FEN BİLİMLERİ ENSİTİTÜSÜ
FARKLI SEVİYELERDE KALSİYUM VE ORGANİK MANGAN İÇEREN RASYONLARIN
BÜYÜYEN JAPON BILDIRCINLARDA PERFORMANS VE BAZI DOKULARDA
KALSİYUM VE MANGAN KONSANTRASYONUNA ETKİLERİ
Ahmet ARICI YÜKSEK LİSANS TEZİ ZOOTEKNİ ANABİLİMDALI
T.C.
SELÇUK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
FARKLI SEVİYELERDE KALSİYUM VE ORGANİK MANGAN İÇEREN RASYONLARIN BÜYÜYEN JAPON BILDIRCINLARDA
PERFORMANS VE BAZI DOKULARDA KALSİYUM VE MANGAN KONSANTRASYONUNA ETKİLERİ
Ahmet ARICI
YÜKSEK LİSANS TEZİ ZOOTEKNİ ANABİLİMDALI
T.C.
SELÇUK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
FARKLI SEVİYELERDE KALSİYUM VE ORGANİK MANGAN İÇEREN RASYONLARIN BÜYÜYEN JAPON BILDIRCINLARDA
PERFORMANS VE BAZI DOKULARDA KALSİYUM VE MANGAN KONSANTRASYONUNA ETKİLERİ
Ahmet ARICI
YÜKSEK LİSANS TEZİ ZOOTEKNİ ANABİLİMDALI
Bu tez 25/02/2010 tarihinde aşağıdaki jüri tarafından oybirliği/ oyçokluğu ile kabul edilmiştir
Doç. Dr. Alp Önder YILDIZ Prof. Dr. Yılmaz BAHTİYARCA
(Danışman) (Üye)
Yrd. Doç. Dr. Yusuf KONCA (Üye)
FARKLI SEVİYELERDE KALSİYUM VE ORGANİK MANGAN İÇEREN RASYONLARIN BÜYÜYEN JAPON BILDIRCINLARDA PERFORMANS
VE BAZI DOKULARDA KALSİYUM VE MANGAN KONSANTRASYONUNA ETKİLERİ
Ahmet ARICI Selçuk Üniversitesi Fen Bilimleri Enstitüsü Zootekni Anabilim Dalı
Danışman: Doç Dr. Alp Önder YILDIZ 2010, sayfa: 57
Jüri: Doç. Dr. Alp Önder YILDIZ Prof. Dr. Yılmaz BAHTİYARCA
Yrd. Doç. Dr. Yusuf KONCA
Bu çalışma, farklı seviyelerde kalsiyum (Ca) ve organik mangan (Mn) içeren rasyonların büyüyen bıldırcınlarda besi performansı ve bazı dokularda Ca ve Mn konsantrasyonuna etkisini tespit etmek maksadıyla yapılmıştır. Denemede mısır-soya küspesi ağırlıklı rasyon (% 24 HP, 2900 kcal/kg ME, % 0.50 metiyonin, % 1.30 lisin, % 0.80 Ca ve 60 ppm Mn) kontrol rasyonu olarak kullanılmıştır. Denemede 4 farklı Ca (% 0.40, 0.80, 1.60 ve 2.40) ve 3 farklı Mn (0, 60 ve 120 mg/kg) seviyeleri
ile bunların kombinasyonlarından oluşan 12 muamelenin etkisi araştırılmıştır. Deneme 35 gün sürmüş ve hayvanlar deneme boyunca ad libitum olarak yemlenmiştir.
Muamele grupları arasında deneme sonu ortalama canlı ağırlık, canlı ağırlık artışı, yem tüketimi, karkas randımanı, kemik külü, kemik Ca ve Mn konsantrasyonu ile plazma Ca konsantrasyonu bakımından önemli farklılıklar vardır. Ancak, deneme rasyonları; yem değerlendirme katsayısını, yaşama gücünü ve plazma Mn konsantrasyonunu önemli olarak etkilememiştir. Sonuç olarak, büyüyen Japon bıldırcınlarında % 0.40 Ca içeren rasyona 120 mg/kg seviyesinde organik Mn ilavesinin uygun olabileceği söylenebilir.
Anahtar kelimeler: Bıldırcın, kalsiyum, mangan, performans, kemik,
ABSTRACT Master Thesis
The effects of rations containing different levels of calcium and organic manganese on
performance and some tissues calcium and manganese concentration of Japanese quails
Ahmet ARICI Selcuk University
Graduate School of Natural and Applies Sciences Department of Animal Science
Supervivor: Assoc. Prof. Dr. Alp Önder YILDIZ 2010, 57 Pages
Jury: Assoc. Prof. Dr. Alp Önder YILDIZ Prof. Dr. Yılmaz BAHTİYARCA
Assis. Prof. Dr. Yusuf KONCA
This study was carried out to determine the effects of diets containing different levels of calcium (Ca) and organic manganese (Mn) on fattening performance and some tissues calcium and manganese concentration of Japanese quails. In the experiment, the diet based corn-soybean meal (22 % crude protein and 2900 kcal/kg metabolizable energy) is used as control. In the experiment, the effect of four different levels of dietary Ca (0.40, 0.80, 1.60 and 2.40 %) and three different levels of dietary organic manganese (0, 60 and 120 mg/kg) and, the effeccts of 12
treatments consisiting their interactions were investigated. The experiment was lasted for 35 days period. The birds were fed as ad libitum.
There were significant differences among the treatment groups in terms of average final body weight, body weight gain, feed consumption, carcass percentage, bone ash, the Ca and Mn concentrations of bone and the Ca concentration of plasma. But, the experimental diets did not effected the average feed conversion ratio, mortality and plasma Mn concentration of quails. As a result, ıt may be say that 120 mg/kg supplementation manganese to diets containing 0.40 % Ca is suitable in growing Japanese quails.
TEŞEKKÜR
Yüksek lisans eğitimin başından sonuna kadar her türlü yardımını esirgemeyen, ileriki yaşamımda ve mesleki hayatımda bana rehberlik edecek olan, her türlü konuda bilgi ve tecrübelerini aktaran çok değerli danışman hocam sayın Doç. Dr. Alp Önder YILDIZ’a en içten teşekkürlerimi ve saygılarımı sunarım.
Yüksek lisans eğitimim ve araştırmamın her safhasında bilgi ve yardımını esirgemeyen sayın Prof. Dr. Yılmaz BAHTİYARCA ve Yrd. Doç Dr. Yusuf CUFADAR ‘a ve hayvancılık işletmesinde, laboratuvardaki çalışmalarımda ve istatistikî analizlerin yapılmasında bana yardımcı olan Arş. Gör. Sayın Osman OLGUN olmak üzere bütün, öğretim elemanlarına en içten teşekkürlerimi ve saygılarımı sunarım.
Yüksek lisans eğitimimde desteklerini hiç esirgemeyen aileme de sonsuz şükranlarımı sunarım.
25.02.2010 Ahmet ARICI
İÇİNDEKİLER Sayfa No ÖZET ………..………...i ABSTRACT...……….,,,…..iii TEŞEKKÜR……….……...v İÇİNDEKİLER……….…….………...vi SİMGELER……….………....vii ÇİZELGE LİSTESİ………....viii 1.GİRİŞ……….…...1 2. KAYNAK ARAŞTIRMASI………...4 2.1.Kalsiyum Özellikleri………..…………...….4
2.1.1.Kalsiyumun absorbsiyonu ve atılması………..………...5
2.1.2.Kalsiyumun fonksiyonları………..6 2.1.3.Kalsiyumun yetersizliği………..………....6 2.1.4.Kalsiyum fazlalığı………..………...7 2.1.5.Kalsiyum kaynakları………..……….7 2.1.6.Kalsiyum gereksinimi………..………...8 2.2.Manganez………..………....9
2.2.1.Manganezin emilmesi ve atılması………10
2.2.2.Manganezin fizyolojik fonksiyonları………..………...11
2.2.3.Manganez yetersizliği……….………...12
2.3.Büyüyen Japon Bıldırcınlarında Besin Madde İhtiyaçları………...13
2.4.Japon Bıldırcınlarında, Broyler ve Yumurta Tavuklarında Kalsiyum ve Manganez ile Yapılmış çalışmalar………....…………...……..15
3. MATERYAL METOT……….…..24
3.1. Materyal..……….…...24
3.1.1.Hayvan materyali………..……….…...24
3.1.2.Yem materyali………..………....25
3.2. Metot………..…….25
3.2.1.Deneme rasyonlarının hazırlanması ve grupların oluşturulması………..…...25
3.2.2.Denemenin yürütülmesi………..…….27
3.2.2.1.Performansla ilgili kriterlerin tespiti………..…………...27
3.2.2.2.Kan numunelerinin alınış tekniği………..………....28
3.2.2.3.Plazma kalsiyum ve manganez konsantrasyonunun tespiti………28
3.2.2.4.Tibia kalsiyum ve manganez konsantrasyonunun tespiti……...29
3.2.2.5.Yemdeki kalsiyum ve manganez konsantrasyonunun tespiti……..29
3.2.3.İstatistik metotlar………..…29
4. ARAŞTIRMA SONUÇLARI………..……...31
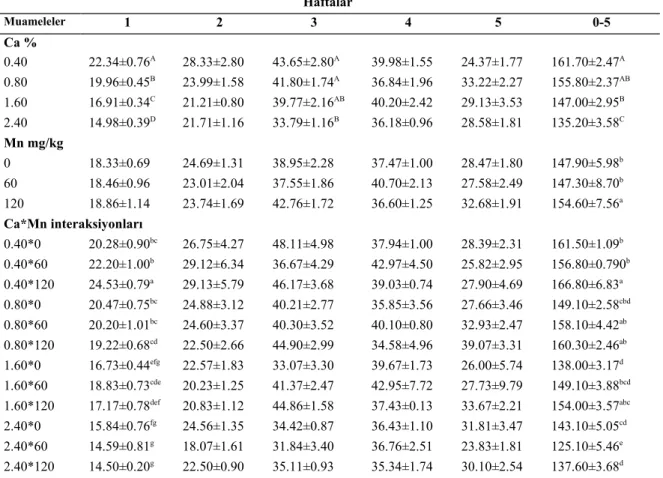
4.1.Canlı Ağırlık………...31
4.2.Canlı Ağırlık Artışı ……… ………..……….…..32
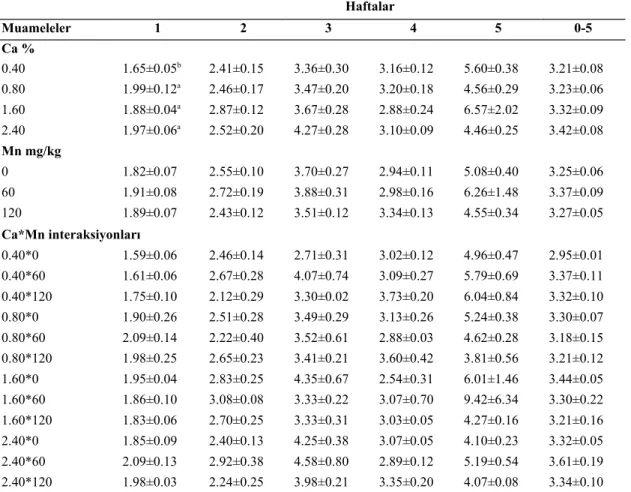
4.3.Yem Tüketimi………...……….…...34
4.4.Yemden Değerlendirme Katsayısı………....35
4.5. Yaşama Gücü………...36
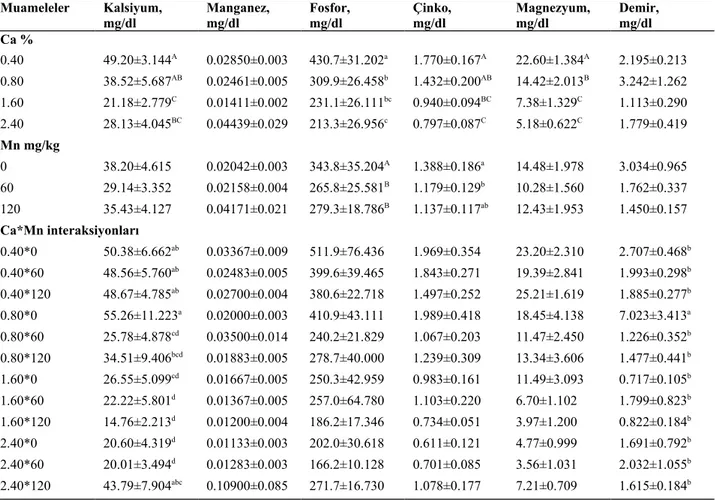
4.6.Karkas Randımanı, Karaciğer Ağırlığı ve Kemik Mineral Değerleri…….…….37
4.7.Plazma Mineral Değerleri………...40
5. TARTIŞMA ve SONUÇ……….42
6. KAYNAKLAR……….……….…47
7. EKLER……….………...52
SİMGELER
Ca : Kalsiyum
CA : Canlı Ağırlık CAA : Canlı Ağırlık Artışı DCP : Dikalsiyum Fosfat
Fe : Demir
HP : Ham Protein
g : Gram
kcal : Kilo Kalori
kg : Kilogram KP : Kullanılabilir Fosfor ME : Metabolik Enerji mg : Miligram ml : Mililitre Mn : Mangan Mg : Magnezyum P : Fosfor
SFK : Soya Fasülyesi Küspesi YA : Yumurta Ağırlığı
YG : Yaşama Gücü
YDK : Yem Değerlendirme Katsayısı YV : Yumurta Verimi
YT : Yem Tüketimi
ÇİZELGE LİSTESİ
Çizelge No Sayfa No
Çizelge 2.1. Çeşitli türlerin kalsiyum gereksinimleri………...…8 Çizelge 2.2. Büyüyen Japon bıldırcınlarının protein , aminoasit ve yağ
ihtiyaçları………...13 Çizelge 2.3. Büyüyen Japon bıldırcınlarının makro ve iz element ihtiyaçları…....…14 Çizelge 2.4. Büyüyen Japon bıldırcınlarının vitamin ihtiyaçları…..……...……..….14 Çizelge 3.1. Araştırmada kullanılan rasyonların hammadde ve hesaplanmış besin maddeleri kompozisyonu………….………...26 Çizelge 3.2. Araştırmada kullanılan rasyonların analiz edilmiş mineral madde
içerikleri………...…….27 Çizelge 4.1. Rasyon kalsiyum ve manganez seviyesinin büyüyen Japon
. bıldırcınlarda CA (g/bıldırcın) etkisi ...………..……….….32 Çizelge 4.2. Rasyon kalsiyum ve manganez seviyesinin büyüyen Japon
bıldırcınlarda CAA (g/bıldırcın/hafta) etkisi,………...33 Çizelge 4.3. Rasyon kalsiyum ve manganez seviyesinin büyüyen Japon
bıldırcınlar yem tüketimine (g/bıldırcın/hafta) etkisi……….35 Çizelge 4.4. Rasyon kalsiyum ve manganez seviyesinin büyüyen Japon
bıldırcınlarda YDK (yem/CAA) etkisi ………...…36 Çizelge 4.5. Rasyon kalsiyum ve manganez seviyesinin büyüyen Japon
bıldırcınlarda karkas randımanı, karaciğer ağırlığı ve kuru kemikteki mineral seviyesine etkisi ………...39 Çizelge 4.6. Rasyon kalsiyum ve manganez seviyesinin büyüyen Japon
bıldırcınlarda plazmada mineral seviyesine etkisi………….……..…..41 Ek Çizelge 1. Farklı seviyelerde Ca ve Mn İçeren rasyonların büyüyen Japon
bıldırcınlarda CA‘ğa etkisine ait varyans analiz sonuçları…………...52 Ek Çizelge 2. Farklı seviyelerde Ca ve Mn içeren rasyonların büyüyen Japon
bıldırcınlarda CAA’na etkisine ait varyans analiz sonuçları………...….53 Ek Çizelge 3. Farklı seviyelerde Ca ve Mn içeren rasyonların büyüyen Japon
bıldırcınlarda YT’ne etkisine ait varyans analiz sonuçları...54 Ek Çizelge 4. Farklı seviyelerde Ca ve Mn içeren rasyonların büyüyen Japon
bıldırcınlarda YDK’nın etkisine ait varyans analiz sonuçları…………..55 Ek Çizelge 5. Farklı seviyelerde Ca ve Mn içeren rasyonların büyüyen Japon
bıldırcınlarda karkas randımanı, karaciğer ağırlığı ve kuru kemik mineral seviyesinin etkisine ait varyans analiz sonuçlar………....56 Ek Çizelge 6. Farklı seviyelerde Ca ve Mn içeren rasyonların büyüyen Japon
bıldırcınlarda plazma mineral değerlerine etkisine ait varyans analiz sonuçları……….……….……….…………57
Bıldırcın yetiştiriciliği bazı Avrupa ve Uzakdoğu ülkelerinde özellikle de Japonya da çok eskiden beri yapılmaktadır. Bıldırcınların kafeste yetiştirilmeye uygun olmaları, birim alanda daha fazla yetiştirilebilmeleri, yumurta verimlerinin yüksek oluşu, erken yaşta cinsi olgunluğa ulaşmaları, hızlı büyümeleri ve generasyonlar arası sürenin kısalığı sebebiyle yılda 4-5 generasyon üretilebilmeleri (Anonymous 1969), bu hayvanların ticari olarak et ve yumurta üretiminde kullanılmasını teşvik edici rol oynamaktadır. Bıldırcın ülkemizde av hayvanı olarak bilinmektedir ve sınırlı sayıdaki işletmede bıldırcın yetiştiriciliği yapılmaktadır. Bıldırcın üretiminin ve tüketiminin arttırılması için ürün fiyatlarının makul seviyelere getirilmesi ve tüketicinin ilgisini çekecek reklamlar yapılıp etinin ve yumurtasının besleyici özelliği hakkında bilgilendirilmesi gerekmektedir. Günümüzde gün geçtikçe bıldırcın üretimi ve ürünlerinin tüketimi artmaktadır.
Hayvan beslemede verimliliğin artmasındaki en önemli sebeplerinden birisi de ihtiyaç duyulan besin maddelerinin, optimum seviyede karşılanmasıdır. Bu doğrultuda hayvanın besin madde ihtiyacının tespiti önemlidir. Kanatlı rasyonlarında büyük miktarlarda ve yaygın olarak bulunan iki yem hammaddesinden birisi dane mısırdır. Dane mısır, kanatlı rasyonlarında % 50 veya daha yüksek seviyelerde kullanılabilmektedir. Ancak dane mısırın manganez (Mn) seviyesi düşüktür. Manganez kemik gelişimde rol oynayan elementlerden birisidir. Bir buğdaygil danesi olan mısırın Ca içeriğinin de yüksek olduğu söylenemez. Rasyonda aşırı miktarda kalsiyum (Ca) olduğunda da ihtiyaç duyulan Mn miktarının arttığı bilinmektedir. Ayrıca elementlerin organik formlarının inorganik olanlara göre kullanılabilirliklerinin daha iyi olduğu ifade edilmektedir (Henry ve ark., 1989; Kidd ve ark.,1992).
Hayvan vücudundaki Mn miktarı gayet düşüktür. Birçok dokuda element sadece iz miktarlarda bulunur. Kemikler, karaciğer, böbrekler, pankreas ve pituiterde elementin konsantrasyonu diğer dokulardan daha yüksektir. Element hidrolaz ve
kinaz grubu enzimlerin aktivatörü ve arjinaz, piruvat karboksilaz ve superoksid mutaz gibi enzimlerin de kompanentidir. Elementin pratik şartlarda noksanlığına ruminantlarda, domuzlarda ve kanatlılarda rastlanılmamıştır. Elementin akut noksanlığının etkileri bütün türlerde aynı olup, bunlar; büyümenin gerilemesi, iskelet anormallikleri, yeni doğanlarda ataksi ve üreme performansında düşmedir. Element kemiğin mukopolisakkaritlerinin sentezi için gerekli glikozil transferaz enziminin katalitik etkisi için gereklidir (Yazgan 2006).
Civcivlerde Mn noksanlığı ayak kemiklerinin teşekkülünde perosis veya tendon kayması denilen anormalliğe sebep olur. Fakat perosisin oluşumunda Mn eksikliği tek faktör olmayıp, kolinin eksikliği, Ca ve fosfor (P) elementlerinin eksikliği de perosise sebep olabilir. Kolin ve Mn arasındaki bir diğer bağlantı karaciğer yağlanmasında görülür. Mangan noksanlığı damızlık kanatlılarda da kuluçka parametrelerinin, kabuk kalınlığının ve diğer kabuk kalite kriterlerinin düşmesine ve civcivlerde başın geriye doğru dönmesine sebep olur.
Yemlerde ve çayır-mer’a otlarında Mn yaygın bulunur. Kaba yemler (özellikle çayır ve mer’a otları) 40-200 mg/kg kuru madde (KM) Mn içerikleriyle elementin zengin kaynaklarıdırlar. Çayır otlarının Mn içerikleri yetişme şartlarına bağlı olarak gayet değişkendir. Danelerin ve yan ürünlerin Mn içerikleri nispeten iyidir (tek istisnası mısır olup mısırın Mn muhtevası düşüktür). Mayaların ve hayvani orijinli gıdaların Mn içerikleri düşüktür. Pirinç kepeği ve değirmencilik artığı yemler Mn’ın zengin kaynaklarıdırlar.
Manganezin toksik dozu ile yemlerdeki normal seviyesi arasında oldukça geniş bir emniyet aralığı mevcuttur. Tavuklarda 1 g/kg KM seviyesinde verilen Mn’in toksik etkisi görülmemiştir. Ayrıca Mn normal kemik gelişiminde, amino asit metabolizmasında ve enzim fonksiyonlarında rol oynayan önemli elementlerden birisidir (Scott ve ark. 1976).
Bazı araştırıcılar kanatlılarda rasyonda aşırı Ca olduğunda Mn kullanımının olumsuz yönde etkilendiğini bildirmektedirler ( Leach ve Lilburn 1987; Underwood 1981; Hurley ve Keen 1987). Ancak Scheideler (1991) civcivlerde rasyon Ca seviyesinin Mn kullanımını azaltmadığını belirtmiştir. Wedekind ve ark. da (1991) rasyonda aşırı Ca’un Mn emilimini ve endojen Mn atılımını olumsuz yönde etkilemediğini bildirmiştir. Ji ve ark.(2005) organik ve inorganik Mn kaynaklarını kullanarak broylerlerde yaptıkları bir çalışmada organik Mn kaynaklarının emiliminin inorganik Mn kaynaklarından daha yüksek olduğunu ifade etmişlerdir. Bu çalışma, farklı seviyelerde kalsiyum (Ca) ve organik mangan (Mn) içeren rasyonların büyüyen bıldırcınlarda besi performansı ve bazı dokularda (kan ve kemik) Ca ve Mn konsantrasyonuna etkisini tespit etmek maksadıyla yapılmıştır.
2.KAYNAK ARAŞTIRMASI
2.1. Kalsiyumun Özellikleri
Ca hayvan vücudunda en fazla bulunan elementtir. Dişlerin ve kemiklerin önemli bir unsurudur. Vücutta mevcut toplam Ca % 99 gibi büyük bir kısmı kemikler ve dişlerde, geriye kalan % 1’lik kısmı ise yumuşak dokular ve vücut sıvılarında bulunur. Başta sinir uyarılarının iletilmesi ve kasların kasılıp gevşemesi için gerekli olan enzimler olmak üzere, Ca birçok enzim sisteminin aktivitesi için gereklidir. Element aynı zamanda kanın pıhtılaşması olayında da görev yapar. Yumurtlayan tavuklarda plazma Ca seviyesi çok daha yüksek olup, 300-400 mg/litre arasındadır (Yazgan, 2006).
Kemikte en bol bulunan iki element Ca ve P dur. Bir kilogram kemik külünde 360 g Ca, 170 g P ve 10 g Mg bulunur. Kimyasal anlamda iskelet dokusu stabil bir yapıda olmayıp, bu dokudan devamlı olarak Ca ve P elementlerinin mobilizasyonu ve bu elementlerin kemikte yeniden depolanmaları olayları vuku bulur. Kan ve diğer yumuşak dokularla kemikler arasında Ca alışverişi devamlı bir işlem olmakla beraber, kemiklerden Ca mobilizasyonu paratiroid bezince kontrol edilir. Hayvanlar düşük Ca’lu rasyonlarla beslendiklerinde hücre dışı sıvısında Ca konsantrasyonu düşer, bu durumda paratiroid bezi stimüle edilir ve artan paratiroid hormonu (PTH) kemiklerden Ca mobilizasyonunu hızlandırır ve hayvanın Ca ihtiyacının bir kısmı böylece kemiklerden karşılanır. Kemiklerde Ca, P ile birlikte bulunduğundan serbest hale geçen Ca ile birlikte P’da serbest hale geçer ve P’un fazlası idrarla dışarı atılır. Paratiroid hormonu 1,25 dihidroksikolekalsiferol sentezini etkileyerek ince bağırsaklarda absorbe edilen Ca’un regülasyonunda da rol oynar. 1,25 dihidroksikolekalsiferol bir vitamin D derivatı olup, Ca bağlayıcı protein (CaBP) sentezini hızlandırır. Paratiroid hormonu ayrıca idrar Ca ‘nun böbreklerden kana reabsorbsiyonunuda hızlandırır. Hormon kan Ca seviyesini artırıcı etkiye sahiptir (Yazgan, 2006).
2.1.1. Kalsiyumun absorbsiyonu ve atılması
Kalsiyumun büyük bir kısmı ince bağırsakta çok az bir kısmı da midede absorbe edilir. Hayvanlar Ca’u gereksinimlerine göre absorbe ederler. Gereksinimindeki değişiklik absorbsiyon etkinliğini de değiştirir. Absorbsiyon üzerine vitamin D, protein, genç hayvanlarda laktoz ve fitin, rasyonun P içeriğinin fazla oluşu, yağ, serbest yağ asitleri, verim, yaş, rasyonun Ca ve P düzeyleri ile oksalik asit etki eder. Vitamin D gereksinimi optimal düzeyde karşılanan hayvanlarda Ca absorbe edilme oranı düşük olur. Yapılan araştırmalarda nitrojensiz rasyonlarla beslenen hayvanlarda Ca’un absorbe edilemediğini göstermiştir. Bu durum Ca’un absorbsiyonu üzerine rasyonun protein içeriğinin önemli bir etkisi olduğu kanısını uyandırmaktadır. Genç memelilerde laktoz, süt Ca’unun emilme oranın arttırır. Kalsiyumun emilebilmesi, emildiği yerdeki eriyebilme durumuna bağlıdır. Asit ortam Ca’u eriyebilir durumda tutar. Sindirim kanalında laktik asite dönüşen laktoz, Ca’un emilmesi üzerine bu şekilde etki yapar. Rasyonlarda bulunan fitin genç hayvanlarda Ca’un emilimini azaltır. Rasyonda P’un fazla oluşu, erimeyen trikalsiyum fosfat oluşumuna neden olarak Ca’un emilmesine olumsuz etki yapar. Bu nedenle Ca’un P’a oranı çok önemlidir. Rasyonda bulunan bir miktar yağın Ca’un emilmesini arttırmasına karşılık serbest yağ asitleri, erimeyen Ca sabunları oluşturduğu için Ca’un emilmesini kısmen engellemiş olur. Kalsiyumun emilimi, rasyonun flor ve Ca düzeylerinin yüksek olması ve yaşın ilerlemesi ile azalır(Okuyan, 1997).
Kalsiyumun bağırsaklarda absorbe edilmeyen kısmı gübre ile dışarı atılır. Ancak gübrede bulunan Ca’un bir kısmını sindirim salgıları ile ince bağırsağa gelen endojen Ca oluşturur. Kalsiyumun atılımı tükürük ile başlar. Tükürükteki Ca yoğunluğu kan serumundakinden daha yüksektir. Sindirim salgıları ile boşaltılan Ca’un en büyük kısmını, ince bağırsak salgısı ile gelen Ca oluşturur. Gübre ile boşaltılan Ca’un %20‘si safra ile boşaltılan Ca’dan ibarettir. İdrarla da oldukça yüksek miktarda Ca boşaltılır ve miktarı hayvanın yaşına göre değişir. Bununla birlikte Ca sütle, yumurta ve terle de boşaltılır (Okuyan 1997).
2.1.2. Kalsiyumun fonksiyonları
Kalsiyumun fonksiyonları şu şekilde özetlenebilir. -Kemiklerin ve dişlerin en yapısal öğesidir.
-Kanın pıhtılaşma mekanizmasında protrombinin trombine dönüşmesi için kanda bulunması zorunludur.
-Bazı enzimatik reaksiyonlarda koenzim ya da enzim aktivatörü olarak görev yapar.
-Serum Ca düzeyi düştüğü zaman sinirlerin duyarlılığı artar. Aşırı düşüklük tetaniye neden olur ve tedavi edilmez ise ölüme götürür.
-Sodyum ve potasyum ile birlikte kalp atışını düzenler.
-Bazı hormonların ve hipotalamustan kaynaklanan hormon salgılatan faktörlerin salgılanmaları için ihtiyaç duyulur.
-Kas kontraksiyonu, hücre zarı geçirgenliği ve kalp kaslarının fonksiyonları için gereklidir (Okuyan 1997).
2.1.3. Kalsiyumun yetersizliği
Kalsiyum yetersizliğinin en önemli hastalığı raşitizmdir. Bu hastalıkla kartilajın ve kemiğin organik matriksinde Ca ve P yoğunluğu azalır. Kalsiyum ya da P yetersizliği vitamin D yetersizliğine benzer semptomlara neden olur. Osteomalasia ergin hayvanlarda raşitizmin kopyasıdır ve kartilaj büyümesi durduğu için kemik matriksinde Ca ve P yoğunluklarının azalması ile karakterize edilir. Kalsiyum ve P’un yetersizliklerinin işaretleri esas olarak genç hayvanlarda görülür. Yetersizlik, kemik sisteminde karakteristik semptomlar ortaya çıkmadan önce, büyümenin durması, ağırlık kaybı ve iştahın azalması ya da kaybolması ile olur, kemiklerin minerilzasyonu azalır. Bu durum her dönemde topallığa ve kolay kırılmaya ve büyüme dönemi süresince de anormal kemik oluşumuna da neden olur.
Kümes kanatlılarında belirtileri; kemik mineralizasyonunda gerileme, kemiklerin kolayca kırılmaları, kül ve Ca içeriklerinin normalin yarısına kadar düşmesidir. Yumurta tavuklarında yumurta verimi azalır ya da durur. Yem tüketimi
ve yemin değerlendirmenin gerilemesi, yumurta kabuğu kalitesinin azalması, yumurta içi kalitesinde düşüş, yumurta büyüklüğünde ve ağırlığında azalma ve döl verimi bozukluklarıdır. İskelette, kemik rezorpsiyonunda artış, kafes yorgunluğu, osteoporosis, raşitizm, felç, yumuşak gaga, kaslarda katılık, topallık, kaburgalarda yumrular, eklemlerde genişleme, anormal duruş ve biçimsiz kemik oluşumu gibi semptomlar ortaya çıkar (Okuyan 1997).
2.1.4. Kalsiyum fazlalığı
Rasyonda Ca düzeyinin yüksek oluşu; proteinler, yağlar, vitaminler, P ve Mn gibi makro elementler ile özellikle demir, iyot, çinko ve Mn gibi iz elementler dahil, rasyondaki diğer besin maddelerinin değerlendirilmesini olumsuz etkiler. Bu besin maddeleri rasyonda gereksinim düzeylerinde bulundukları zaman bile, Ca fazlalığı ters etki yapabilir. Bunun sonucu, söz konusu besin maddelerinin yetersizlik semptomlarının ortaya çıkmalarına neden olabilir. Gereksinimden fazla Ca tüketildiği zaman, çinkonun organizmada dağılımı düzensizleşir. Bu da parakeratoza neden olur. Ayrıca gereksinim duyulan yerlerde çinkonun yetersiz oluşu, canlı ağırlık kaybına neden olur. Ruminantlar Ca fazlalığına karşı daha dayanıklıdırlar. Yüksek düzeyde Ca’un etkisi altında kalan süt inekleri doğumdan sonra süt humması hastalığına sebep olabilir(Okuyan 1997 ).
2.1.5.Kalsiyum kaynakları
Başta baklagil yeşil yemleri olmak üzere bütün yeşil yemler, şeker pancarı posası, süt ve süt mamulleri Ca bakımından zengindirler. Tahıllar, kök ve yumru yemler Ca’un fakir kaynaklarıdır. Bazı yemlerdeki Ca okzalik asit ile Ca okzalat formunda kompleks oluşturur. Bu formdaki Ca hayvan tarafından absorbe edilemeyip dışkı ile atılır. Kemik unu ve balık unu gibi yemler hayvani Ca’un mükemmel kaynaklarıdır. Süt sığırları ve yumurta tavuklarının yüksek Ca ihtiyaçlarını karşılayabilmek için bu hayvanların ve diğer çiftlik hayvanlarının rasyonlarına kireç taşı, dikalsiyum fosfat ve istimlenmiş (buhar ile muamele edilmiş) kemik unu gibi Ca ek yemleri katılır. Hayvana ek Ca ve P kaynağı olarak Ca fosfatlı
kaya tuzları veriliyor ise bu kaynağın toksik seviyede (>%0.25’den fazla ) flor ihtiva etmemesine dikkat edilmelidir. Aksi takdirde hayvanlarda flor toksitisesi (florosis) görülebilir. Basit mideli hayvanların rasyonlarına zaman zaman ilave edilen yağ, yağ asitleriyle Ca tuzlarının (sabun)oluşmasına sebep olarak Ca absorbsiyonunu düşürür (Yazgan 2006).
2.1.6. Kalsiyum gereksinimi
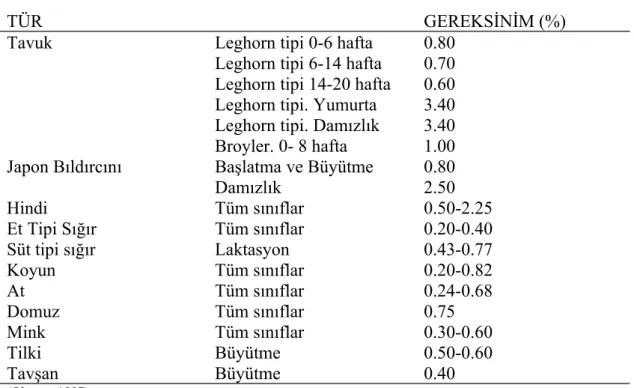
Bazı türlerin Ca ihtiyaçları Çizelge 2.1.’de verilmiştir.
Çizelge 2.1. Çeşitli Türlerin Kalsiyum Gereksinimleri
TÜR GEREKSİNİM (%)
Tavuk Leghorn tipi 0-6 hafta 0.80
Leghorn tipi 6-14 hafta 0.70 Leghorn tipi 14-20 hafta 0.60 Leghorn tipi. Yumurta 3.40 Leghorn tipi. Damızlık 3.40
Broyler. 0- 8 hafta 1.00
Japon Bıldırcını Başlatma ve Büyütme 0.80
Damızlık 2.50 Hindi Tüm sınıflar 0.50-2.25 Et Tipi Sığır Tüm sınıflar 0.20-0.40 Süt tipi sığır Laktasyon 0.43-0.77 Koyun Tüm sınıflar 0.20-0.82 At Tüm sınıflar 0.24-0.68 Domuz Tüm sınıflar 0.75 Mink Tüm sınıflar 0.30-0.60 Tilki Büyütme 0.50-0.60 Tavşan Büyütme 0.40 (Okuyan 1997). 2.2. Manganez
Manganez, bitkiler ve hayvanlar için esansiyel bir elementtir. Kümes kanatlılarının Mn gereksinimleri, diğer çiftlik hayvanlarına kıyasla oldukça yüksektir. Manganez yetersizliği bu türlerde yaygın olarak görülür. Hayvan vücudunda bulunan Mn miktarı gayet küçüktür. Çoğu dokuda element sade eser (iz) miktarda bulunur. Element kemik, karaciğer, böbrekler, pankreas ve pituiter gibi
organlarda daha yüksek miktarda bulunur. Element birçok enzimin aktivatörü ve bazı enzimlerin kompanenti olarak görev yapar. Mesela, Mn hidrolaz ve kinaz grubu enzimlerin aktivatörü arjinaz, piruvat karboksilaz ve Mn superoksid dismutaz gibi enzimlerin de yapısında bulunur (Yazgan, 2006 ).
İnorganik mineral kaynaklarından serbest hale geçen metal iyonları diğer rasyon unsurları ile kompleks oluştururlar. Bu olay ile o elementin hayvanda sindirilebilirliği ve kullanılabilirliği önemli ölçüde düşürülür. Mineraller şelat formunda veya organik formda olduklarında diğer bileşiklerle reaksiyona girmeleri önlenerek, teorik olarak bu formlardaki elementlerin absorbe olabilirlikleri, inorganik kaynaklardaki elementlerdekinden daha yüksektir. Rasyonlara şelat formunda mineral ek yemleri ilavesi günümüzde gayet aktif bir çalışma alanıdır. Şelat bileşiklerinin en çok bilinenlerinden biri sentetik bir bileşik olan etilen diamin tetra asetik asittir (EDTA). Bileşik ağır metallerle stabil kompleks oluşturur ve ağır metal zehirlenmelerinde detoksifiye edici olarak kullanılır. Şelatların elementlerin absorbsiyonuna ve kullanılabilirliklerine etkileriyle ilgili araştırma sonuçları çelişkilidir. Bazı çalışmalarda şelatların etkisi müspet olurken, diğer bazılarında sonuçlar beklenenden farklı olmuştur. Kanatlı rasyonlarında EDTA ve benzer şelat bileşiklerinin ilavesiyle bazı mineral elementlerinin kullanılabilirliklerinin iyileştiği bildirilmiştir. Bazı araştırmacılar fitik asit bakımından zengin rasyonlara şelat ilavesiyle Zn ve Mn’ın kullanılabilirliklerinin arttırıldığını bildirmişler ise de, bütün çalışmalarda benzer sonuçlar elde edilmemiştir. Şelat bileşiklerin mineral beslenmesine etkileriyle ilgili sonuçlar domuzlarda da değişken olmuştur. Bazı metallerle EDTA arasındaki bağlantı gayet kuvvetlidir. Şelat bileşiklerin mineral beslenmesine etkileriyle ilgili sonuçları değişken olmasının başlıca sebebi onların özelliklerinin farklı olmasıdır. Maliyetlerinin yüksek oluşu göz önünde bulundurularak özel uygulamalar ve durumlar dışında şelat bileşiklerinin inorganik mineral kaynaklarının yerine geçebilmeleri ihtimali çok düşüktür (Yazgan 2006).
Manganez ince bağırsağın her yerinde eşit olarak emilir. Emilme iki basamakta gerçekleşir. İlk önce ince bağırsak boşluğundan alınır ve sonra mukoza hücrelerinden taşınır. Manganezin emilme oranı tüm türlerde düşüktür. Kanatlı türlerinde ağızdan verilen dozun % 0.1’inden daha azı emilebilmektedir. Sığırlarda ise, tüketilen Mn’in yaklaşık %1’i emilir. İlk sıralarda Ca, P ve demir olmak üzere, diğer mineraller Mn’in emilmesini etkilerler. Kanatlılarda yüksek fosfat düzeyleri Mn’in emilme oranını düşürürler. Civcivlerde rasyonda yüksek demir düzeyleri, muhtemelen Mn’in emilme oranını düşürerek perosise neden olurlar. Fazla Mn ve kobalt da demir yetersizliğini teşvik eder. Manganez, vücutta düşük yoğunlukta olmakla beraber, yaygın olarak bulunur. Kemik, karaciğer, böbrekler ve pankreas Mn yoğunlukları, iskelet kaslarının yoğunluklarından daha yüksektir. Karaciğerin Mn depolama kapasitesi; bakır ve selenyum kapasitesine kıyasla sınırlıdır. Buna karşılık vücut Mn içeriğinin yaklaşık dörtte biri iskelette bulunur. Ancak bu rezerv rasyonun Mn içeriği düşük olduğu zaman pek kolay kullanılmaz. Değişik düzeylerde Mn tüketen hayvanlar, sağlıklarını ve verimlerini korumak için, değişik düzeylerde Mn absorbe ederler ve atarlar. Doku Mn düzeyleri, emilimden çok boşaltım düzeyi değiştirilerek ayarlanır. Vücut Mn havuzu küçük olduğu için, vücuttan atılan toplam Mn miktarı tüketimine hemen hemen eşit olur. Günlük tüketim vücut havuzunun % 25-50’si kadardır. Bunun için fazla Mn tüketimi hem emilme etkinliğini azaltır, hem de boşaltım oranını arttırır. Rasyonda alınan manganez, esas olarak gübre ile (% 95-98), az miktarda da idrarla (% 0.1-3) atılır ( Okuyan, 1997).
2.2.2. Manganezin fizyolojik fonksiyonları
Manganez aşağıdaki fizyolojik fonksiyonları yerine getirir;
- Enzim aktivatörüdür ve metalloenzimlerin bir öğesidir. Kemiklerin bileşimlerine girerler.
- Lipid ve karbonhidrat metabolizmalarına katılır.
- Hücre fonksiyonlarına ve hücre zarının yapımına katılır. - Bağışıklık sisteminde fonksiyonları vardır.
- Beyin fonksiyonlarını etkiler.
Diğer iz elementlerinde olduğu gibi, Mn hem bir enzim aktivatörü hem de metalloenzimlerin bir öğesi olarak görev yapar. Arginaz, piruvat karboksilaz ve Mn süperoksit dismutaz Mn içeren enzimlerdir. Manganez içeren enzimler sınırlı oldukları halde, Mn’le aktive olabilen enzimler çok fazladır. Örneğin hidrazlar, kinazlar, dekarboksilazlar ve transferazlar Mn ile aktive edilebilirler. Manganez, bir aktivatör ya da bir enzim öğesi de olsa çoğu zaman öncelikli bir katyon olarak kullanılır. Magnezyum enzim aktivitesinde çok az bir azalma ile ya da hiç azalma olmaksızın, kısmen Mn’in yerine kullanılabilir(Okuyan, 1997).
Manganezce yetersiz kemikler normalden kısa ve kalın olurlar. Manganez, kemiklerin organik matrikslerinin gelişmeleri için esansiyeldir. Organik matriks, geniş ölçüde, mukopolisakkaridlerden oluşur. Manganez yetersizliği sonucu mukopolisakkarid sentezi bozulur ki; bu da glikoziltransferazların aktive edilmeleri ile ilgilidir. Bu enzimler, polisakkarid ve glikoprotein sentezleri için önemlidirler. Manganez, gereksinilen metal iyonların en etkinidir.
Manganez yetersizliğinin ilk işaretleri üreme etkinliğinde gözlenmiştir. Yetersizlik; genç civciv, sıçan ve kobaylarda dengesizlik ile karakterize edilen doğum öncesi dönüşümsüz bir hastalığa neden olabilir. Dişi kemirgenlerde, Mn yetersizliğinin birkaç basamağı vardır. Bunlar, (1) ataksiyalı, fakat yaşayabilecek bir yavru doğumu, (2) kısa bir süre ölecek bir yavru doğumu, (3) östrusun döl verimini önleyecek şekilde düzensizleşmesidir. Östrus düzensizliği, sığır ve koyunlarda da görülür. Yumurta tavuklarında yetersizlik etkisi, yumurta veriminin azalmasına, kabuk kalitesinin bozulmasına, çıkış gücünün düşmesine ve kondrodistrofi denilen embriyonik bir yetersizlik hastalığına neden olur. İnsülin oluşumunda ya da aktivitesindeki rolü dolayısıyla, Mn yetersizliğinde glukoz kullanımı geriler. Manganez yetersizliğinde glikoproteinlerin biyosentezleri de gerileyebilir. Manganez
yetersizliğinde; hücre fonksiyonlarında anormallikler ortaya çıkar. Manganezin bağışıklık fonksiyonlarında da rolü vardır. Manganez ile nötrfiller ve makrofajlar arasında etkileşim olduğu kanıtlanmıştır (Okuyan 1997).
2.2.3.Manganez yetersizliği
Manganez yetersizliğinin esas belirtileri; büyümenin gerilemesi, iskelet anormallikleri, döl verimi bozuklukları, yeni doğanlarda yürüyüşte duraksama, sendeleme ve düşme, lipid ve karbonhidrat metabolizmalarında bozukluklar üzerinde çalışan türlerin tümünde gözlenmiştir. Ancak, bu belirtilerin gerçek görünüşleri yetersizliğin süresi ve derecesi ile değişebilmektedir.
Kümes kanatlıları; manganez yetersizliğine memelilerden daha duyarlıdırlar. Çünkü bunların Mn gereksinimleri daha yüksektir. Perosis, civcivlerde Mn yetersizliğinde en çok gözlenen bir hastalıktır. Bu hastalık; genişlemiş ve hatalı oluşmuş metatarsal eklemler, çarpık ve eğri metatarsus, kısa ve kalınlaşmış esas kemikler ve kondilinden kaymış aşil tendonu ile karakterize edilen bir kusurlu kemik oluşumudur. Bacakların biri ya da ikisi birden etkilenebilir. Kolin, biyotin ve vitamin B12 gibi diğer B grubu vitaminlerinin yetersizlikleri perosisi teşvik ederler. Hastalık, aşırı Ca ve P tüketimi ile ağırlaşır.
Damızlık tavuk rasyonlarında Mn yetersizliği; civciv embriyolarında besinsel kondrodistrofi adı ile bilinen bir hastalığa neden olur. Kondrodistrofi; kusurlu büyüme, ödem, yüksek ölüm yüzdesi, bacaklarda ve kanatlarda kısalık ve kalınlık, alt gagada papağanda olduğu gibi kısalık ve küre biçiminde bir baş ile karakterize edilir.
Manganez yetersizliği genç civcivlerde tiamin yetersizliğinde olduğu gibi sinirsel bozukluklara neden olur. Ayrıca yumurta veriminde ve yumurta çıkış gücünde düşmeye, ince kabuklu veya kabuksuz yumurta sayısında artışa neden olur (Okuyan, 1997).
Büyümekte olan Japon bıldırcınlarının besin madde ihtiyaçları Çizelge 2.2, Çizelge 2.3 ve Çizelge 2.4’ de verilmiştir.
Çizelge 2.2. Büyüyen Japon bıldırcınlarının protein, aminoasit ve yağ ihtiyaçları
(rasyonda % olarak, % 90 KM esasına göre)1.
Besin maddeleri Birim Başlama ve büyütme 29002
Protein ve amino asit
Protein % 24.0 Arjinin % 1.25 Glisin+serin % 1.15 Histidin % 0.36 İzolösin % 0.98 Lösin % 1.69 Lisin % 1.30 Metiyonin % 0.50 Metiyonin+sistin % 0.75 Fenilalanin % 0.96 Fenilalanin + tirosin % 1.80 Teronin % 1.02 Triptofan % 0.22 Valin % 0.95 Yağ Linoleik asit % 1.0 1NRC, 1994 2ME, kcal/kg
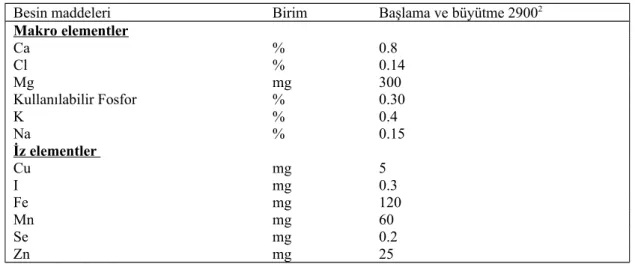
Çizelge 2.3. Büyümekte olan Japon bıldırcınlarının makro ve iz element ihtiyaçları
(rasyonun her kg’ında mg ya da % olarak, % 90 KM esasına göre)1
Besin maddeleri Birim Başlama ve büyütme 29002
Makro elementler Ca % 0.8 Cl % 0.14 Mg mg 300 Kullanılabilir Fosfor % 0.30 K % 0.4 Na % 0.15 İz elementler Cu mg 5 I mg 0.3 Fe mg 120 Mn mg 60 Se mg 0.2 Zn mg 25 1NRC, 1994 2ME, kcal/kg
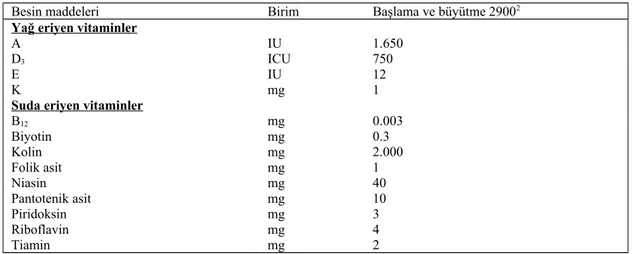
Çizelge 2.4. Büyümekte olan Japon bıldırcınlarının vitamin ihtiyaçları (rasyonun her kg için birim % olarak, % 90 KM esasına göre)1
Besin maddeleri Birim Başlama ve büyütme 29002
Yağ eriyen vitaminler
A IU 1.650
D3 ICU 750
E IU 12
K mg 1
Suda eriyen vitaminler
B12 mg 0.003 Biyotin mg 0.3 Kolin mg 2.000 Folik asit mg 1 Niasin mg 40 Pantotenik asit mg 10 Piridoksin mg 3 Riboflavin mg 4 Tiamin mg 2 1NRC, 1994 2ME kcal/kg
2.4. Japon Bıldırcınlarında, Broyler ve Yumurta Tavuklarında Ca ve Mn ile Yapılmış Çalışmalar
Bray ve ark. (1964), yumurta veriminin başlaması ile ilgili olarak meydana gelen kemik Ca depolanmasına cinsi olgunluktaki yaşın etkisini belirlemek maksadıyla yaptıkları çalışmada, erken cinsi olgunluğa ulaşan hayvanların üretimin 6-7. haftalarında iskelet ağırlıklarının % 26’sını kaybettiklerini, buna karşılık düşük Ca’lu yemle yemlenen ve geç cinsi olgunluğa ulaşan hayvanların ise iskelet ağırlıklarının sadece % 6’sını kaybettiklerini bildirmişlerdir. Bu bulguların aksine Berg ve ark. (1964), cinsi olgunluk yaşına, özgül ağırlığa, yumurta ağırlığına veya yumurta verimine devamlı ve düzenli bir etki bulamamışlardır. Anderson (1966), yumurtlama öncesi Ca, P ve Ca/P oranları farklı rasyonlarla yemlemenin yumurta verimi ve yumurta özelliklerine devamlı etkili olduğunu göstermiştir. Anderson (1967), yumurtacı piliçleri Ca seviyesi % 0.62-2.0 arasında değişen rasyonlarla yemlemiş ve çevre ile rasyon Ca seviyesi arasında oldukça yüksek bir interaksiyonun olduğunu bildirmiştir. Bu yüksek seviyedeki interaksiyon piliçlerin uygulanan yetiştirme şartlarının sınırları içinde Ca’u yumurta kabuğu teşekkülünde kullanmalarının onların yumurtlama öncesi Ca tüketimlerinin metabolik adaptasyonları ile ilgili olduğunu göstermiştir. Bunlara ilaveten piliçler ve yumurta tavuklarının sadece yumurtlama öncesi rasyon Ca seviyesi ve tabii tutuldukları fiziksel ve çevre şartlarından etkilenmedikleri, fakat yumurtlama öncesi adaptasyon
periyodunda büyüme ve endokrin sisteminin gelişme hızına da bağımlı oldukları bildirilmiştir.
Hurwitz ve Bar (1969), düşük Ca’lu (% 0.24) yemlerle yemlenen tavuklarda yumurta sayısının ve tavuğun depolayabileceği Ca miktarının onun Ca depolarının bir fonksiyonu olduğunu, tavuğun kemik depolarındaki Ca’u kullanma kabiliyetinde yaşa bağlı farklılıkların fazla olmadığını bildirmişlerdir. Araştırıcılar, erken dönemde Ca kısıtlamalarına genç piliçlerin birkaç aydır yumurtlayan tavuklardan daha hassas oldukları sonucuna varmışlardır. Yapılan bir diğer çalışmada piliçler 5,10,15 ve 19 haftalık yaşta değişik seviyede Ca içeren rasyonlarla yemlenmişler. Düşük Ca’lu (% 0.65) rasyonla yemlenen piliçlerin ayak kemiklerinde (tibia) % 2.0 veya 2.6 Ca içeren rasyonlarla yemlenen piliçlerden daha fazla Ca45 depolanmıştır (Mraz. 1972).
Hurwitz ve Bar (1971), 15 günlük yaştaki piliçlerin % 1.3-4.1 Ca ve % 0.70-0.97 P içeren rasyonlarla yemlemişlerdir. Yüksek seviyede Ca ve P içeren rasyonla yemlenen piliçlerin düşük Ca ve P içeren rasyonlarla beslenen piliçlere kıyasla yumurtlamaya başlamadan önceki iki haftada 400-500 g yumurta verimlerinin ilk ayında 80 g daha fazla canlı ağırlık artışı yaptıklarını bildirmişlerdir. Yüksek Ca ve P içeren rasyonlarla yemlenen tavuklarda aynı zamanda yumurtlama öncesi but kemiğinde Ca depolanmasının, yumurtlama döneminde yumurta ve kabuk ağırlığının arttığı gözlenmiştir. Yumurtlama öncesi rasyonlarına ilave edilen Ca ve P kemik mineralizasyonuna yardımcı olmuş ve yumurta kabuk kalitesini iyileştirmiştir. Ne yumurtlama öncesi verilen Ca ne de P yumurtlama döneminin ilk bir ayında verilen Ca’ca noksan rasyonlara tepkide herhangi bir değişikliğe sebep olmamıştır.
Yumurtacı piliçlerde ve tavuklarda yapılan bir seri çalışmada, (Miller ve Sunde,1975), yumurtlama öncesi % 0.4-4.5 arasında Ca içeren rasyonların yumurtlama döneminde yumurta verimi, yumurta ağırlığı ve kabuk kalitesini etkilemediğini bildirmişlerdir. Daha sonra Scott ve ark.(1977), 18-22 haftalık yaşlardaki piliçleri iskelette olabilecek muhtemel etkileri belirlemek maksadıyla % 1.0 ve 3.5 Ca içeren rasyonlarla yemlemişler ve % 3.5 Ca içeren rasyonla yemlenen piliçlerde canlı ağırlıkta, iskelet külünde ve Ca’unda önemli artışlar meydana
geldiğini bildirmişlerdir. Araştırıcılar 18-22 haftalık yaşlar arasındaki piliçlerde % 1.0 rasyon Ca’unun maksimum kemik kalsifikasyonu için yeterli olmadığını bildirmişlerdir. İskelet dokusunda maksimum Ca depolanması için % 3.5 rasyon Ca’unun daha uygun olacağı bildirilmiştir. Hamilton ve Cipera (1981), büyüme periyodunda % 0.51 Ca içeren rasyonla yemlenen piliçlerde ölüm oranının ve yumurtlama sırasında canlı ağırlığın, % 3.19 Ca içeren rasyonla yemlenenlere kıyasla daha yüksek olduğunu bildirmişlerdir. Classen ve Scott (1982), 25-125 günler arasında piliçleri % 0.89 veya 0.35 Ca içeren rasyonlarla ve serbest seçim Ca kaynağı ile (Ca karbonat) yemlemişledir. Bu muameleleri 177-225 günler arasında % 3.5 Ca içeren rasyonla yemleme takip etmiştir. Yumurtlama başlamadan önceki 19. günde piliçlerin Ca tüketimleri rasyonun % 2.0’sine çıkmış ve bu artış önemli bulunmuştur. Araştırıcılar piliçlerin Ca tüketiminin onların büyüme periyodu ve yumurtlama periyodundaki Ca ihtiyaçlarını yansıttığını bildirmişlerdir. Aynı zamanda araştırma sonuçları büyüme periyodundaki Ca noksanlıklarını Ca tüketimini arttırarak telafi ettiklerini bildirmişlerdir.
Wideman ve ark (1985), büyüme periyodunda yüksek Ca’lu (% 3.0-3.5) ve düşük kullanılabilir P seviyeli (% 0.38-0.40) rasyonlarla yemlenen piliçlerde böbrek lezyonları ( nefritis), iç organlar arasında Ca birikmesi sebebiyle ölüm vakalarının arttığını bildirmiştir. Büyüme periyodunda görülen ölüm vakaları yumurtlama periyodunda da görülmüştür. Yalnız bu çalışmada yüksek Ca’lu rasyonların veriliş süresi çok uzun ( 8-22 haftalar arası) olmuştur.
Yumurtacı piliçlerin 15 haftalık yaştan itibaren % 1.0, 2.0 ve 3.0 Ca içeren rasyonlarla veya 17. ve 19. haftadan itibaren 21 haftalık yaşa kadar % 2.0 ve 3.0 Ca içeren rasyonlarla beslendiği bir çalışmada (Brooks, 1986) erken yaşta cinsi olgunluğa ulaşan piliçlerin yumurtlama öncesi 17 veya 19 haftalık yaştan itibaren % 1.0 yerine % 2.0 veya % 3.0 Ca içeren rasyonlarla yemlenmesiyle serum Ca, kemik Ca, P, K, Mg ve S muhtevasının, kabuk kalitesi ve YV’nin arttırılabileceği bildirilmiştir. Bu çalışmada muamelelerin ölüm oranı, CA, CAA, karaciğer ağırlığı, karaciğer ve böbrek mineral kompozisyonuna önemli bir etkisi olmamış ise de % 2.0 ve 3.0 Ca’lı rasyonlarla YT ve YDK önemli ölçüde artarken, serum Ca seviyesi
rakamsal olarak, serum Mg seviyesi ise bazı yaş dönemlerinde önemli olarak artmıştır. Yüzde 1.0 Ca’lı rasyona nispetle daha yüksek Ca içeren rasyonlarla yumrta ve kabuk ağırlığı ve özgül ağırlık daha yüksek iken, kabukta daha kalın olup, yem tüketimi ve yemden yararlanma kabiliyeti artmıştır. Onyedi-21 haftalık dönemde % 2.0 ve 3.0 Ca’lı rasyonlarla kemik % kül ve mineral miktarı artmıştır.
Keshavarz (1987), yaptığı çalışmada büyüme periyodunda farklı sürelerle farklı seviyelerde Ca içeren rasyonlarla yemlemenin büyüme ve takip eden yumurtlama döneminde yumurtlama performansını araştırmıştır. Kontrol grubu hayvanlar % 0.8 Ca ve % 0.4 toplam fosfor (TP), muamele grupları ise % 3.5 Ca ve % 0.61 TP içeren rasyonlarla 14,15,16,17,18. haftalardan 20. haftalık yaşa kadar yemlenmişlerdir. Yüksek Ca içeren rasyonun büyüme periyodunda ağırlık artışına, yem tüketimine ve ölüm oranına menfi etkisinin olmadığı bildirilmiştir. Yüksek Ca’lu rasyonlarla yemlenen grupla kontrol grubu arasında böbrek ağırlığı, karaciğer yağ miktarı ve plazma ürik asit seviyesi arasındaki farklılıkların önemli olmadığı bildirilmiştir. Büyüme periyodunda iki hafta daha uzun süre yüksek Ca’lu rasyonlarla yemlenen grupların tibia kül ve Ca miktarları kontrol grubundan ile bazı muamele gruplarından daha yüksek bulunmuştur. Büyüme döneminde yüksek Ca’lu rasyonla yemleme cinsi olgunluğun gecikmesine neden olmuş, büyüme periyodunda değişik sürelerle yüksek Ca’lu rasyonlarla yemleme yumurtlama döneminde performans kriterlerine menfi bir etkiye sahip olmamıştır. Sonuçta araştırıcı piliçlerin yumurtlamaya başlamadan en az iki ile üç hafta önce rasyon da Ca seviyesinin arttırılmasının yetiştiricilik açısından müspet sonuçlar veren bir uygulama olduğunu, bu uygulamanın kabuk kalitesini etkin bir şekilde arttırmamış ise de, kemikte kül ve Ca miktarını arttırdığını ve bu uygulamanın avantajlı olabileceğini bildirmiştir.
Yumurtlayan Japon bıldırcınlarında Ca ve P ihtiyaçlarını tespit etmek için yapılan bir çalışmada 10 haftalık yaştaki bıldırcınlar 26 hafta müddetle 4 farklı Ca (% 2.0, 2.4, 2.8 ve 3.2) ve her bir Ca seviyesinde 3 farklı total P (%0.55, 0.70 ve 0.85) içere rasyonlarla yemlenmişlerdir (Shrivastas ve ark. 1989). Araştırıcılar % 2.8 Ca ve % 0.70 total P içeren rasyonla YV, YA ve kabuk kalitesinin arttığını bildirmişlerdir. Artan P konsantrasyonu ile serum Ca seviyesi önemli derecede
düşmüştür. Araştırıcılar yumurtlayan Japon bıldırcınlarında Ca ve total P ihtiyaçlarının sırasıyla % 2.8 ve % 0.70 total P(% 0.25 KP) ve rasyon Ca/P oranının ise 4/1 olduğu bildirmişlerdir.
Yaşlı damızlık broylerlerde rasyon kalsiyum ve bor seviyesinin etksisini tespit etmek için 2 ayrı deneme yapılmıştır (Qin ve Klandorf, 1991). İlk denemede 60 haftalık yaştaki sürüden 32 tavuk seçilmiştir. Bu hayvanlar kalsiyum seviyesi yüksek (% 3.5) ve düşük (% 1.5) olan ilave bor içeren (denemenin ilk 2 haftasında 100 mg/kg yem, diğer haftalarda 60 mg/kg) ve içermeyen rasyonlarla beş hafta boyunca yemlenmişlerdir. Yemleme denemenin ilk 2 haftasında adlibitum olarak yapılırken müteakip haftalarda tavuk başına 150 g/gün ile sınırlandırılmıştır. İkinci denemede ise, 75 haftalık yaştaki 52 adet tavuk, günde tavuk başına 50 g yulaf verilerek tüy döktürülmüştür. Tavuklar, % 25 canlı ağırlık kaybına ulaştıkları dönemden itibaren 35 gün müddetle % 1.2 kalsiyum içeren rasyonla (36 adedi), geriye kalan (16 adet) ise aynı rasyona 100 mg/kg bor ilave edilip yemlenmişlerdir. Otuzbeşinci günün sonunda aydınlatma 8 saatten 16 saate, rasyon kalsiyum seviyesi ise % 3.5’e yükseltilirken bor seviyesi 100 mg/kg olarak kalmıştır. Deneme 20 hafta sürmüştür. Sonuçta 1. denemede her 2 kalsiyum seviyesindede bor içeren rasyonlarla ortalama YV, % 3.5 kalsiyumlu rasyonla karşılaştırıldığında önemli derecede düşmüş ve YV % 3.5 kalsiyumlu rasyonla karşılaştırıldığında önemli derecede düşmüş ve YV bu düşüş yüksek kalsiyumlu rasyonla daha fazla olmuştur. İkinci denemede YV’ne muamelelelerin önemli bir etkisi olmamıştır.
Yumurtlayan Japon bıldırcınlarının farklı seviyelerde Ca (% 2.0, 2.5, 3.0 ve 3.5) ve KP içeren rasyonlarla (% 0.35, 0.50 ve 0.65) beslendiği diğer bir çalışmada ( Raju ve ark. 1992), rasyon KP seviyesinin vücutta tutulan P miktarını etkilemediği, rasyonda artan KP seviyesi ile serum Ca konsantrasyonunun önemli derecede düştüğü ve % 0.65 KP içeren rasyonla yumurta ağırlığının (YA) önemli derecede düştüğü bildirilmiştir.
Yumurtacı piliçler üzerinde yapılan bir tez çalışmasında (Tombul, 2004) yumurtacı piliçleri büyüme döneminin 16-20 haftaları arasında % 2.0, 2.5, 3.0, 3.5 ve
4.0 Ca içeren rasyonlarla beslemenin yumurtacı piliçlerde performans, bazı kemik özelliklerine ve müteakip yumurtlama dönemi (20-36 hafta) performansa etkisi incelenmiştir. Büyütme dönemi rasyon Ca seviyesi, piliçlerin CA, CAA, ölüm oranı, kuru kemik (tibia) ağırlığını, kemik külünü, kemik Ca ve P seviyeleri ile plazma Ca ve P seviyeleri önemli derecede etkilemezken, yem tüketimini önemli olarak (P<0.05) etkilemiştir. Yüzde 3.5 ve 4.5 Ca içeren rasyonla yemlenen piliçlerin yem tüketimi, % 3.0 Ca’lı rasyonla beslenen piliçlerden önemli derecede yüksek bulunmuştur. Büyütme rasyonları tavukların deneme sonu Ca’larını, 20-36 haftalık dönemdeki CAA, ölüm oranı, yumurta verimi, yumurta ağırlığı ve kitlesi, yem değerlendirme katsayısı, kemik ağırlığı ve külünü, kabuk kalite kriterlerinden birim yumurta yüzey alanı başına kabuk ağırlığını ve yumurta kabuğunda günlük depolanan Ca miktarını önemli olarak etkilememiştir.
Yapılan çalışmalarda 14 ve 25 mg/kg Mn içeren rasyonlarıyla yemlenen broylerlerde bu seviyedeki Mn’ın normal büyüme, kemik külü ve perosisin önlenmesi için yeterli olduğunu bildirmekle (Watson ve ark. 1970,1971) birlikte NRC (1994) Japon bıldırcını ve broylerler rasyonlarında 60 ppm Mn tavsiye etmektedir. Ancak pratik şartlar altında yetersizlik ihtimaline karşı sıklıkla rasyondaki Mn seviyesi daha yüksek tutulur. Farklı kaynaklarından 1000 mg/kg seviyesine kadar Mn’ın toksik olmadığı bildirilmektedir (Black ve ark. 1985, Miles ve ark. 2003).
Hayvan yemlerinde kullanılan Mn kaynakları genellikle inorganik Mn tuzları özellikle de sülfat formundadır. Bir çok araştırma organik mineral kaynaklarının kullanılabilirliğinin inorganik mineral kaynaklarından daha fazla olduğunu ve bunun kompleks ve şelat organik mineral kaynaklarının ileumdan absorbsiyonunun daha yüksek olmasından kaynaklandığını bildirmişlerdir (Henry ve ark 1989, Ji ve ark. 2006). Organik ligandlar farklı organik komplekslerden oluşur. Çoğu ticari ligandlar, amino asit, çoklu amino asit, hidroliz protein ve organik asitlerden oluşur. Çeşitli ligandlardan oluşan organik kompleksler kanatlılardaki absorbsiyonlarını etkileyen (stabilitesi, bağ etkinliği, fonksiyonel grup, molekül ağırlığı ve molekül boyutu gibi) farklı yapıdadırlar. İncebağırsaktan minerallerin alınımını azaltan fizikokimyasal
faktörler mineralin biyolojik kullanımını etkiler (Dreosti 1993). İnorganik mineraller sindirim sıvısında diğer rasyon unsurlarıyla interaksiyona girerek absorbsiyonu azaltan veya engelleyen kompleksler oluştururlar. Organik iz mineral şelatlarının sindirim kanalında şelat yapısını koruduğu için istenmeyen reaksiyonlarının az olduğuna inanılmaktadır (Ammerman ve ark . 1998). Ancak, organik ve inorganik Mn kaynaklarının kullanılabilirliği arasında fark olmadığını bildiren çalışmalar da mevcuttur (Baker ve Halpin 1987; Scheideler 1991). Smith ve ark. (1995) broylerlerde organik ve inorganik Mn kaynakları arasında farklılığın olmadığını, sıcaklık stresi altında yem tüketiminin azalması durumunda organik Mn kaynaklarının daha etkili olabileceğini bildirmişlerdir. Diğer bir çalışmada ise MnSO4 ve Mn proteinatın CAA, YT ve YDK’nı (Baker ve Halpin 1987), MnCO3 ve Mn metiyonin yemlenen broylerlerde performansı etkilemediğini bildirmişlerdir.
Li ve ark (2004), 546 adet günlük yaşta broyler civcivleri kullanarak ve Mn kaynağının nisbi biyolojik kullanılabilirliğini tespit etmek amacıyla bir çalışma yapmışlardır. Çalışmada, 5 adet Mn metiyonin kompleksi ( A,B,C,D,E), 5 adet Mn amino asit ( A, B,C, D, E), 2 adet Mn proteinat ( A, B), 1 adet Mn sülfat kullanılmıştır. Mangan seviyesi 60,120,180 mg/kg olarak katılmıştır. Rasyonda Mn seviyesinin artışına bağlı olarak kemik ve kalp Mn konsantrasyonu ile Kalp Mn superoksid dismutase mRNA seviyeside artmıştır. Mangan kaynağı, ilave edilen Mn seviyesi ve interaksiyonlarda günlük YT, CAA, YDK önemli derecede etkilememiştir.
İşcan ve Güçlü (2004), 128 adet 62 haftalık Nick chick yumurta tavuğu kullanılarak bir çalışma yapmışlardır. Dört grup oluşturulmuş ve rasyon kontrol grubu % 3.5 Ca, deneme grupları sırasıyla % 4 Ca, % 3.5 Ca + 1g/kg eggshell 49 ve % 4 Ca+1g/kg eggshell 49 içeren mısır-soya esaslı bazal rasyonla beslenmiştir. Organik Mn ve Zn kaynağı olarak 4500 ppm Mn ve 7500 ppm Zn içeren eggshell 49 kullanılmıştır. Yapılan bu araştırma sonunda ortalama CA, YV, YT, YDK, yumurta ağırlığı bakımından gruplar arasında istatistik açıdan önemli bir farklılık görülmemiştir. Serum Ca, Mg, P değerleri arasındaki farklılık önemli bulunmamıştır.
Li ve ark (2005), günlük yaşta 624 adet broyler civcivi kullanarak yaptıkları çalışma da, yüksek Ca’ lu rasyona ( kontrol rasyonu 18.5 g/kg Ca+ 20 mg/kg Mn) 60,120,180 mg/ kg seviyesinde Mn sülfat (MnSO4H2O) , Mn metiyonin, Mn aminoasit B ve Mn aminoasit C katılmıştır. Rasyonda Mn seviyesi arttıkça kalp Mn konsantrasyonu ve Mn içeren bir enzim olan superoksid dismutase enzimi mRNA seviyesi önemli derecede artmıştır. Farklı Mn kaynaklarının belirlenmesinde kalp Mn seviyesine nispetle kalp superoxid dismutase mRNA seviyesinin daha hassas bir kriter olduğu bulunmuştur. Superoksid dismutase kriter olarak alındığında 3 organik Mn kaynağının biyolojik kullanılabilirliği Mn sülfata nisbetle 1.12, 1.45, 1.80 olarak bulunmuştur. Normal Ca içeren rasyonla mukayese edildiğinde Mn sülfatın biyolojik değeri 1 olarak kabul edilirse, üç organik Mn kaynağının biyolojik kullanılabilirliği 0.99, 1.32 ve 1.13 olarak bulunmuştur. Bu sonuçlar, sindirim esnasında yüksek Ca rasyonun Mn absorbsiyonu üzerindeki olumsuz etkisinin Mn aminoasit B ve C ile kısmen veya tamamen ortadan kaldırdığını göstermiştir. Rasyonda kullanılan Mn kaynakları 1-21 günlük denemede civcivlerin günlük ortalama CAA, günlük ortalama YT ve YDK’sını önemli olarak etkilememiştir. Kaynak x seviye interaksiyonu günlük ortalama CAA, YDK, YT’ ni önemli olarak etkilememiştir. Mangan miktarı bacak kusurları ve kemik külünde önemli etkiye sahip olmamıştır. İlave Mn seviyesi ise CAA, YDK, YT ve bacak kusurlarını önemli olarak etkilemiştir. Mangan seviyesi kül Mn ve kalp MnSOD (superoksid dismutase) aktivitesini etkilemiş olup, Mn seviyesi arttıkça ikisinin miktarının da arttığı ifade edilmiştir.
Lu ve ark (2005), 288 adet günlük yaşta Arbor Acres hattı erkek broyler civciv kullanarak yaptıkları çalışmada, rasyona manganez ilavesinin karkas özellikleri, et kalitesi ve karın yağı ile etteki enzim aktivitesine etkisini araştırmışlardır. Civcivler 6 muamele grubunda 8 tekerrürlü ve her alt grupta 6 civciv olacak şekilde rastgele gruplara dağıtılmışlardır. Civcivler 0-3 hafta ilave Mn içermeyen mısır-soya küspesi ağırlıklı 9.5 g Ca/kg ve 22.74 mg Mn/kg içeren rasyon ile, 4-6 haftalarda ise 8.8 g Ca/kg ve 18.86 mg Mn/kg içeren rasyonlar beslenmişler. Aynı zamanda basal rasyon olarak kullanılan bu rasyonlara Mn sülfat formunda 100, 200, 300, 400 ve 500 mg/kg Mn ilave edilerek deneme rasyonları hazırlanmış ve bu
rasyonlar 42 gün boyunca yedirilmiştir. İlave Mn seviyesi, karkas randımanı, but ve göğüs miktarına, su tutma kapasitesine, kesme gücüne ve göğüs ve bacak kaslarındaki kas içi yağlara etkisi olmamıştır. Ayrıca, ilave Mn seviyesi; bacak kaslarındaki ph değerlerini, MDA (malondialdehyde) miktarını, göğüs kasındaki Mn içeren süperoksit dismutaz (MnSOD) aktivitesini, karın yağlarındaki MDH ve HSL aktivitelerini etkilememiştir. Çalışmadan elde edilen sonuçlar; broyler rasyonlarına 100 mg Mn/kg ilavesinin karın yağındaki LPL aktivitesinin azalmasına bağlı olarak karın yağı miktarını azaltabileceğini göstermiştir.
Ji ve ark (2006), piliçlerin ince bağırsağın farklı bölgelerinde (duedonum, jejenum ve ileum) Mn emilimi üzerine Ca ve Mn kaynaklarının etkilerini araştırmak için deneme yürütmüşlerdir. Deneme sununda elde edilen verilere göre etlik piliçler için ileumum Mn absorpsiyonu için önemli bir bölge olduğunu ve organik Mn kaynaklarının inorganiklere göre daha fazla miktarda emildiğini göstermiştir.
Bao ve ark (2007), etlik piliç rasyonlarına organik element( Cu, Fe, Mn, Zn) kaynaklarını ilave ederek bir çalışma yapmışlardır. Organik elementlerin inorganiklere göre daha yüksek seviyelerde yarayışlı olduğunu ifade etmişlerdir. Sonuç olarak, etlik piliç rasyonlarına 4 mg Cu, 40 mg Fe,40 mg Mn ve 40 mg Zn organik element ilavesinin 29 gün boyunca yeterli olduğunu ve bu sayede dışkı ile atılan element miktarının azaltılabileceğini ifade etmişlerdir.
3. MATERYAL METOT
Bu araştırma Selçuk Üniversitesi, Ziraat Fakültesi Zootekni Bölümü Prof. Dr. Orhan Düzgüneş Hayvancılık Araştırma ve Uygulama Çiftliği Bıldırcın ünitesinde yürütülmüştür. Deneme 5 hafta (35 gün) sürmüştür. Deneme hayvanları kuluçka makinesinden çıkarıldıkları tarihten itibaren 35 gün boyunca elektrikle ısıtılan apartman tipi büyütme kafeslerinde barındırılmıştır. Civcivler bu süre zarfında mısır – soya küspesine dayalı NRC (1994) tarafından tavsiye edilen besin maddeleri baz alınarak hazırlanan deneme rasyonları ile yemlenmişlerdir.
Deneme hayvanları 5 haftalık süre içerisinde katlı kafeslerde her katında 2 göz bulunan ve her gözünde 10 adet bıldırcın olacak şekilde yetiştirilmiştir. Bıldırcınların ve deneme rasyonlarının (muamelelerin) kafes gözlerine dağıtımı şansa bağlı olarak yapılmıştır. Araştırma boyunca bıldırcınlara yem ve su ad-libitum olarak verilmiş ve 23 saat aydınlatma programı uygulanmıştır.
3.1. Materyal
3.1.1 Hayvan materyali
Araştırmada toplam 360 adet karışık cinsiyette Japon bıldırcını kullanılmıştır. Araştırma 5 hafta sürmüştür. Araştırma boyunca gruplara ait canlı ağırlıklar her hafta aynı saatte grup tartımları şeklinde tartımları yapılarak tespit edilmiştir. Aynı şekilde
her hafta grupların tükettikleri yemden arta kalan yem tartılarak, grupların haftalık yem tüketimleri belirlenmiştir.
3.1.2.Yem materyali
Deneme rasyonlarının hazırlanmasında kullanılan yem hammaddeleri ticari bir yem fabrikasından satın alınmıştır. Satın alınan dane yem ve küspeler 5 mm’lik eleğe sahip, yerli imalat çekiçli bir değirmende kırıldıktan sonra ana rasyonlar hazırlanmıştır.
3.2.Metot
Aşağıda deneme rasyonlarının hazırlanması, deneme gruplarının oluşturulması, verilerin toplanması ve sonuçların değerlendirilmesi kullanılan metotlar hakkında bilgi verilmiştir.
3.2.1. Deneme rasyonlarının hazırlanması ve grupların oluşturulması
Otuz beş gün süren denemede toplam 360 adet günlük Japon bıldırcını kullanılmıştır. Araştırma, her alt grupta 10 civciv olmak üzere üç tekerrürlü olarak (36x10=360 hayvan) yapılmıştır.
Denemede % 24 ham protein ve 2900 kcal/kg ME, % 0.50 metiyonin, % 1.30 lisin, % 0.8 Ca ve 60 ppm Mn içeren rasyon kontrol rasyonu olarak kullanılmıştır (NRC 1994). Deneme rasyonları % 0.4, 0.8, 1.6 ve 2.4 Ca ve 0, 60, 120 mg/kg Mn içerecek şekilde (4x3 = 12) tesadüf parselleri deneme planına göre yürütülmüştür.
Dört farklı Ca seviyesi ile her birine 3 farklı seviyede Mn ilavesinden oluşan toplam 12 rasyonun etkisi denenmiştir. Araştırma 3 tekerrürlü olarak yürütülmüş (12 x 3 = 36 alt grup ) olup, her bir tekerrüre rastgele hayvanlar yerleştirilmiştir.
Rasyonda arzu edilen seviyede Mn miktarını sağlamak için denemede denenen Mn miktarları tartılıp iz mineral karması, metiyonin, vitamin premiksi gibi materyallerle birlikte plastik kova içinde 3 kg mısır ile karıştırıldıktan sonra diğer materyallerle karıştırılmış ve daha sonra tüm materyaller mikserde karıştırılmıştır.
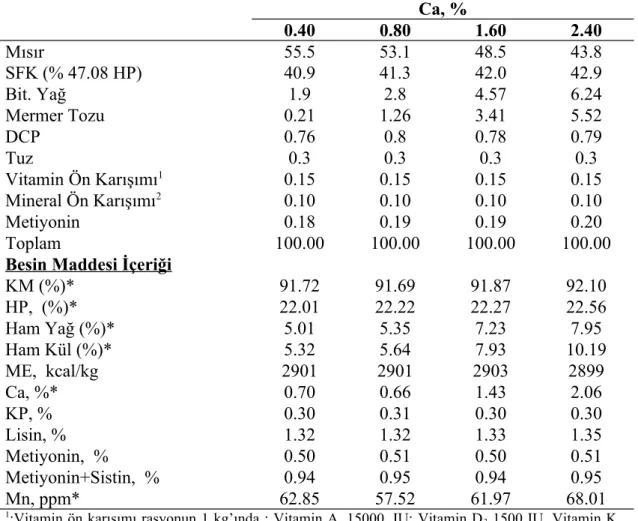
Çizelge 3.1. Araştırmada kullanılan ana rasyonların hammadde ve hesaplanmış besin
maddeleri kompozisyonu Ca, % 0.40 0.80 1.60 2.40 Mısır 55.5 53.1 48.5 43.8 SFK (% 47.08 HP) 40.9 41.3 42.0 42.9 Bit. Yağ 1.9 2.8 4.57 6.24 Mermer Tozu 0.21 1.26 3.41 5.52 DCP 0.76 0.8 0.78 0.79 Tuz 0.3 0.3 0.3 0.3 Vitamin Ön Karışımı1 0.15 0.15 0.15 0.15 Mineral Ön Karışımı2 0.10 0.10 0.10 0.10 Metiyonin 0.18 0.19 0.19 0.20 Toplam 100.00 100.00 100.00 100.00
Besin Maddesi İçeriği
KM (%)* 91.72 91.69 91.87 92.10 HP, (%)* 22.01 22.22 22.27 22.56 Ham Yağ (%)* 5.01 5.35 7.23 7.95 Ham Kül (%)* 5.32 5.64 7.93 10.19 ME, kcal/kg 2901 2901 2903 2899 Ca, %* 0.70 0.66 1.43 2.06 KP, % 0.30 0.31 0.30 0.30 Lisin, % 1.32 1.32 1.33 1.35 Metiyonin, % 0.50 0.51 0.50 0.51 Metiyonin+Sistin, % 0.94 0.95 0.94 0.95 Mn, ppm* 62.85 57.52 61.97 68.01
1:Vitamin ön karışımı rasyonun 1 kg’ında : Vitamin A, 15000, IU; Vitamin D
3 1500 IU, Vitamin K,
5.0 mg; Vitamin B1, 3mg; Vitamin B2, 6 mg; Vitamin B6.5 mg; Vitamin B 12, 0,03 mg; Niasin, 30 mg; Biotin, 0.1 mg Kalsiyum D-pantotenat, 12.0 mg ; Folik asit, 1.0 mg; Kolin klorid, 400 mg temin eder.
2:Mineral ön karışımı rasyonun 1 kg ‘ında: Manganez, 80 mg; Demir,35 mg; Çinko,50mg; Bakır,5.0
mg; İyod, 2mg; Kobalt, 0.04 mg temin eder. *Analiz yapılarak bulunmuş değerler.
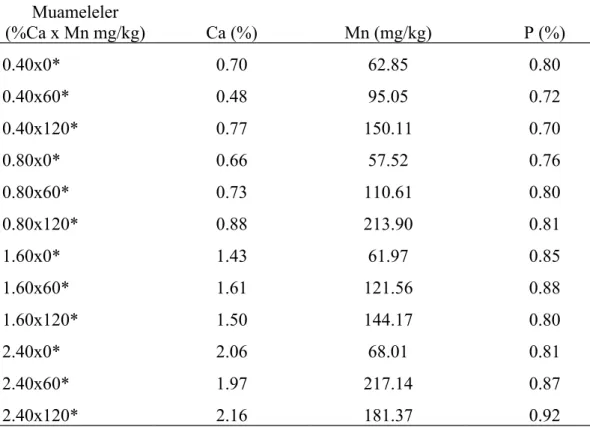
Çizelge 3.2. Araştırmada kullanılan rasyonların analiz edilmiş mineral madde içerikleri Muameleler (%Ca x Mn mg/kg) Ca (%) Mn (mg/kg) P (%) 0.40x0* 0.70 62.85 0.80 0.40x60* 0.48 95.05 0.72 0.40x120* 0.77 150.11 0.70 0.80x0* 0.66 57.52 0.76 0.80x60* 0.73 110.61 0.80 0.80x120* 0.88 213.90 0.81 1.60x0* 1.43 61.97 0.85 1.60x60* 1.61 121.56 0.88 1.60x120* 1.50 144.17 0.80 2.40x0* 2.06 68.01 0.81 2.40x60* 1.97 217.14 0.87 2.40x120* 2.16 181.37 0.92
* Yem laboratuarında yapılan analiz sonucu bulunan yemdeki mineral değerleri.
3.2.2. Denemenin yürütülmesi
3.2.2.1. Performansla ilgili kriterlerin tespiti
Deneme gruplarındaki Japon bıldırcınlarının yem tüketimleri ile canlı ağırlıkları denemenin başladığı tarihten itibaren aynı saatlerde yapılan haftalık tartımlarla saptanmıştır. Her tartımdan önce alt gruplardaki yemlikler ait oldukları kovalarına boşaltılarak kovalardaki artan yemler tartılmış, buna göre alt grupların haftalık ve tüm deneme dönemindeki ortalama yem tüketimleri hesaplanmıştır. Her bir alt gruptaki hayvanlar grup olarak plastik kaplara konularak haftalık tartımları yapılmış ve daha sonra haftalar itibariyle alt gruplara ait canlı ağırlık artışları hesaplanmıştır. Denemenin başından itibaren ölümler günlük olarak kaydedilmiş ve alt grupların yem tüketimleri hesaplanırken bu kayıtlar dikkate alınmıştır. Grupların gerek haftalık gerekse kümülatif yem değerlendirme katsayıları (YDK), denemenin
sonunda gruplara ait haftalık canlı ağırlık artışı (CAA) ve yem tüketimi (YT) ortalamalarına ait değerlerden yararlanılarak hesap edilmiştir.
Kan analizleri için her alt gruptan rastgele 2 Japon bıldırcınından plazma Ca ve Mn konsantrasyonunu tespit için kan numunesi alınmıştır. Kan numunesi alınan hayvanlar kesilip iç organları çıkarıldıktan sonra karkas kriterlerini değerlendirmek maksadıyla sıcak karkas ağırlıkları tespit edilmiştir. Tibia analizleri için her alt gruptan 2 adet kesilen bıldırcınların (1 erkek ve 1 dişi, toplam 72 adet) tibiaları alınmış ve torbalara konulmuştur.
3.2.2.2. Kan numunelerinin alınış tekniği
Deneme hayvanlarının son tartımları yapıldıktan sonra her alt gruptan rastgele 1 dişi ve 1 erkek bıldırcın olmak üzere 2 adet hayvan alınarak toplamda 72 adet hayvanın plazma Ca ve Mn seviyesini tespit etmek için enjektör yardımıyla kan alınmıştır. Alınan kanlar heparinli tüplere boşaltılmıştır. Takiben kan numuneleri 2500 devirde 7.5 dakika santrifüj edilerek plazma ayrılmıştır. Ayrılan plazmalar tüplere konularak hangi gruplara ait olduklarını belirtmek için numaraları yazıldıktan sonra analiz yapılıncaya kadar -20 0C derin dondurucuda muhafaza edilmiştir.
3.2.2.3. Plazma kalsiyum ve manganez konsantrasyonunun tespiti
Plazma Ca ve Mn konsantrasyonunun tespitinde ICP cihazı kullanılmıştır. Plazma Ca ve Mn konsantrasyonlarını tespit etmek maksadıyla yaş yakma metodundan yararlanılmıştır(Wedekind ve ark., 1992). Bu maksatla daha önceden hazırlanan plazmalardan 0.5 ml alınarak % 98 saflıktaki sülfirik asitle yakılıp saf su ile 10 ml’ye tamamlandıktan sonra ICP’ de okuma değerleri tespit edilmiştir. Daha sonra Bayraklı ( 1987)’nın bildirdiği metoda göre plazma Ca ve Mn seviyeleri bulunmuştur.
Denemenin sonunda alınan tibialardan Ca ve Mn seviyesini tespit etmek maksadıyla kül fırınında 550 OC de 20 saat yakılmıştır. Yakma işleminden sonra
tibiaların külleri tartılmıştır. Yakma işleminden sonra kalan küllerin üstüne 5 ml 6 N HCl ilave edilip mavi bantlı filtre kağıdında süzülmüştür. Bu işlemlerden sonra materyal 50 ml’ye saf suya tamamlanmıştır. Daha sonra ICP cihazında okunarak tibia külünde Ca ve Mn seviyesi tespit edilmiştir (Seal ve Weeth, 1980).
3.2.2.5. Yemdeki kalsiyum ve manganez konsantrasyonunun tespiti
Yemdeki Ca ve Mn konsantrasyonunun tespitinde ICP cihazı kullanılmıştır. Yemdeki Ca ve Mn konsantrasyonunun tespitinde yaş yakma metodundan yararlanılmıştır (Wedekind ve ark. 1992). Bu maksatla denemenin son günü hayvanlara yedirilen yemlerden numune alınmış ve üzerinde numaraları yazılı naylon torbalara ayrı ayrı konulmuştur. Daha sonra organik bulaşmaları önlemek için asitli sudan geçirilerek kurutulmuş olan cam kaplara konulmuştur. Cam kaplara konan yem örnekleri 105 OC 24 saat süreyle etüvde tutularak kurutulmuşlardır. Kurutulan
yem örnekleri öğütülüp nem kapmaması için kilitli naylon torbalara konulmuştur. Yaklaşık 0.5 g yem numunesi alınarak % 98’lik sülfirik asitle yakıldıktan sonra saf su ile 50 ml’ ye tamamlanıp ICP de okuma değeri tespit edilmiştir. Daha sonra Bayraklı (1987)’nin bildirdiği metoda göre yemlerin Ca ve Mn konsantrasyonu tespit edilmiştir.
3.2.3. İstatistik metotlar
Araştırmada 4 farklı Ca (% 0.4, 0.8, 1.6 ve 2.4) ve 3 farklı seviyede Mn (0, 60 ve 120) seviyesinin oluşturduğu toplam 12 muamele tesadüf parsellerinde, faktöriyel deneme planında ve 3 tekerrürlü olarak denendiği için veriler faktöriyel deneme planına göre analiz edilmiştir. Muamelelerin incelenen parametreleri önemli olarak etkileyip etkilemedikleri MINITAB istatistik paket programı kullanılarak variyans analizi ile test edilmiş ve F değerlerinin önemli bulunduğu durumlarda, farklı ortalamaların tespiti Duncan testi ile yapılmıştır ( Düzgüneş 1975 ). Denemenin
matematik modeli aşağıdaki gibi olup, modelde rasyon Ca ve rasyon Mn seviyesi ana etkiler olarak ve bunların oluşturduğu muamele kombinasyonları interaksiyon etkileri olarak yer almıştır.
Denemenin matematik modeli ; Y ijk=µ+ai+bj+(ab)ij+eijk
Yijk=i’inci Ca ve j’inci Organik Mn seviyesindeki k’ıncı grubun incelenen özelliği
µ: Genel ortalama
ai: i. Ca seviyesinin etkisi bj: j. Mn seviyesinin etkisi (ab) ij : interaksiyon etkisi eijk: Hata veya tesadüfî etkiler