Reconstitution of RNA Polymerase II subunits and analysis of their interaction with PIC components
Tam metin
Şekil
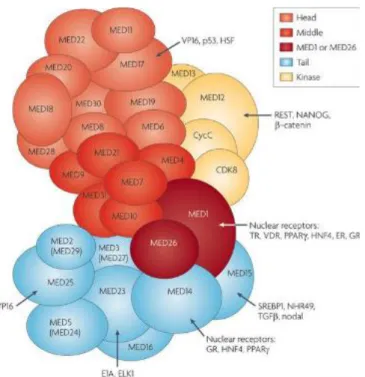
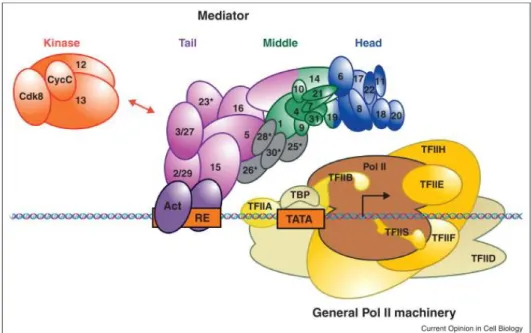
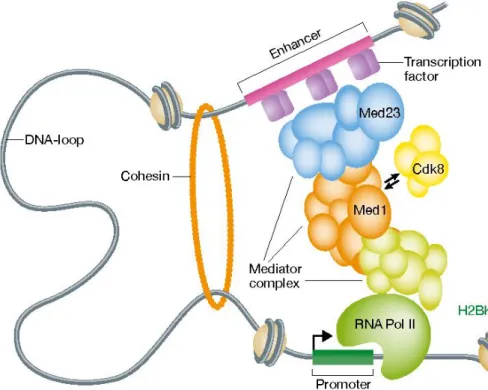
![Figure 1.3 Representation of predicted Pol II- Mediator Complex interaction. [34]](https://thumb-eu.123doks.com/thumbv2/9libnet/5987768.125662/23.892.335.582.759.994/figure-representation-predicted-pol-ii-mediator-complex-interaction.webp)

Benzer Belgeler
Last, although it is hardly ever possible to predict exactly the time of crises, CAMELS has a quite successful performance to analy- ze the risks of
Farklı özellik olarak; Triangular ve Asimile tip insidansı Hindistan'da bizim çalışmamızdan yüksek, Kesik ve Nodüler tip insidansları ise düşük çıkmıştır. Bunun
Bandyopadhy gibi geometriciler Chaki pseudo simetrik manifold kavramı yardımyla pseudo-projektif Ricci simetrik manifold ve zayıf simetrik manifold tanımlarını verip bu
The results of this experimental proof-of-concept study show that it is possible to accurately track multiple targets using a UWB radar sensor network in indoor environments based
Sonuç olarak antioksidan, antiinflamatuvar, antiviral, antimutajen, antibakteriyel, antialerjik, hepatoprotektif ve nöroprotektif etkileri gösterilmiş olan
Amaç: Yoğun bakım ünitesinde (YBÜ) sepsis tanılı hastalarda sıvı resüsitasyonuna yanıtın değerlendirilmesi için kullanılan santral venöz basınç (SVB), arteryel
Odak figür için doğum günü hediyesi olan sarı kır çiçeği, bir başkasının hüznünü anlatmıştır ve Nermin’i hayatın gerçekleriyle yüzleştirmiştir,
By using a parallel plate waveguide that incorporated an upper metal grid, images of the propagating field in the presence of the bare metal cylinder and cloaked cylinder were
