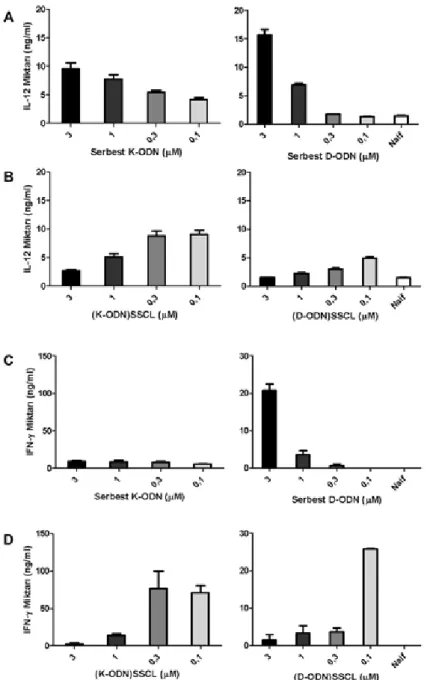
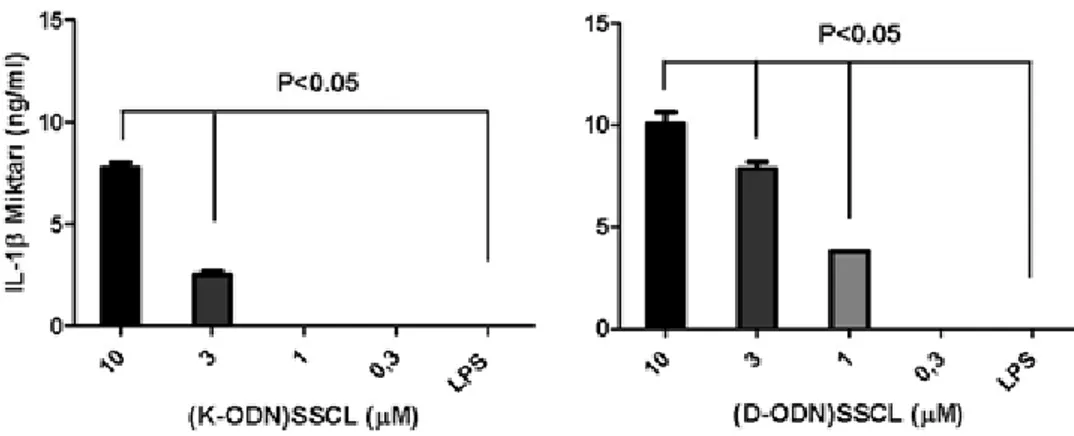
Inflammasome induction and immunostimulatory effects of CpG-ODN loaded liposomes containing DC-cholesterol
Tam metin
Şekil


Benzer Belgeler
Therefore the model does not provide any distinction between the Chls a and the Chls b (the identity problem) and no distinction could be made between the molecular x and y axes of
Ratings for novel objects (stars) that inherited material properties from corresponding familiar objects tended to overlap with that of the familiar objects; however, this
combination of each acquisition creates biases in coil sensitivity estimates due to signal modulations from bSSFP sensitivity profiles, the performance of subsequent
The ABC C-code is translated from the QuickBasic dialect source code extended for easy use of mathematical routines such as complex numbers, ar- bitrary precision
Çizelge 32 incelendiğinde, Bisiklet festivallerindeki harcamaların cinsiyete göre farklılık gösterdiği görülmektedir (t=2,089; p= 0,038) Bu sonuçtan hareketle kadınların
If every label position in K overlaps at most one other label position in K, and each graph object has at least two label positions, then Algorithm Fast Matching produces a
We analyze the ef- fects of the market structure in the components market on R&D decisions of a durable good monopolist and find that if the monopolist engages in partial
İşlerinden düşük düzeyde tatmin olan annelerin iş stresi ortalama puanlarının orta ve iyi düzeyde işlerinden tatmin olan annelere göre önemli düzeyde daha düşük olduğu,
