) ve Mac-1‟e bağlanarak immünositlerin saldırısına karĢı kanser hücrelerini korur böylelikle metastazı kolaylaĢtırır (Ghislin ve ark., 2012).
20
ġekil 2- 6 Metastazda LFA-1 ve ICAM-1 ĠliĢkisi (Ghislin ve ark., 2012)
2.4. Aptamer
Ġlk olarak 1990 yılında (Ellington ve Szostak, 1990 ; Tuerk ve Gold, 1990) tanıtılan aptamerler , metal iyonları, kimyasal bileĢikler , proteinler, hücreler gibi geniĢ bir hedef yelpazesine yüksek bir etkinlik ve özgünlük ile bağlanabilen tek zincirli kısa oligonükleotit dizileridir (Wang ve ark., 2018). Aptamer kelimesi Latince kökenli olup “uymak” anlamına gelen “aptus” ve “bir bütünün parçası” anlamına gelen “meros” kelimelerinden üretilmiĢtir (Jayasena, 1999; Darmostuk ve ark., 2015). Gen ekspresyonunu düzenleyen riboswitchler 2004 yılında keĢfedilen doğal aptamerlerdir. Riboswitchlerin keĢfiyle aptamer alanındaki araĢtırmalar artıĢ göstermiĢtir (Mandal ve Breaker, 2004). Aptamerlerin molekül ağırlıkları antikorların molekül ağırlığına göre çok daha küçüktür. Aptamerlerin molekül ağırlıkları yaklaĢık 6 ile 40 kDa, antikorların molekül ağırlıkları ise yaklaĢık 150 kDa‟dur.Aptamerler; hedef moleküllere yüksek özgünlük ve afinite ile bağlanabilen üç boyutlu konformasyonel yapıya sahiptirler. Üç boyutlu bu yapı tRNA‟nın globüler Ģekline benzerdir. Aptamerlerin kendilerine özgü üç boyutlu yapıları vardır (Proske ve ark. 2000). Üç boyutlu yapılarına dayanarak, aptamerler tekli moleküllerden çok çeĢitli hedeflere iyi bir Ģekilde bağlanabilir. Sentetik nükleik asit aptamerleri, benzersiz bir yüksek afiniteli ligandlar sınıfını temsil eder (Hamaguchi ve ark., 2001). Aptamerler tedavi, hedefli ilaç verme, sensör ve tanı reaktifleri arasında değiĢen uygulamalarda potansiyel göstermektedir (Herr ve ark., 2006). Bununla birlikte, aptamerler potansiyel olarak protein konformasyonları,
21
enzimatik aktivite ve sinyalleme ağlarını modüle etmek için aktivasyon aracı olarak görev yapabilirler (Hartmann ve ark., 1998 ; Win ve ark., 2009 ). Aptamerin hedefe bağlanması, yapı uyumu, elektrostatik etkileĢimler, hidrojen bağları sayesinde veya bu etkileĢimlerin birleĢiminden kaynaklanmaktadır. Aptamer seçimi, büyük ölçüde hidrofobik karaktere sahip ve negatif yüklü moleküller için daha zordur. GeliĢtirilen ilk aptamerlerRNA‟lerden oluĢmaktadır (Ellington ve Szostak, 1990). Daha sonra, baĢka hedeflere spesifik ssDNA aptamerleri ve kimyasal olarak modifiye edilmiĢ nükleotitleri içeren aptamerler seçilmiĢtir (Green ve ark., 1995; Mi ve ark., 2005). Kimyasal modifikasyonlar aptamerlere yeni özellikler kazandırabilir, bağlanma yeteneklerini geliĢtirebilir veya stabilitelerini arttırabilir (Gold ve ark., 1995). Aptamerler, kiral molekülleri ayırt edebilir. Bu özellikleri sayesinde hedef molekülün farklı bir epitopunu ayırt edebilir (Michaud ve ark., 2003; Jenison ve ark., 1994). Yüzey alanının geniĢ olması sebebiyle hedef molekülde oluĢan en küçük farklılık bile aptamerin bağlanmasını engelleyebilir. Srinivasan ve arkadaĢlarının geliĢtirdiği ADP aptameri sadece bir fosfat grubu fazla olmasına rağmen ATP‟ye bağlanmayarak etkin bir Ģekilde ayırt edebilmektedir (Srinavasan ve ark., 2004). Aptamerler bu özellikleri sayesinde, belirli bir protein ailesindeki benzer proteinleri ya da aynı proteinin farklı biçimsel yapılarını dahi birbirlerinden ayrılabilmektedirler(Darmostuk ve ark., 2015). Proteinler, mükemmel aptamer hedefleri yapan çok geniĢ, çok iĢlevli yüzeyler sergilerler. Trombin, normalde nükleik asitlerle etkileĢmeyen aptamer seçiminde çalıĢılan ilk protein hedefidir. Anti-trombin DNA aptameri G-kuartet yapısına katlanır ve trombin fonksiyonunu inhibe edebilir. Bu aptamer, en sık çalıĢılan aptamerlerden biridir ve aptamer uygulamaları için yaygın olarak model sistem olarak kullanılmaktadır(Bock ve ark., 1992).
IgE, IFN-g, alfa thrombin gibi terapötik hedeflere karĢı çok sayıda aptamer seçilmiĢtir (JF Lee ve ark. 2006). Örneğin, Pfizer® tarafından pazarlanan Pegaptanib (Macugen), yaĢa bağlı maküler dejenerasyonun (AMD) tedavisinde, vasküler epidermal büyüme faktörüne (VEGF) karĢı seçilen bir RNA aptamer olarak kullanılır (Darmostuk ve ark., 2015). Bugüne kadar terapötikler için kullanılan en baĢarılı aptamer, anti-vasküler endotelyal büyüme faktörü (anti-VEGF) aptameridir ve ticari olarak onaylanmıĢ ilk aptamerdir (Yüce ve ark., 2018). VEGF, gözlerde anormal yeni kan damarlarının geliĢimini destekler ve görme kaybına neden olur. Aptamer, vitreus boĢluğuna doğrudan
22
uygulanarak ve VEGF'ye bağlanarak kullanılır ve VEGF'nin reseptörüne bağlanmasını inhibe eder (Cunningham ve ark. 2005). Klinik araĢtırmalar, hastaların%80‟inden fazlasının tedaviden üç ay sonra stabil veya düzelmiĢ görme gösterdiğini saptanmıĢtır ve FDA tarafındanonaylanmıĢtır. Yakın zamanda kullanılması beklenen baĢka bir aday aptamer, antikoagülan aptamerdir. Heparinin etkisinin uzun süre devam etmesi ve farmakokinetiğinin kontrol edilmesi güç olmakla birlikte, Gilead, koroner arter baypas greft cerrahisi sırasında geçici antikoagülan aktiviteyi arttırmaktadır ve bu aptamer faz I klinik denemelerini geçmiĢtir (Bock ve ark. 1992). Ayrıca pıhtılaĢma faktörü IXa için geliĢtirilmiĢ bir RNA aptameri, heparinin yerini alma kapasitesine sahiptir. Bu aptamer, 2'fluoropirimidin içerir ve kandaki yarı ömrünü uzatan bir kolesterol grubuna konjuge edilir. Ek olarak bu aptamere karĢı 17 baz çifti 2‟-O-metil RNA oligonükleotitleri antidotu geliĢtirilmiĢtir. Antidot, aptamere bağlanır, yapısını bozar ve antikoagülan aktivitesinin tersine çevrilmesinin daha iyi kontrol edilmesine ve daha iyi zamanlamaya olanak tanıyarak inaktif hale getirir (Bayraç, 2011).TanımlanmıĢ tekli hedef moleküllere ek olarak, SELEX teknolojisi karmaĢık hedef komplekslerinde, hücrelerde ve dokularda da kullanılabilir. Bu durumda, son aptamer havuzu, potansiyel hedef moleküllerin sayısı ve yoğunluğuna, aynı zamanda aptamerlerin afinitesine bağlı olarak daha karmaĢık olabilir. Kompleks karıĢımlara yönelik bu SELEX deneylerinin çoğunda, seçilen aptamerler genellikle proteinler olan hücre yüzey moleküllerini hedef almaktadır. Aptamer seçimi, örneğin çevre koĢullarındaki değiĢikliklerin veya hastalıkların neden olduğu hücre yüzey yapılarındaki değiĢikliklere yönelik olabilir (Blank ve ark., 2001; Darmostuk ve ark., 2015).
23
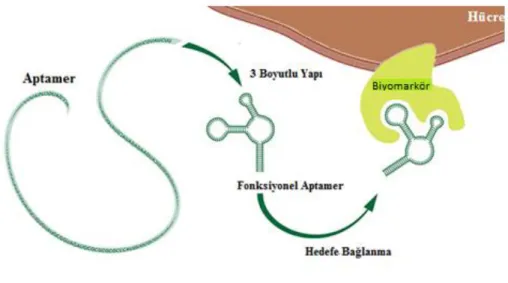
ġekil 2- 7 Aptamerin hedefe bağlanması (Sun ve ark., 2014)
DNA aptamerleri birçok avantaja sahiptir; yüksek düzeyde kararlıdırlar, kolay ve ucuz olarak üretilebilirler, oda sıcaklığında saklanabilirler, çok komplekskimyasal modifikasyonları kolayca yapılabilir, hedef analitlere yüksek özgüllük ve afinitede bağlanabilmektedirler (Hamaguchi ve ark., 2001). Antikor ve protein-bazlı diğer sensörler tek kullanımlık olmasına rağmen aptamerler yeniden katlanabilme özelliklerini korudukları için tekrar tekrar kullanılabilmektedirler (Guo ve ark., 2008). DNA aptamerlerinin hedefe bağlanması (ġekil 2-7) Watson-Crick baz eĢleĢmesinde bulunan adenin-timin ve guanin-sitozin eĢleĢmesi gibi baz eĢleĢmesi ile olmamaktadır. Aptamer seçiliminde kullanılan DNA farklı dizilerle elde edilen farklı Ģekillerle bağlanmayı sağlamaktadır ve hedef küçük moleküller, proteinler makromoleküller, hücreler olabilmektedir (Darmostuk ve ark., 2015).
Aptamerler bir in vitro çoğaltma ve seçme yöntemi olanSELEX teknolojisi ile geliĢtirilmektedir. Bu teknik 1013
-1014 molekül içeren bir sentetik oligonükleotit havuzundan hedefe bağlanabilen dizilerin zenginleĢtirerek seçilmesinden oluĢmaktadır. Varyasyon sayısı değiĢken bölgenin uzunluğuna bağlıdır. Örneğin 25 oligonükleotitin değiĢken bölgesi ile teorik olarak 425
(≅ 1015) farklı oligonükleotit dizisi elde edilebilmektedir (Strehlitz ve ark., 2008).Oligonükleotit havuzu 20-80 arası rastgele nükleotitler içeren ve iki ucunda sabit primer bulunan diziler bütünüdür. SELEX iĢlemi sırasında oligonükleotit havuzu hedef ile inkübe edilir ve bağlanmayan diziler yıkanarak
24
uzaklaĢtırılır. Bağlanan diziler hedeften ayrılır ve uçlarındaki primer bölgelerinden polimeraz zincir reaksiyonu (PZR) ile çoğaltılırlar. ZenginleĢtirilen bu ikincil havuz tek zincirli hale getirilerek SELEX‟in ikinci turunda kullanılmak için hazır hale getirilir. Bu iĢlem hedefin kompleksliğine bağlı olarak da 6 ila 20 tur arasında değiĢim göstermektedir. Hedefe bağlanan dizilerin yeterince zenginleĢtirilebilmesi için SELEX protein ve küçük organik moleküllerde 10-15 tur, canlı hücreler gibi daha kompleks hedeflerde ise 20 tura kadar tekrarlanmaktadır (Pasut ve Veronese, 2012). Çok sayıda SELEX turu yapmanın bir sebebi de iĢlem sona erdirildiğinde dizileme iĢleminde zenginleĢtirilmiĢ aptamerleri yakalayabilmektir. Hedefe karĢı pozitif bir seçilim uygulanırken amaca göre bağlanması istenmeyen hedeflere karĢı negatif bir seçilim de uygulanabilmektedir. Böylece oligonükleotit havuz hedefe karĢı zenginleĢtirilirken istenmeyen moleküllere bağlanması engellenebilir. SELEX turları ile zenginleĢtirilen havuzlar hedefe belli bir afinite ile bağlandığında seçilim sona erdirilir ve son oligonükleotit havuz dizilenir. Dizileme sonucu elde edilen sonuçlarda en çok rastlanan diziler aday aptamer olarak seçilir ve hedefle oluĢturdukları kompleksin çözüĢüm katsayıları (Kd) hesaplanır. DüĢük çözüĢüm katsayısına sahip diziler aptamer olarak
seçilir ve ilerleyen çalıĢmalarda kullanmak için hazırdır (Sefah ve ark., 2010).
Aptamerler hayvanlardan veya hücre hatlarından bağımsız bir in vitro prosesle seçilir, böylece SELEX süreci fizyolojik olmayan koĢullar altında da uygulanabilir. Toksik hedef moleküller veya düĢük immünojenisitesi olan moleküller için aptamerlerin seçimi mümkündür. Seçimi istenen aptamer özelliklerine veya aptamerlerin amaçlanan uygulamalarına yönlendirilmesi için temel SELEX iĢleminde çeĢitli modifikasyonlar yapılabilir. Seçimlerinden sonra aptamerleri modifiye etmek için, çoğunlukla stabilitelerini arttırmak veya aptamerlerin miktarını ve immobilizasyonunu sağlamak için çeĢitli yöntemler geliĢtirilmiĢtir (örneğin, nükleotit seviyesinde kimyasal modifikasyonlar, fonksiyonel gruplar). Bununla birlikte, Ģimdiye kadar metastatik kanserin moleküler belirlenmesinde çok az aptamer probu bildirilmiĢtir. Metastatik kanserin erken teĢhisi ve uygun tedavisi için metastatik kanser hücrelerini spesifik olarak tanıyabilen moleküler ligandların geliĢtirilmesi büyük önem taĢımaktadır.
Aptamerler, ekspresyon kasetlerinin transkripsiyonu ile hücrelere verilebilir veya hücreler içinde (intramerler) ifade edilebilir. Bu tür stratejiler, proteinlerin canlı hücreler veya organizmalar bağlamında iĢlevini ve etkileĢimini incelemeye izin verir (Famulok,
25
2001). Aptamerler, hedef moleküllerin hücre içi tespiti için kullanılabilir veya hücre içi inhibitörler olarak iĢlev görebilir (Burgstaller, 2002). Aptamerler, biyomedikal alanda, yüksek özgüllük ve bağlanma afinitesi, kolay sentez, kolay modifikasyon, küçük boyut, toksik olmama ve iyi stabilite gibi birçok avantaja sahip bir biyo-biliĢim elemanları sınıfı olarak giderek artan bir Ģekilde uygulanmaktadır. Kanser gibi birçok hastalık morfolojik ve moleküler düzeyde hücresel sapmalar gösterir. Uygun tanıma molekülü ve verimli bir sinyal iletim sistemi kullanıldığında, moleküler özelliklere dayanan tıbbi teĢhis oldukça spesifik ve son derece hassas olabilir(Hassan ve ark., 2017). Geleneksel biyo-tanıma molekülü, antikor birçok formatta mükemmel tanı tahlilleri geliĢtirmek için kullanılmıĢtır. Antikor kullanımına kıyasla aptamerlerin bazı avantajları vardır. Aptamerler kolay ve ucuz bir Ģekilde üretilebilirler, kimyasal olarak modifiye edilebilirler, yüksek termal kararlılıktadırlar, protein yapılar olmadıkları için çevresel koĢullara daha dayanıklılardır, raf ömürleri daha uzundur, taĢıma için özel Ģartlar gerektirmezler, kurutulmuĢ formda yıllarca oda sıcaklığında saklanabilirler, sentez ve modifikasyon proseslerindeki maliyetleri çok daha düĢüktür, antijenlere göre daha küçük olması hedef baĢına bağlanabilme sayılarını oldukça arttırmaktadır ve hücre içi, dokular arası taĢınmasını kolaylaĢtırmaktadır (Yan ve Levy,2009). Aptamerler, taĢıyıcı materyal üzerinde immobilizasyondan sonra veya hayvanlara aktarıldıktan sonra bağlanma ve inhibitör davranıĢlarını koruyabilir ve çeĢitli fonksiyonel gruplarla etiketlenebilirler. Aptamerlerin bu özellikleri, saflaĢtırma iĢlemlerinde ve ilaç geliĢtirmede kullanılmaktadır (Guo ve ark., 2008).
Antikorlar protein yapıda moleküllerdir ve sıcaklıkla çok kolay denatüre olabilirler. Fakat aptamerler denatürasyon-renatürasyon arasında bir döngüye sahiptirler. Bu özelliğinden dolayı aptamerler hedef moleküle bağlandığında doğal konformasyonuna dönebilir ve çok geniĢ assay koĢulları altında kullanılabilirler (Mascini ve ark. 2008). Antikorların aksine, aptamerler kimyasal olarak sentezlenebilirler ve bu özellikleri sayesinde ilaç geliĢtirmede önemli bir avantaj sağlarlar(Guo ve ark., 2014).Dizisi bilinen bir aptamer, DNA sentezi yapabilen herhangi bir firmadan uygun fiyatlara elde edilebilir ve uçlarına amino, tiyol, floresans vb. modifikasyonlar kolaylıkla eklenebilir. Diğer bir özellikleri ise aptamerlerin düĢük toksisiteye sahip olmalarıdır. Buna karĢın antikorlar belirgin ölçüde immünojeniktir ve bu etkisi doz ile iliĢkilidir (Ireson ve Kelland 2006). Eyetech çalıĢma grubu Vaskuler Endotelyal Büyüme Faktörüne (VEGF)
26
özgü aptamerlerin maymunlara 1000 kat daha fazla yüksek dozda verildiğinde dahi çok düĢük immün etki oluĢturduğunu gözlemlemiĢlerdir (Eyetech, 2002). Aptamerlerde hedef bölgeyi araĢtırmacı seçebilirken antikorlarda hedef bölgeyi immün sistem belirler. Aptamerler herhangi bir hedefe karĢı seçilebilirken, toksik veya non-immünojenik moleküllere karĢı antikorlar geliĢtirmek oldukça zordur (Jayasena, 1999). Aptamerleri terapötik ilaçlar olarak kullanarak, ilaç aktivitesini seçici olarak kontrol edebilen eĢleĢtirilmiĢ antidotlar kolaylıkla üretilebilir (Rusconi ve ark., 2004). Aptamerlerin daha geniĢ bir kullanımını terapötik olarak sınırlandıran bir dizi eksiklik vardır. Örneğin kan akıĢında aptamer ömrünü arttırmak ve böbrek filtrasyonu ile hızlı temizlenmelerini önlemek için aptamere genellikle bir polietilen molekülü eklenir (Bouchard ve ark., 2010). Bu konjugasyon aptamerin molekül ağırlığını arttırır ve kandaki ömrünü uzatır. Tüm bu değiĢiklikler, aptamerlerin üretim maliyetini önemli ölçüde arttırmaktadır. Aptamerlerin çoğu, protein hedefinin saflaĢtırılmıĢ, çözünür formları kullanılarak seçilmiĢtir.
Genel olarak, SELEX deneylerinden elde edilen en iyi sonuçlar, hedef protein, kararlı bir konformasyon sağladığında, yapısal bir epitopun bir tur seçimden diğerine tutarlı bir Ģekilde sunulmasına izin verdiği zaman elde edilir(Lee ve ark., 2006). Hücre yüzeyinde birçok ilginç terapötik hedef veya tanı belirteçleri ifade edilir. Ayrıca, birçok hücre proteini diğer hücre yüzey bileĢenleri ile koreseptörler olarak iĢlev görür. Bu proteinlerin hücre yüzeyi üzerinde kopyalanması, optimal fonksiyonel aktivite ve ortak reseptör alt birimleri tarafından aptamer bağlanması için ek epitoplar sağlayabilir (Hwa ve ark., 2008). Çizelge 2-3‟de aptamer- antikor arasındaki temel farklılıklar genel olarak gösterilmiĢtir.
Aptamer Antikor
Belli bir hedef molekülüne karĢı geliĢtirilmektedir.
Toksik veya immünojenik olmayan moleküllere karĢı geliĢtirilemezler. Seçim süreci daha ucuz ve daha kısadır. Seçim süreci pahalı ve zaman isteyen bir
süreçtir.