Elektroensefalografik fokal epileptik deşarjlar ve jeneralizasyon belirteçleri
Tam metin
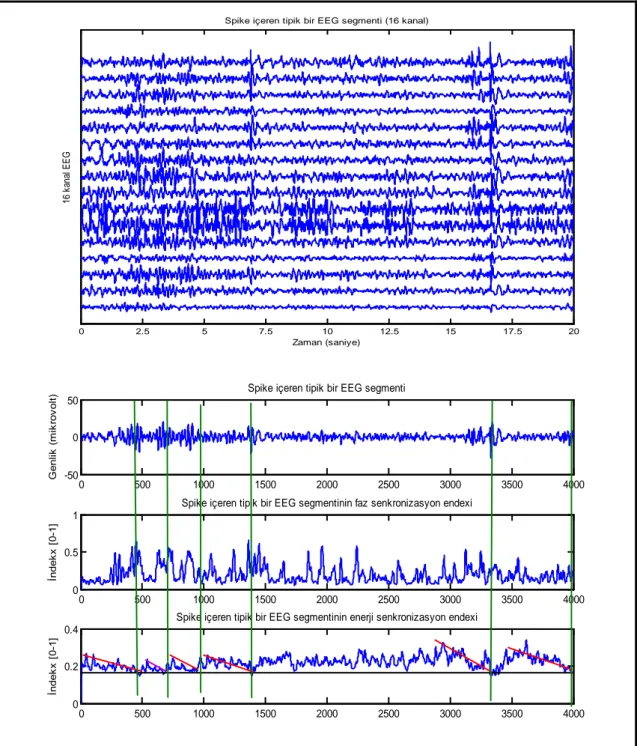
Şekil
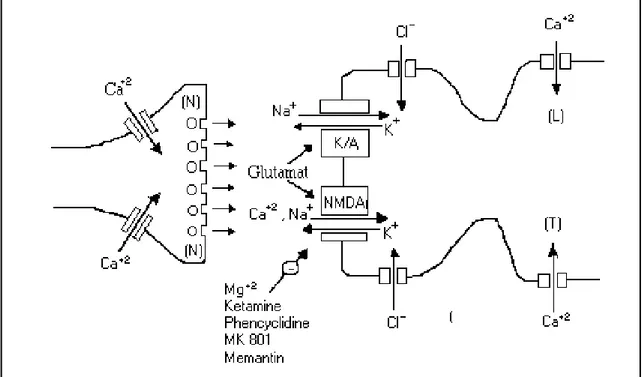
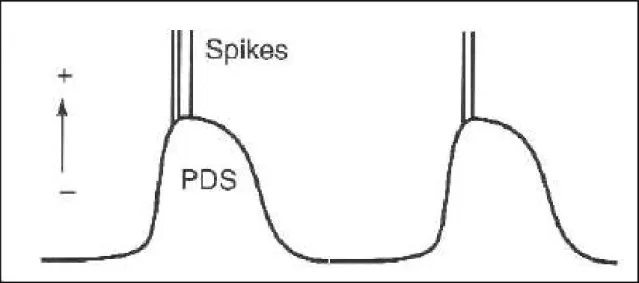
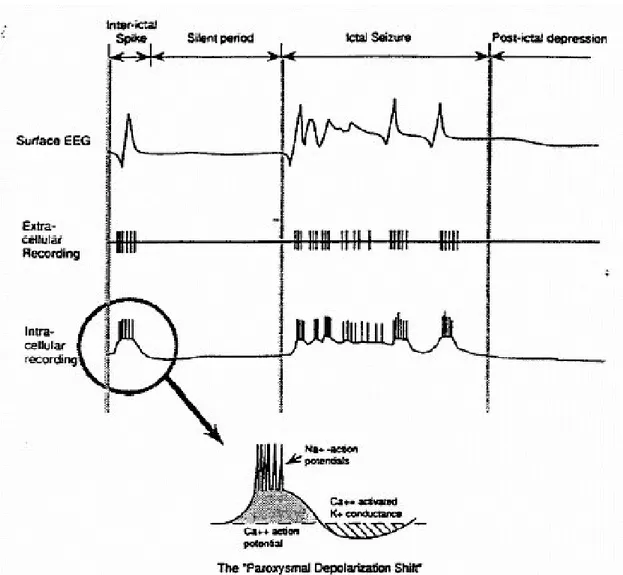
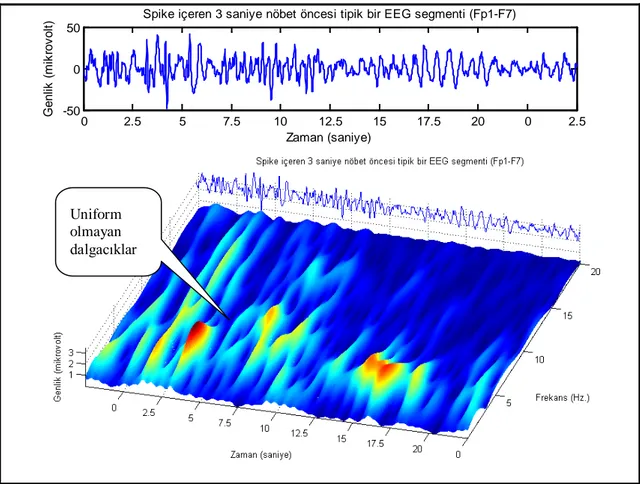

Outline
Benzer Belgeler
Botu- linum enjeksiyonu sonras›nda gerek minor niflasta testinin kalitatif skorlar›nda, gerekse de hastalar›n subjektif yak›nma skor- lar›nda istatistiksel olarak anlaml›
(Salah & Amoura, 2016) "The language of electronic disclosure XBRL and its role in improving the efficiency of financial markets" touches on the importance of the role
In this paper, we summarized the most common network structures used in the processing of medical images and provided a glimpse of the most important techniques
BBT’de kalsifik lezyon görüldüğünde kalsifikasyon gösteren tümörlerin de ayırıcı tanısını yapmak amacıyla MRG ve fokal kortikal lezyonların her zaman
Sekonder jeneralize (fokal başlangıçtan bilateral tonik klo- nik nöbete dönüşen) olan ve olmayan grup karşılaştırıldı- ğında ise epilepsi süresi, nöbet sıklığı,
Gelişimsel venöz anomaliler veya diğer adıyla venöz malformasyonlar serebral vasküler malformasyonların en sık görüleni olup genellikle manyetik rezonans
Fokal adezyonlar, hücre iskeletinin aktin filamentleri, sitozolik proteinler, plazma membran proteinleri ve ekstraselüler bileşenleri de içine alan makromoleküler
Olgumuzda 1.5 Tesla kranial MRG ile T2, FLAIR ve difüzyon a¤›rl›kl› görüntülerin elde edildi¤i protokol uygulanm›fl ve s CJH için tipik olarak kabul edilen bilateral
