TARIM BILIMLERI DERGISI 2004, 10 (2) 187-190
Mortality Factors and Age-specific Mortality Rates of
Chionaspis
salicis
L. (Hom.: Diaspididae) on Poplar Trees in Van, Turkey
*
Remzi ATLIHAN 1 M. Salih
özGOKçE l
Geliş Tarihi: 15.09.2003
Abstract: Willow scale, Chionaspis salicis L. (Ham.: Diaspididae) is a major pest of poplar trees in Van. To understand population structure and density, and to determine reasons for population fluctuation of C. salicis, its population dynamics were investigated and a life table was constructed in 2000. Data were collected in three poplar tree plantations. Sampling was carried out every fortnight and 5 cm 2 parts of the stern of poplar trees were used as sampling units. C. salicis had one generation per year in Van. Generation mortality was extrennely high and approximately 71% of it occurred during the egg-crawler stage. Egg-crawler deaths caused by various factors were the most important mortality factors underlying fluctuations in the population. Parasitism and predation contributed to total generation mortality at the pre-ovipositing female stage and caused mortality rates of 14.7% and 18.6% respectively. Zaomma Iambinus (Walker) (Hym.: Encyrtidae) and Inostemma sp. (Hym.: Platygasteridae) were found as parasitoids, and Chilocorus bipustulatus L. (Col: Coccinellidae) and Temnostethus sp. (Hem.: Anthocoridae) as predators.
Key Words: Chionaspis salicis, life table, poplar trees
Chionaspis salicis L. (Hom.: Diaspididae)'in Van'da Kavak A
ğ
açlar
ı
Üzerindeki
Ölüm Faktörleri ve Ya
ş
a Ba
ğ
l
ı
Ölüm Oranlar
ı
Özet: Chionaspis salicis L. (Ham.: Diaspididae) Van (Türkiye)'da kavak ağaçlarının önemli bir zararlısıdır. C.
salicis'in populasyon yapısı ve yoğunluğu hakkında bilgi edinmek ve populasyon değişimlerinde rol oynayan faktörleri belirlemek için 2000 yılında Van'da zararlının populasyon gelişmesi izlenmiş ve yaşam çizelgesi oluşturulmuştur. Örneklemeler üç kavak plantasyonunda 15 günde bir yapılmış ve kavak ağalannın gövdesi üzerinde 5 cm 2 lik alanlar örnekleme ünitesi olarak kullanılmıştır. C. salicis'in Van'da yılda bir döl verdiği belirlenmiştir. Bir döldeki ölüm oranı oldukça yüksek olarak bulunmuş ve bunun yaklaşık %71'i yumurta - hareketli larva döneminde meydana gelmiştir. Çeşitli faktörler tarafından meydana getirilen yumurta - hareketli larva ölümlerinin zararlı populasyonundaki değişmelere neden olan ana faktör olduğu belirlenmiştir. Parazitizm ve predasyon pre-ovipozisyon döneminde görülmüş ve bu dönemde meydana getirdikleri ölüm oranları sırasıyla %14.7 ve %18.6 olarak bulunmuştur. Zararlının parazitoidleri olarak Zaomma
lambinus (Walker) (Hym.: Encyrtidae) ve Inostemma sp. (Hym.: Platygasteridae), predatörleri olarak ise Chilocorus bipustulatus L. (Col: Coccinellidae) ve Temnostethus sp. (Hem.: Anthocoridae) bulunmuştur.
Anahtar Kelimeler: Chionaspis salicis, yaşam çizelgesi, kavak ağacı
Introduction
Scale insects are important pests of many annual and perennial plants in and out of agricultural areas all over the world (Borchsenius 1966, Yanin 1976, Kozstarab and Kozar 1988). One of them, Chionaspis salicis L. (Hom.: Diaspididae), is widely distributed and is an important pest of various shrubs and trees in Turkey (Bodenheimer 1949, Yıldız 1972, Çanakçıoğlu 1977). It is especially a pest of poplar trees (Populus spp.) in Van in eastern Turkey where these trees are widely growrçand important for the local economy (Yaşar et al. 1995). Its feeding activity (sucking plant sap) causes stunted growth and dying of young trees in heavy infestations. There is no empirical information about the population dynamics of C.
salicis or about the factors which are responsible for the
variation in its population density. In order to devise a sound management plan, it is important to understand the population structure and dynamics of any pest species and to determine factors (biological and physical mortality factors) causing the population fluctuation. For this reason
the population dynamics of C. salicis were investigated and a life table was constructed in order to determine the temporal variation in population density and the reasons for population fluctuations. Important data, which could be used for developing descriptive models and control strategies for C. salicis, were obtained by determining age-specific mortality rates and mortality factors. The contribution of various mortality factors to generation mortality of C. salicis was calculated and mortality factors were determined.
Material and Methods
The study was carried out in three poplar (Populus
nigra L.) plantations of the same age located in different
places in Van region: Akköprü which is on North Hemp., Hatuniye on South Hemp., and Iskele on West Hemp. during 2000. No control practices were applied for C. Supported by Yüzüncü Yıl Univ. Research Foundation
188 TARIM BİLİMLERİ DERGİSİ 2004, Cilt 10, Sayı 2
salicis during the study. Four trees were selected
randomly for sampling in each of plantation. Due to the fact that C. salicis prefers the stern of the tree, one stern part with a surface area of 5 cm 2 was selected randomly as a sampling unit from alt four sides of each selected tree. Sampling was carried out every fortnight and the number of dead and alive individuals of C. salicis in the sampling unit was counted for each stage (egg, first and second instars, male, pre-ovipositing female and ovipositing female) except crawler and the causes of mortality were recorded. The number of crawlers was derived from the differences between hatched eggs and first instar nymphs due to the fact that they disperse.
With the data obtained by sampling in three plantations, a life table describing the mortality rates and factors in each age interval of C. salicis was constructed (Harcourt 1969, Krebs 1972, Morris and Miller 1954, Morris 1963):
age of cohorts,
the number of individuals entering x over an entire generation
the number dying from various mortality factors
mortality factors responsible for dx, percentage mortality of /x.
The life table included five C. salicis age intenıals (x): Egg-crawler (Age interval of scale before settling was shown in a row as egg-crawler), first instar, second instar, pre-ovipositing female and ovipositing female.
Four mortality factors were identified: Parasitoids, predators, othersı and others2. The percentage parasitism was determined by counting parasitoid exit holes. Percentage predation was determined by counting scales which showed symptoms of feeding (chewed edges or ragged holes in them). Othersı represents wind dispersal, failure to find suitable sites for settling and unexplained death caused by various factors before settling. Others2 represents unexplained death caused by various factors and individuals that disappeared by unknown factors in sampling units after settling.
The relative contribution of the individual mortality factors to the generation mortality of C. salicis was determined by expressing mortalities as k, values, a logarithmic measure of the killing power of a mortality factor. The killing power of an individual mortality factor (k,) was calculated as the following:
ki= logN,J — logNij +1,
where Nii is the number of individuals entering the
i
th
stage before action of the mortality factor, and Ni] + ı is thenumber of surviving individuals of the
j
th
stage after action of mortality factor (Varley and Gradwell 1960). Total generation mortality (K) was calculated by summing submortalities (individual k values) as follows:Total mortality K = ki+ k2 + k3 + k4,
where ki= parasitism, k2 = predation, k3 = Othersı, ka = Others2.
lndividual k values were plotted against the log of the number entering the stage (age interval) on which it acts to determine density-dependence of a mortality factor (Southwood 1978). A mortality factor was assumed density-dependent when a positive relationship was found between submortality factor (k,) and the log of the number entering stage with coefficient, b, significantly greater than 0 (Luck 1971).
Results
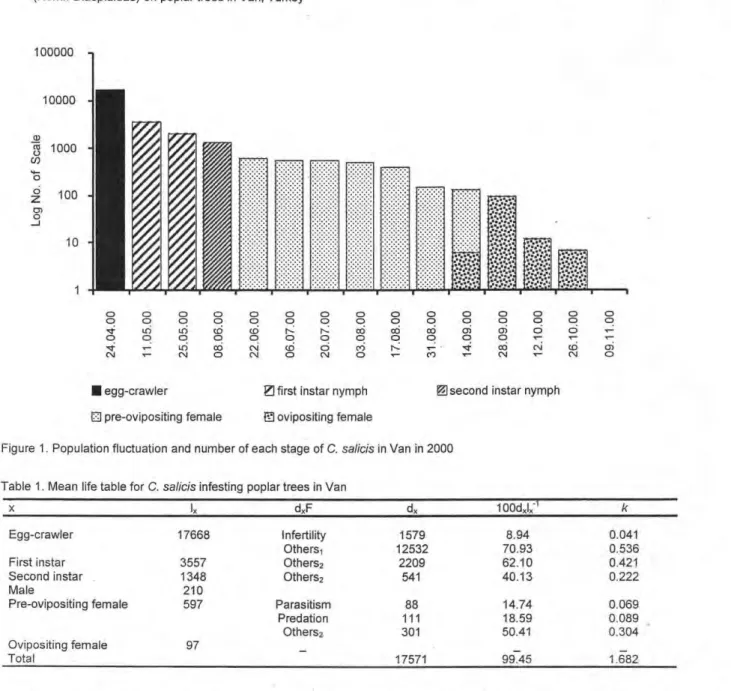
C. salicis had one generation per year in Van, overwintering in the egg stage beneath the scale cover. Hatching occurred in the first quarter of May and the crawlers settled mainly on the stern and rarely on the shoots of the poplar trees. The first and second instar nymphs completed their development by the end of the June and pre-ovipositing female appeared in the second half of June. The ovipositing female appeared in the middle of September for the first time and alt of them died during the first two weeks of November (Fig. 1). Sex discrimination was apparent (by the shape of scale cover) from the second fortnight of June and sex ratio was 73.9 : 26.1 (female : male).
The population dynamics of C. salicis was summarized in a life table, representing three locations (Table 1). Approximately 9% of deposited C. salicis eggs failed to hatch because of unknown reasons. Possible causes of this egg mortality may have included natural egg sterility, disease and undetected predation. After egg hatching a large number of crawlers died (Table 1). Othersı (k3) had the highest contribution to the decline of the egg-crawler population and caused about 71c/0 mortality. This is apparent from the high k value of othersı
(k3) shown in Table 1. Others2 (k4) made a considerable
contribution to mortality of the first and second instar nymphs and pre-ovipositing female. The greatest contribution to total generation mortality (K) was made by others2 (k4) after scale settling (Tablet). Parasitism was observed at only the pre-ovipositing female stage and mortality caused by parasitoids was lower than other mortality factors operating at this stage. Zaomma lambinus (Walker) (Hym.: Encyrtidae) and Inostemma sp. (Hym.: Platygasteridae) were the only parasitoids recovered. The contribution of predation to scale mortality could only be observed at the pre-ovipositing female stage although predators were observed Ön the trees from May to the end of October. The ladybeetle Chilocorus bipustulatus L. (Col: Coccinellidae) (as chewing predator) and Temnostethus sp. (Hem.: Anthocoridae) (as piercing-sucking predator) were the predators recovered. C. bipustulatus was more encountered than Temnostethus sp. at alt scale stages except the egg-crawler stage. Rates of predation might be higher than observed because the contribution of
Temnostethus sp. to mortality could not be determined
and some of the scale individuals that disappeared at the first and second instars might have been connpletely consumed by C. bipustulatus. Overall generation mortality of C. salicis was found to be 99.45% and after settling of the crawlers only 2.7% of scale survived.
O O O) 0 cd N o 0 0 csi 0 0 c..) o o oi 0 0 O LC; •1"-• 0
cm
0 0 0 aD 0 O N o 0 aD o (-5 O O o O o N: N: O o Cd O O (.1 o o o o 0•
0
• Od N OZ first instar nymph
ovipositing female
■ egg-crawler
O pre-ovipositing female
second instar nymph 100000 -
10000 -
1000
100
10
Figure 1. Population fluctuation and number of each stage of C. salicis in Van in 2000
Log
No.
o
f Sca
le
ATLIHAN, R. and M. S. ÖZGÖKÇE, "Mortality factors and age-specific mortality rates of Chionaspis salicis L. (Hom.: Diaspididae) on poplar trees in Van, Turkey"
Table 1. Mean life table for C. salicis infesting poplar trees in Van
x
Ix
dx F d x 1 00dx1„1 kEgg-crawler 17668 Infertility 1579 8.94 0.041 Othersi 12532 70.93 0.536 First instar 3557 Othersz 2209 62.10 0.421 Second instar 1348 Othersz 541 40.13 0.222
Male 210
Pre-ovipositing female 597 Parasitism 88 14.74 0.069 Predation 111 18.59 0.089 Othersz 301 50.41 0.304 Ovipositing female 97
Total 17571 99.45 1.682
189
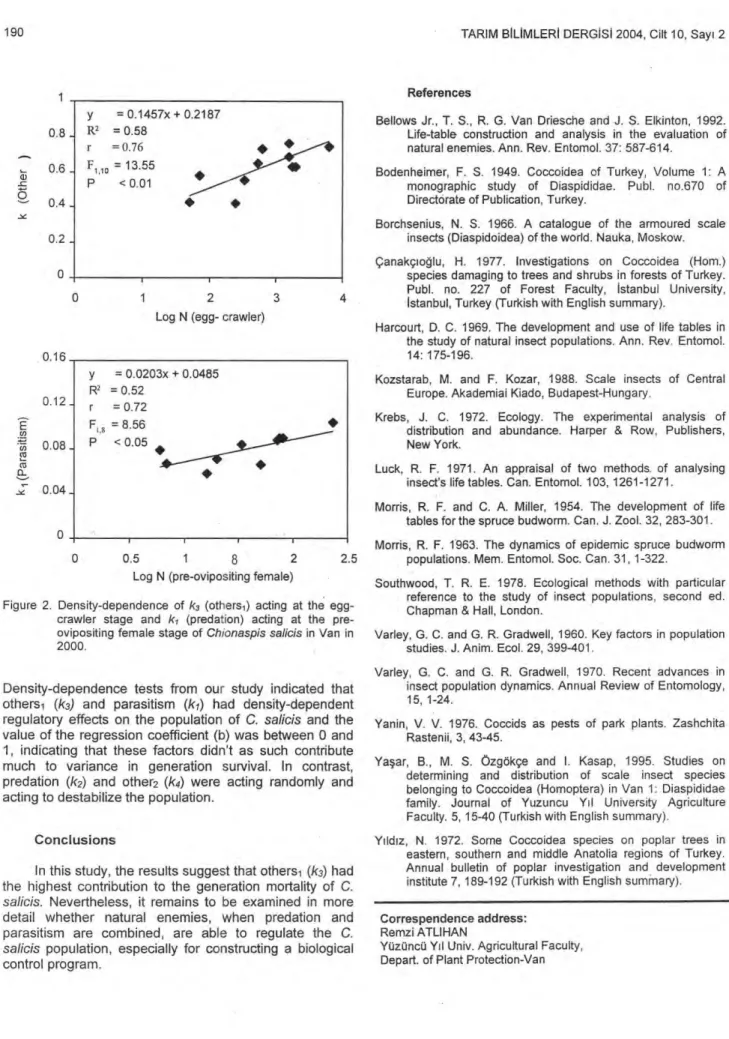
From density-dependence tests, a significant positive relationship was found between ki and log (initial density) of the pre-ovipositing female, and between k3 and log (initial density) of egg-crawler stage (Fig 2). The slopes for acting on pre-ovipositing female and k3, acting on egg-crawler stage were significantly greater than 0 (P > 0.05), which implies density-dependence. Some regression coefficients for the relationship between the other mortality factors and the number of scales on which they acted were positive, but smaller than 0.05, while some were negative, implying inverse density-dependence.
Discussion
The number of eggs that did not hatch was not very high, but after eggs hatching considerable mortality was observed at the crawler stage. Reasons for high crawlers mortality could be wind dispersal, failure to find suitable settling sites and undetected predation. Predation might impose considerable mortality at the egg-crawler stage, because the predators (C. bipustulatus and Temnostethus
sp.) were observed at this scale stage. Others2 (k4), which acts on the first instar, second instar and pre-ovipositing female stages, appeared to be of considerable importance in contributing to total generation mortality. Killing power
(k) values of this mortality factor show its contribution clearly. The rate of predation could have been higher if its real contribution could have been determined at all scale stages. Owing to this reason, taking the possible underestimation of predation at all scale stages into account, the possibility of the predation being able to regulate the population of C. salicis should not be excluded. The mortality rate caused by parasitism at the pre-ovipositing female stage was 14.74%. It was suggested that this rate was important for an organism in a biological control program (Bellows et a1.1992).
Varley and Gradwell (1970) stated that density-dependent mortality factors could cause variation in generation survival when the regression coefficient (b) is between 1.0 and 2.0. According to Luck (1971), variance in generation survival can be caused by both density-dependent and density-indensity-dependent mortality factors.
0 1 2 3 Log N (egg- crawler)
E o_ 016 0.12 _ 0.08 _ 0.04 _ 0.5 1 8 2 2.5 y = 0.1457x + 0.2187 R' = 0.58 r = 0.76 = 13. 55 P < 0.01 0 0 y = 0.0203x + 0.0485 R' = 0.52 r = 0.72 F18 = 8.56 P < 0.05 •
190 TARIM BİLİMLERİ DERGISI 2004, Cilt 10, Sayı 2
Log N (pre-ovipositing female)
Figure 2. Density-dependence of k3 (othersı) acting at the egg-crawler stage and ki (predation) acting at the pre-ovipositing female stage of Chionaspis salicis in Van in 2000.
Density-dependence tests from our study indicated that othersı (k3) and parasitism (ki) had density-dependent regulatory effects on the population of C. salicis and the value of the regression coefficient (b) was between 0 and 1, indicating that these factors didn't as such contribute much to variance in generation survival. In contrast, predation (k2) and other2 (k4) were acting randomly and acting to destabilize the population.
Conclusions
In this study, the results suggest that othersı (k3) had the highest contribution to the generation mortality of C.
salicis. Nevertheless, it remains to be examined in more detail whether natural enemies, when predation and parasitism are combined, are able to regulate the C.
salicis population, especially for constructing a biological control program.
References
Bellows Jr., T. S., R. G. Van Driesche and J. S. Elkinton, 1992. Life-table construction and analysis in the evaluation of natural enemies. Ann. Rev. Entomol. 37: 587-614.
Bodenheimer, F. S. 1949. Coccoidea of Turkey, Volume 1: A monographic study of Diaspididae. Publ. no.670 of Directörate of Publication, Turkey.
Borchsenius, N. S. 1966. A catalogue of the armoured scale insects (Diaspidoidea) of the world. Nauka, Moskow. Çanakçıoğlu, H. 1977. Investigations on Coccoidea (Hom.)
species damaging to trees and shrubs in forests of Turkey. Publ. no. 227 of Forest Faculty, İstanbul University,
İstanbul, Turkey (Turkish with English summary).
Harcourt, D. C. 1969. The development and use of life tables in the study of natural insect populations. Ann. Rev. Entomol. 14: 175-196.
Kozstarab, M. and F. Kozar, 1988. Scale insects of Central Europe. Akademiai Kiado, Budapest-Hungary.
Krebs, J. C. 1972. Ecology. The experimental analysis of distribution and abundance. Harper & Row, Publishers, New York.
Luck, R. F. 1971. An appraisal of two methods of analysing insect's life tables. Can. Entomol. 103, 1261-1271.
Morris, R. F. and C. A. Miller, 1954. The development of life tables for the spruce budworm. Can. J. Zool. 32, 283-301. Morris, R. F. 1963. The dynamics of epidemic spruce budworm
populations. Mem. Entomol. Soc. Can. 31, 1-322.
Southwood, T. R. E. 1978. Ecological methods with particular reference to the study of insect populations, second ed. Chapman & Hali, London.
Varley, G. C. and G. R. Gradwell, 1960. Key factors in population studies. J. Anim. Ecol. 29, 399-401.
Varley, G. C. and G. R. Gradwell, 1970. Recent advances in insect population dynamics. Annual Review of Entomology, 15, 1-24.
Yanin, V. V. 1976. Coccids as pests of park plants. Zashchita Rastenii, 3, 43-45.
Yaşar, B., M. S. Özgökçe and I. Kasap, 1995. Studies on determining and distribution of scale insect species belonging to Coccoidea (Homoptera) in Van 1: Diaspididae family. Journal of Yuzuncu Yıl University Agriculture Faculty. 5, 15-40 (Turkish with English summary).
Yıldız, N. 1972. Some Coccoidea species on poplar trees in eastern, southern and middle Anatolia regions of Turkey. Annual bulletin of poplar investigation and development institute 7, 189-192 (Turkish with English summary).
Correspendence address: Remzi ATLIHAN
Yüzüncü Yıl Univ. Agricultural Faculty, Depart. of Plant Protection-Van