https://doi.org/10.1007/s12210-019-00796-7
Effect of cadmium, chromium, and lead on micropropagation
and physio‑biochemical parameters of Bacopa monnieri (L.) Wettst.
cultured in vitro
Muhammet Dogan1
Received: 4 January 2019 / Accepted: 4 April 2019 / Published online: 12 April 2019 © Accademia Nazionale dei Lincei 2019
Abstract
Heavy metals are released into the environment as a result of anthropogenic activities such as melting metallic ores, industrial productions, power stations, and pesticides. Among these, cadmium (Cd), chromium (Cr) and lead (Pb) are the most common heavy metals in the environment and can cause serious toxic effects on both animals and plants. In this study, the effect of Cd, Cr, and Pb on the propagation of Bacopa monnieri (L.) Wettst. by tissue culture was investigated. In addition, various physio-biochemical parameters (photosynthetic pigments contents, protein contents, lipid peroxidation, and hydrogen per-oxide) and survival frequency of the plant were evaluated. The shoot tip explants of B. monnieri were cultured in Murashige and Skoog (MS) nutrient media supplemented with 0.50 mg/L 6-benzylaminopurine (BAP) and different concentrations of Cd, Cr, and Pb (0.25–1.00 mg/kg) for 10 weeks. 100% shoot regeneration was determined in MS medium containing 0.25, 0.50, and 0.75 mg/kg Cd, 0.25 mg/kg Cr and 0.25 and 0.50 mg/kg Pb. Mean number of shoots per explant and shoot lengths decreased significantly with increasing metal concentration. In Cd, Cr, and Pb applications, maximum number of shoots per explant was obtained as 30.52, 22.47, and 27.12 shoot/explant and the highest shoot lengths were determined as 4.16, 4.27, and 4.35 cm, respectively. As the concentration of the metal exposed to the explant increased, the photosynthetic pigment contents of the plant decreased. The negative impact of metals on photosynthetic pigment contents was found to increase in the order: Cd < Cr < Pb. Similarly, the protein content of plants also decreased due to the negative effect of metals. On the other hand, malondialdehyde (MDA) and hydrogen peroxide (H2O2) values elevated with increasing concentrations of heavy metals. The findings of this study carried out under in vitro conditions may shed light on researchers who want to investigate the effects of heavy metals on plants.
Keywords Acclimatization · Heavy metal · Shoot regeneration · Tissue culture · Toxicity
1 Introduction
The term “heavy metals” refers to any metallic element which has a relatively high density and is toxic even at low concentration. Heavy metals include lead (Pb), cadmium (Cd), nickel (Ni), cobalt (Co), iron (Fe), zinc (Zn), chro-mium (Cr), iron (Fe), arsenic (As), silver (Ag), and platinum group elements (Nagajyoti et al. 2010). In recent years, rapid urbanization and industrialization, as well as the increase
in the use of pesticides and fertilizers has caused the heavy metal content in soil and wetlands to increase. Heavy metal emissions from anthropogenic sources such as power sta-tions, metal industries, cement industries and urban traffic are increasing rapidly (Sarwar et al. 2017; Ogbonna et al.
2018). These heavy metals may remain in the environment for a long time and tend to accumulate in living systems. For these reasons, heavy metals are among the most dangerous environmental pollutants (Wang et al. 2014; Ahmad et al.
2016).
Heavy-metal stress causes severe morphological, meta-bolic, and physiological abnormalities such as blocking photosynthetic pigmentation mechanism of plants, lipid peroxidation, and protein degradation (Sytar et al. 2013; Maleki et al. 2017). Heavy-metal accumulation in plants prevents the biosynthesis of photosynthetic pigments and * Muhammet Dogan
1 Department of Biology, Kamil Ozdag Faculty of Science,
Karamanoglu Mehmetbey University, Yunus Emre Campus, 70100 Karaman, Turkey
causes a decrease in chlorophyll and carotenoid content. Heavy metals disrupt chloroplast functions and change the component of the photosynthetic electron transport chain (ETC) and thus photooxidative damage occurs (Sytar et al.
2013). On the other hand, reactive oxygen species (ROS) such as hydrogen peroxide (H2O2), superoxide free radicals (O2·–), hydroxyl free radicals (·OH), and singlet oxygen ( O∗2 ) are produced in cells with heavy metal toxicity. ROS may cause protein oxidation and modification, lipid peroxidation, cell structure, and membrane deterioration, and even pro-grammed cell death. The level of lipid peroxidation is widely used as an indicator of ROS-mediated damage (Anjum et al.
2015a; Maleki et al. 2017). Plants have antioxidant defense systems consisting of non-enzymatic and enzymatic com-ponents to resist heavy-metal toxicity and oxidative stress. Nonenzymic components of the antioxidative defense sys-tem contain ascorbate, glutathione, tocopherol, carotenoids, and phenolic compounds. The enzymatic components of the antioxidative defense system include superoxide dismutase, catalase, glutathione peroxidase, ascorbate peroxidase, glutathione S-transferase, glutathione reductase and dehy-droascorbate reductase (Sharma et al. 2012; Maleki et al.
2017). In addition, plants respond to heavy-metal stress by the synthesis of binding proteins such as metallothioneins or phytochelatins (Mejáre and Bülow 2001).
Generally, as a result of human activities, Cd, Cr, and Pb are spread over the environment. These metals show acute and chronic toxic effects on plants. As a result of Cd expo-sure, photosynthetic activities of plants, nutrient intake, and water intake decrease. In plants exposed to excessive levels of Cd, chlorosis, growth retardation, browning of the root tips, as well as plant deaths, are seen (Wójcik and Tukien-dorf 2004; Mohanpuria et al. 2007). Cr is the most persistent metal in the soil and highly toxic to living systems. Some of the signs of Cr toxicity in plants are the prevention of ger-mination, root growth, and seedling growth. It also causes physiological and biochemical changes such as chlorosis and necrosis (Singh et al. 2013). Pb is one of the most toxic metals in soil. It also has negative effects on the morphol-ogy, growth, and photosynthetic activity of plants. The dete-rioration of mineral uptake, decrease in enzyme activities, water imbalance and changes in membrane permeability are observed in plants exposed to high levels of Pb (Sharma and Dubey 2005).
Bacopa monnieri (L.) Wettst., commonly known as Brahmi, is a plant of Indian origin. It can grow in wetlands, swamps, and humid places (Sharma et al. 2010). B. monnieri is renowned for its medical importance. In general, ayurve-dic meayurve-dicine is used as a nerve tonic, diuretic, cardiotonic, and therapeutic agents against epilepsy, insomnia, asthma, and rheumatism. Studies have shown that the extracts of this plant have anti-depression and antioxidant activity (Uabun-dit et al. 2010). B. monnieri is classified as a drug that is
effective in minimizing mental stress, anxiety, and necrosis states and improving intelligence functions (Bhusari et al.
2013; Srivastava et al. 2017). In addition to its intelligence-enhancing function, it has been suggested to be useful in the treatment of cardiac, respiratory, and neuropharmaco-logical disorders such as insomnia, depression, psychosis, and epilepsy (Russo and Borrelli 2005). Antiinflammatory, antipyretic, analgesic, free radical scavenger, and antilipid peroxidative activity have been reported (Russo and Borrelli
2005; Naik et al. 2017).
Plants are exposed to biotic and abiotic stresses in their natural environment (Agır et al. 2017; Kılıç et al. 2018). These stress agents negatively affect the production and productivity of plants. The responses of plants to these stress agents differ. In vitro selection-based tissue culture techniques have been widely used recently to determine the stress states of plants. This technique can be carried out in small areas, short periods and under controlled conditions, and thus stress-tolerant variants can be determined (Rai et al.
2011). The responses of plants propagated by tissue culture under stress conditions such as salt (Summart et al. 2010; Rezende et al. 2017), drought (Safarnejad 2004; Matheka et al. 2008) and heavy metal (Leung 2017; Dogan et al.
2018) have been studied previously.
In this study, the effects of Cd, Cr, and Pb on in vitro propagation of B. monnieri were investigated. Besides, pho-tosynthetic pigments, protein contents, lipid peroxidation, and hydrogen peroxide changes in plants were evaluated. According to our researches, there is no study on the effects of heavy metals on the growth of B. monnieri under tissue culture conditions. Therefore, this study is the first report and will contribute significantly to the literature.
2 Materials and methods
2.1 Tissue culture conditions and experimental setup
Bacopa monnieri (L.) Wettst. (Plantaginaceae) was obtained from the aquarium store in Konya (Turkey). Surface sterili-zation of the plants was performed according to the proto-col applied by Karatas et al. (2013). The shoot tip explants isolated from sterile plants were incubated in a Murashige and Skoog (1962) (MS) nutrient medium (Fig. 1) containing 0.50 mg/L 6-benzylaminopurine and 0.25, 0.50, 0.75, and 1.00 mg/kg cadmium [Cd(NO3)·4H2O—Sigma], chromium [Cr(NO3)3·9H2O—Sigma] and lead [Pb(NO3)2—Sigma] for 10 weeks. In experiments, MS basal medium with vita-mins containing 3% sucrose (Duchefa) and 0.65% plant agar (Duchefa) were used.
Prior to the preparation of heavy-metal solutions, stock solutions were prepared at certain rates and other
concentrations were prepared by diluting from this stock solution. In the preparation of the solutions, inert, and suf-ficiently large glass materials were used. Ultrapure water was used in media preparation. The pH of the nutrient medium was adjusted to 5.7 ± 0.1 using 1 N NaOH and 1 N HCl and then sterilized under 1.2 atmospheric pressure at 120 °C for 20 min. In the experiments, the explants were cultured at a temperature of 24 ± 1 °C and 16 h of light photoperiod under white LED light.
Since adventitious roots were obtained from regener-ated shoots in the culture medium, rooting studies were not applied. The nutrient medium from the regenerated plants was removed with tap water without damaging the plants, then the plants were transferred to the beaker with water (400 mL) for acclimation to the external con-ditions. 3–4 cm high river sand was placed to beaker base and the temperature was set to 24 °C and lighting was performed at the 16-h photoperiod. In addition, 10% Hoagland’s nutrient medium (Hoagland and Arnon 1950) was added to the water medium. The survival rates of plants in acclimatization studies were recorded.
2.2 Analysis of photosynthetic pigment content Washed fresh leaf samples (100 mg) were homogenized with 5 mL of 80% acetone. The final volume of the extract was then completed with 10% (v/v) acetone and centri-fuged at 3000 rpm for 5 min. The absorbance values against acetone were read at 663 nm for chlorophyll a, 646 nm for chlorophyll b and at 470 nm for carotenoid in a spectrophotometer (Thermo Fisher Scientific, Mul-tiskan Go). Chlorophyll a, chlorophyll b and carotenoid calculations were performed according to Lichtenthaler and Wellburn (1983). Total chlorophyll content is given as chlorophyll a + b.
2.3 Protein content
Protein analysis was performed according to the method determined by Lowry et al. (1951) and protein content is given as mg/g fw. 0.5 g of fresh plant material was homog-enized in 5 mL 0.1 M phosphorus buffer (pH 7.4) and cen-trifuged at 12,000 rpm for 10 min. 0.3 mL of the superna-tant and 3 mL of alkaline solution were added and allowed to stand at room temperature for 15 min. Then, 0.3 mL of Folin–Ciocalteu reagent was added and left to stand at room temperature for 30 min and the absorbance was read at a wavelength of 750 nm. The same method for standard curve drawing was applied using bovine serum albumin solutions prepared at different concentrations.
2.4 Lipid peroxidation
Lipid peroxidation was determined by measuring the malon-dialdehyde (MDA) concentration according to the method described by Heath and Packer (1968). 0.5 g of plant sam-ples was homogenized in 5 mL of 0.1% trichloroacetic acid (TCA) followed by centrifugation at 10,000 rpm for 5 min. 0.5% thiobarbituric acid (TBA) prepared in 4 mL of 20% TCA was added to 1 mL of supernatant. The prepared mix-ture was heated at 95 °C for 30 min, then it was cooled in ice quickly. The cooling tubes were centrifuged at 10,000 rpm for 15 min and then the absorbance of the supernatants was recorded at 532 and 600 nm by the spectrophotometer (Thermo Fisher Scientific, Multiskan Go). Non-specific absorbance at 600 nm was subtracted from the absorbance at 532 nm and activity was calculated.
2.5 Hydrogen peroxide levels
0.5 g of fresh shoots was ground with liquid nitrogen and suspended in 1.5 mL of 100 mM potassium phosphate
Fig. 1 The shoot-tip explants of B. monnieri cultured in MS nutrient medium
buffer at pH 6.8. Then H2O2 content in tissues was measured according to (Gay and Gebicki 2000).
2.6 Statistical analysis
All trials were established according to the experimental design of random plots. The experimental setups were made for all heavy metals (Cd, Cr and Pb) and applied in Magenta GA7® containers with three replicates. The data were analyzed with SPSS 21 for Windows (IBM Corporation, Armonk, NY, USA) program and Duncan’s multiple range test (DMRT) were applied for Post Hoc tests. Pearson’s cor-relation coefficients for the parameters obtained after Cd, Cr, and Pb applications were performed. Data given in percent-ages were subjected to arcsine transformation (Snedecor and Cochran 1997) before statistical analysis.
3 Results
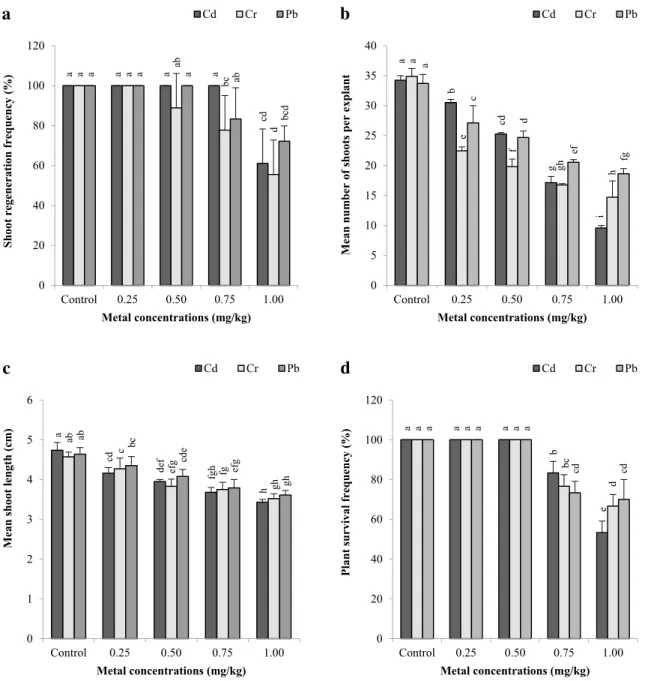
After 10 weeks, the shoot tip explants of B. monnieri were cultured in MS nutrient medium without heavy metal (Fig. 2a) and containing different concentrations of Cd (Fig. 2b), Cr (Fig. 2c) and Pb (Fig. 2d). Experiments have also been established in the culture medium which does not contain heavy metals. The percentage of shoot regeneration, the mean number of shoots per explant, mean shoot lengths of the explants under heavy metal effect were analyzed, and the acclimation rates of the plants grown in this culture medium were investigated (Fig. 3a–d).
The first shoot formation from explants in in vitro condi-tions was determined on the 10th day in the control group, on the 15th day in the Pb applications, on the 17th day in the Cr applications and on the 18th day in the Cd applica-tions. At the end of 5 weeks, multiple shoot regeneration was observed in all culture media and intensive adventitious root formations were recorded. At the end of 10 weeks, some physiological and morphological changes in plants were determined by the effects of heavy metals. Compared to the control group, the leaves of the plants exposed to Cd were smaller, and yellowish on the tips of the shoots, and local browning on the stem. In Cr applications, thickening of the stem, yellowing of leaves and a fragile branch structure were observed. In Pb application, browning of leaves on shoot tip, yellowing on body and leaves and softening of some shoots were determined.
3.1 Shoot regeneration frequency
The percentage of shoot regeneration in Cd, Cr, and Pb treated media was 100–61.11%, 100–55.55%, and 100–72.22%, respectively (Fig. 3a). 100% shoot regenera-tion was obtained in culture medium containing 0.25, 0.50,
and 0.75 mg/kg Cd, 0.25 mg/kg Cr and 0.25 and 0.50 mg/ kg Pb. In addition, 100% shoot occurrences were observed in heavy metal-free (control group) culture media. On the other hand, the lowest shoot regeneration frequencies were recorded in culture media using the highest concentrations of these three heavy metals. The results showed the adverse effects of heavy metals on shoot regeneration. In addition, according to the current results, Cr heavy metal showed the most negative effect for the regeneration of the shoot. It was determined that after Cd, Cr, and Pb applications a statistically positive correlation was recorded between shoot regeneration frequency and the number of shoots per explant [r = 0.699, p < 0.01 for Cd (Table 1); r = 0.658, p < 0.01 for Cr (Table 2); r = 0. 671, p < 0.01 for Pb (Table 3)].
3.2 Average number of shoots per explant
The shoot counts of B. monnieri from shoot tip explants were different with Cd, Cr, and Pb applications and these differences were statistically significant at p < 0.05 (Fig. 3b). The number of shoots per explant ranged from 34.24 to 9.58, 34.91 to 14.72, and 33.71 to 18.64 for Cd, Cr, and Pb, respectively. The highest number of shoots was determined in the control group in all three heavy metal applications. Significant reductions with the effects of heavy metals were observed in shoots from explants (p < 0.05). In culture media containing heavy metals, maximum shoots per explant were obtained in 0.25 mg/kg Cd, Cr, and Pb applications as 30.52, 22.47, and 27.12 shoots/explant, respectively. The increase in heavy metal concentration showed a decrease in the num-ber of shoots. The lowest shoot numnum-bers were determined as 9.58, 14.72, and 18.64 shoots/explant in MS culture medium containing 1.0 mg/kg Cd, Cr, and Pb, respectively. In heavy-metal applications up to 0.50 mg/kg, the highest number of shoots was determined in Cd applications, while the lowest number of shoots was obtained in 1.0 mg/kg Cd applications.
3.3 Average shoot length
The effects of heavy metal applications on the length of regenerated shoots were evaluated (Fig. 3c) and the data were statistically significant at 95% confidence interval (p < 0.05). The highest shoot lengths were obtained from the explants in the control group (heavy metal-free media), on the other hand, the highest shoot lengths in Cd, Cr, and Pb applications were determined as 4.16, 4.27, and 4.35 cm, respectively, in culture media containing 0.25 mg/kg. Gener-ally, the highest shoot lengths in all metal applications were obtained in Pb. This reveals that Pb has the least negative effect on shoot lengths. Shorter shoots were formed by an increase in concentrations of heavy metals. Furthermore, the negative effects of high concentrations of Cd were higher
than other metals. The mean shoot lengths in the 1.0 mg/kg Cd, Cr, and Pb applications were recorded as 3.43, 3.52, and 3.61 cm, respectively. All of these results showed that heavy
metals had inhibitory effects on prolongation of the shoot. According to the correlation matrices, there was a statisti-cally positive relationship between length of shoots and the
Fig. 2 In vitro shoot regenera-tion from shoot tip explants of B. monnieri in MS medium with Cd (b), Cr (c) and Pb (d) and without heavy metal (a) after 10 weeks of culture. The plants that are acclimated to the aquatic environment after 4 weeks (e)
number of shoots per explant shoot regeneration frequency and the number of shoots per explant [r = 0.929, p < 0.01 for Cd (Table 1); r = 0.887, p < 0.01 for Cr (Table 2); r = 0.932, p < 0.01 for Pb (Table 3)].
3.4 Acclimatization
Plants propagated under the influence of heavy metal were transferred into containers containing water for acclimatiza-tion (Fig. 2e). Plants acclimatized to 100% external condi-tions were determined in plants obtained in control group
plants and culture media containing 0.25 and 0.50 mg/kg heavy metal. The lowest acclimation values were 53.33% for Cd, 66.67% for Cr, and 70.00% for Pb (Fig. 3d). As it is understood from these results, the tolerance of acclima-tization to the external conditions of plants grown in MS medium with high concentrations of Cd is lower than that of other metals. Plants growing in heavy metal-containing media in tissue culture conditions can adversely affect the life cycle, even if transferred to a heavy metal-free medium because heavy metals have bioaccumulation and enter the structure in living beings. A significant correlation was
a a a a cd a a ab bc d a a a ab bc d 0 20 40 60 80 100 120 Control 0.25 0.50 0.75 1.00
Shoot regeneration frequency
(% ) Metal concentrations (mg/kg) Cd Cr Pb a b cd g i a e f gh h a c d ef fg 0 5 10 15 20 25 30 35 40 Control 0.25 0.50 0.75 1.00 Mean nu mb er
of shoots per explan
t Metal concentrations (mg/kg) Cd Cr Pb a cd de f fgh h ab c ef g fg gh ab bc cd e ef g gh 0 1 2 3 4 5 6 Control 0.25 0.50 0.75 1.00
Mean shoot length (c
m) Metal concentrations (mg/kg) Cd Cr Pb a a a b e a a a bc d a a a cd cd 0 20 40 60 80 100 120 Control 0.25 0.50 0.75 1.00 Plan ts urvival frequency (% ) Metal concentrations (mg/kg) Cd Cr Pb
a
c
d
b
Fig. 3 In vitro growth characteristics for shoot regeneration frequency
(a), mean number of shoots per explant (b) and mean shoot length (c) of shoot tip explants of B. monnieri treated with Cd, Cr, and Pb at dif-ferent concentrations after 10 weeks, and the survival frequencies of
in vitro-propagated plants after 4 weeks (d). All are mean values of triplicates ± SD (n = 3). Vertical bars indicate standard error of three separate experiments. Different superscript letters indicate signifi-cantly different values (p < 0.05; DMRT). Control = no heavy metal
Table 1 P earson ’s cor relation coefficients f or t he par ame ters obt ained af ter Cd applications *Cor relation is significant at t he 0.05 le vel (tw o-t ailed) **Cor relation is significant at t he 0.01 le vel (tw o-t ailed) Relations in Cd application Shoo t reg ener ation freq uency
The number of shoo
ts per
explant
Shoo
t lengt
h
Plant surviv
al freq uency Chlor oph yll a Chlor oph yll b To tal c hlor oph yll Car otenoid Pr otein M DA H2 O2 Shoo t r eg ener ation fr eq uency 1 0.699** 0.558* 0.817** 0.554* 0.578* 0.574* 0.664** 0.788** − 0.607* − 0.766**
The number of shoo
ts per explant 1 0.929** 0.907** 0.848** 0.931** 0.887** 0.907** 0.948** − 0.864** − 0.963** Shoo t lengt h 1 0.758** 0.875** 0.879** 0.902** 0.913** 0.822** − 0.844** − 0.866** Plant sur viv al fr eq uency 1 0.729** 0.806** 0.763** 0.818** 0.951** − 0.723** − 0.937** Chlor oph yll a 1 0.788** 0.995** 0.962** 0.777** − 0.755** − 0.855** Chlor oph yll b 1 0.846** 0.869** 0.886** − 0.895** − 0.862** To tal c hlor oph yll 1 0.975** 0.818** − 0.801** − 0.882** Car otenoid 1 0.877** − 0.805** − 0.912** Pr otein 1 − 0.778** − 0.964** M DA 1 0.779** H2 O2 1 Table 2 P earson ’s cor relation coefficients f or t he par ame ters obt ained af ter Cr applications *Cor relation is significant at t he 0.05 le vel (tw o-t ailed) **Cor relation is significant at t he 0.01 le vel (tw o-t ailed) Relations in Cr application Shoo t reg ener ation freq uency
The number of shoo
ts per
explant
Shoo
t lengt
h
Plant surviv
al freq uency Chlor oph yll a Chlor oph yll b To tal c hlor oph yll Car otenoid Pr otein M DA H2 O2 Shoo t r eg ener ation fr eq uency 1 0.658** 0.743** 0.672** 0.572* 0.676** 0.598* 0.658** 0.855** − 0.518* − 0.826**
The number of shoo
ts per explant 1 0.887** 0.651** 0.878** 0.798** 0.877** 0.843** 0.709** − 0.752** − 0.800** Shoo t lengt h 1 0.700** 0.813** 0.819** 0.825** 0.814** 0.817** − 0.712** − 0.869** Plant sur viv al fr eq uency 1 0.834** 0.893** 0.856** 0.837** 0.879** − 0.723** − 0.938** Chlor oph yll a 1 0.904** 0.997** 0.958** 0.765** − 0.811** − 0.877** Chlor oph yll b 1 0.933** 0.908** 0.842** − 0.800** − 0.918** To tal Chlor oph yll 1 0.963** 0.789** − 0.820** − 0.896** Car otenoid 1 0.782** − 0.770** − 0.903** Pr otein 1 − 0.711** − 0.937** M DA 1 0.728** H2 O2 1
recorded between plant survival frequency and the number of shoots per explant [r = 0.907, p < 0.01 for Cd (Table 1); r = 0.651, p < 0.01 for Cr (Table 2); r = 0.785, p < 0.01 for Pb (Table 3)].
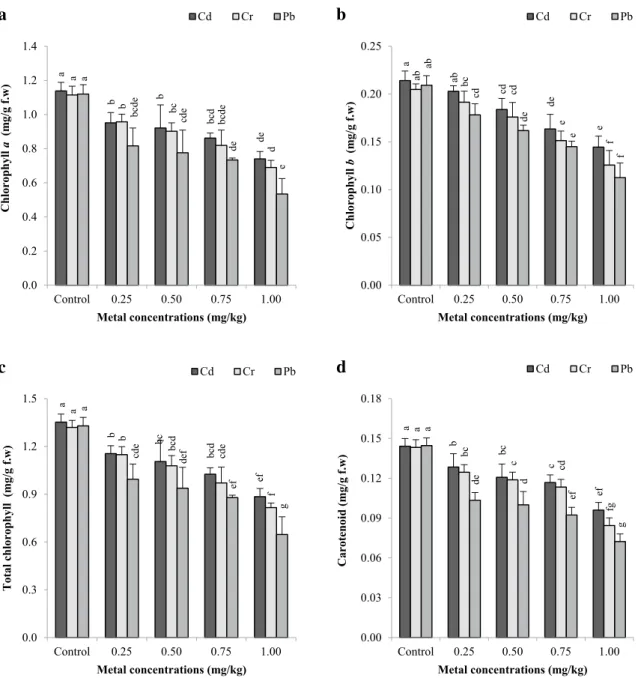
3.5 Photosynthetic pigment content
The photosynthetic pigment contents of B. monnieri, exposed to heavy metal stress, were analyzed (Fig. 4a–d). As the metal concentrations of plants increased, the photo-synthetic pigment contents of the plants decreased. With the effect of metals, yellowing and light green in the leaves of the plants were observed. Faster colour losses were observed especially in the upper leaves of the plant. There were sta-tistically significant decreases in chlorophyll a and b and total chlorophyll contents compared to the control group (p < 0.05). The most negative impact on photosynthetic pigment contents is Pb > Cr > Cd. The lowest chlorophyll a contents compared to the control were 35.11% for Cd, 37.86% for Cr, and 52.30% for Pb (Fig. 4a). As can be seen from this result, the high-dose application of Pb significantly decreased the content of chlorophyll a. The lowest chloro-phyll b contents were determined as 31.22% for Cd, 37.17% for Cr, and 46.40% for Pb (Fig. 4b) and the lowest total chlorophyll contents were obtained 34.60% for Cd, 38.18% for Cr, and 51.37% for Pb (Fig. 4c). According to the corre-lation matrices, there was a statistically positive recorre-lationship between the number of shoots per explant and chlorophyll a [r = 0.848, p < 0.01 for Cd (Table 1); r = 0.878, p < 0.01 for Cr (Table 2); r = 0.899, p < 0.01 for Pb (Table 3)], chlo-rophyll b [r = 0.931, p < 0.01 for Cd (Table 1); r = 0.798, p < 0.01 for Cr (Table 2); r = 0.896, p < 0.01 for Pb (Table 3)] and total chlorophyll [r = 0.887, p < 0.01 for Cd (Table 1); r = 0.877, p < 0.01 for Cr (Table 2); r = 0.914, p < 0.01 for Pb (Table 3)].
3.6 Carotenoid content
The change in carotenoid contents of plants under heavy metal effect is shown in Fig. 4d. Carotenoid contents decreased with increasing metal concentration. While the carotenoid contents of the plants have been observed in Cd and Cr applications, there has been a rapid decrease in carot-enoid level in Pb application (especially at 0.25 mg/kg). The highest decreases in carotenoid levels compared to the con-trol were recorded in the MS media containing 1.00 mg/kg metals as 33.43% for Cd, 41.00% for Cr and 49.80% for Pb applications, respectively. When the heavy-metal effects on carotenoid contents were compared, the most adverse effects were Pb > Cr > Cd. Significant correlations were determined between carotenoid values and the number of shoots per explant in Cd (r = 0.907, p < 0.01) (Table 1), Cr (r = 0.843, Table 3 P earson ’s cor relation coefficients f or t he par ame ters obt ained af ter Pb applications *Cor relation is significant at t he 0.05 le vel (tw o-t ailed) **Cor relation is significant at t he 0.01 le vel (tw o-t ailed) Relations in Pb application Shoo t reg ener ation freq uency
The number of shoo
ts per
explant
Shoo
t lengt
h
Plant surviv
al freq uency Chlor oph yll a Chlor oph yll b To tal c hlor oph yll Car otenoid Pr otein M DA H2 O2 Shoo t r eg ener ation fr eq uency 1 0.671** 0.730** 0.850** 0.571* 0.655** 0.593* 0.627* 0.761** − 0.780** − 0.804**
The number of shoo
ts per explant 1 0.932** 0.785** 0.899** 0.896** 0.914** 0.923** 0.802** − 0.921** − 0.878** Shoo t lengt h 1 0.803** 0.813** 0.828** 0.829** 0.834** 0.800** − 0.901** − 0.875** Plant sur viv al fr eq uency 1 0.669** 0.800** 0.700** 0.695** 0.897** − 0.766** − 0.892** Chlor oph yll a 1 0.865** 0.997** 0.971** 0.738** − 0.867** − 0.838** Chlor oph yll b 1 0.899** 0.893** 0.901** − 0.864** − 0.926** To tal Chlor oph yll 1 0.976** 0.774** − 0.881** − 0.865** Car otenoid 1 0.751** − 0.866** − 0.837** Pr otein 1 − 0.792** − 0.940** M DA 1 0.900** H2 O2 1
p < 0.01) (Table 2) and Pb (r = 0.923, p < 0.01) applications (Table 3).
3.7 Protein content
The protein contents of the plants exposed to Cd, Cr, and Pb were determined as 15.70–9.08, 15.69–12.53, and 15.06–11.82 mg/g fw, respectively (Fig. 5a). As with other parameters, the increase in heavy metal concentration applied to the plant caused a decrease in protein content. On the other hand, the protein contents of the lowest dose
of metal applied plants were increased compared to the control group (p < 0.05). In Cd, Cr, and Pb applications, the lowest protein contents were measured as 9.08 mg/g fw (41.40% lower than the control), 12.53 mg/g fw (20.08% lower than the control) and 11.82 mg/g fw (24.03% lower than the control), respectively. There was a statistically significant positive relationship between protein content and the number of shoots per explant in Cd (r = 0.948, p < 0.01) (Table 1), Cr (r = 0.709, p < 0.01) (Table 2) and Pb (r = 0.802, p < 0.01) applications (Table 3). a b b bc d de a b bc bcde d a bcde cde de e 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 Control 0.25 0.50 0.75 1.00 Chlorophyll a (m g/g f. w) Metal concentrations (mg/kg) Cd Cr Pb a ab cd de e ab bc cd e f ab cd de e f 0.00 0.05 0.10 0.15 0.20 0.25 Control 0.25 0.50 0.75 1.00 Chlorophyll b (m g/g f. w) Metal concentrations (mg/kg) Cd Cr Pb a b bc bc d ef a b bc d cd e f a cd e de f ef g 0.0 0.3 0.6 0.9 1.2 1.5 Control 0.25 0.50 0.75 1.00 Total chlorophyll (m g/g f.w ) Metal concentrations (mg/kg) Cd Cr Pb a b bc c ef a bc c cd fg a de d ef g 0.00 0.03 0.06 0.09 0.12 0.15 0.18 Control 0.25 0.50 0.75 1.00 Carotenoid (m g/g f. w) Metal concentrations (mg/kg) Cd Cr Pb
a
b
c
d
Fig. 4 Photosynthetic pigment values for chlorophyll a (a),
chloro-phyll b (b), and total chlorochloro-phyll (c) and carotenoid (d) contents of B. monnieri treated with Cd, Cr, and Pb at different concentrations after 10 weeks. All are mean values of triplicates ± SD (n = 3).
Verti-cal bars indicate standard error of three separate experiments. Differ-ent superscript letters indicate significantly differDiffer-ent values (p < 0.05; DMRT). Control = no heavy metal
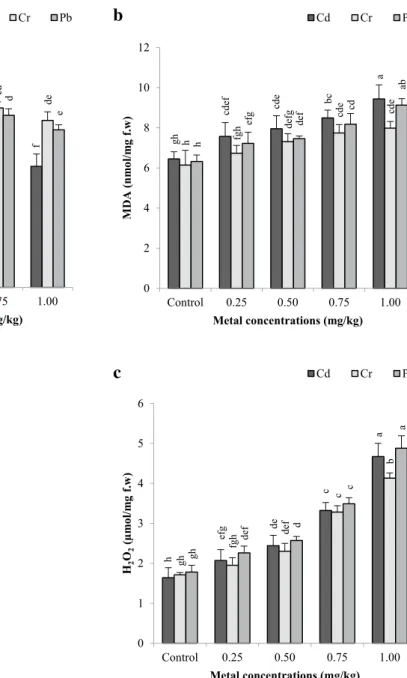
3.8 Lipid peroxidation (MDA content)
The data presented in Fig. 5b show the effects of differ-ent doses of Cd, Cr, and Pb on malondialdehyde (MDA) contents of B. monnieri, and MDA were measured between 6.45–9.44, 6.14–7.98, and 6.32–9.13 nmol/mg fw, respec-tively. Unlike other parameters, MDA contents increased with increasing concentration of metal concentrate (p < 0.05). As a result of spectrophotometric measure-ments, the highest MDA contents at 1.00 mg/kg Cd, Cr,
and Pb were determined as 9.44 nmol/mg fw (46.36% higher than that of the control), 7.98 nmol/mg fw (29.97% higher than that of the control) and 9.13 nmol/mg fw (44.46% higher than that of the control), respectively. Con-sidering the stress effects of metals on MDA, it is listed as Cd > Pb > Cr. According to the correlation matrices, there were a statistically negative relationship between MDA contents and the number of shoots per explant [r = − 0.864, p < 0.01 for Cd (Table 1); r = − 0.752, p < 0.01 for Cr (Table 2); r = − 0.921, p < 0.01 for Pb (Table 3)].
a a bc de f a a ab cd de a a bc d e 0 3 6 9 12 15 18 Control 0.25 0.50 0.75 1.00 Protein (m g/g f.w) Metal concentrations (mg/kg) Cd Cr Pb gh cdef cd e bc a h fgh defg cd e cde h ef g de f cd ab 0 2 4 6 8 10 12 Control 0.25 0.50 0.75 1.00 MDA (n mo l/mg f.w ) Metal concentrations (mg/kg) Cd Cr Pb h ef g de c a gh fgh def c b gh def d c a 0 1 2 3 4 5 6 Control 0.25 0.50 0.75 1.00 H2 O2 (µ mo l/m g f.w ) Metal concentrations (mg/kg) Cd Cr Pb
a
b
c
Fig. 5 Protein (a), malondialdehyde-MDA (b) and hydrogen
perox-ide–H2O2 (c) contents of B. monnieri treated with Cd, Cr and Pb at
different concentrations after 10 weeks. All are the mean values of
triplicates ± SD (n = 3). Vertical bars indicate standard error of three separate experiments. Different superscript letters indicate signifi-cantly different values (p < 0.05; DMRT). Control = no heavy metal
3.9 Hydrogen peroxide (H2O2)
The effects of different metal combinations on the H2O2 level of B. monnieri were shown in Fig. 5c. The H2O2 lev-els of shoots exposed to metal effect increased significantly (p < 0.05) and they ranged between 2.07–4.67, 1.95–4.13, and 2.26–4.88 µmol/mg for Cd, Cr and Pb applications, respectively. The highest H2O2 levels were recorded at 1.00 mg/kg Cd, Cr and Pb applications. While there was no statistically significant difference between Cd and Pb in 1.00 mg/kg metal applications, a statistically significant dif-ference was found in Cr application.
A statistically significant negative correlation was recorded between H2O2 values and the number of shoots per explant in Cd (r = − 0.963, p < 0.01) (Table 1), Cr (r = − 0.800, p < 0.01) (Table 2) and Pb (r = − 0.878, p < 0.01) applications (Table 3).
4 Discussion
Bacopa monnieri is a medically important plant (Kean et al.
2017; Smith et al. 2018) and is also a good heavy metal accumulator (Shukla et al. 2007). Therefore, tissue culture (Sharma et al. 2010; Kashyap et al. 2018; Srivastava et al.
2017; Dogan and Emsen 2018) and heavy metal (Mishra et al. 2013; Dogan et al. 2015; Jain et al. 2017) studies with B. monnieri have been carried out by many researchers recently. However, there is no detailed and comprehensive study of B. monnieri’s responses to heavy metal stress in tissue culture conditions. Therefore, in the present study, the effects of Cd, Cr, and Pb on in vitro production of B. monnieri and some biochemical changes in the plant were investigated.
The first regenerated shoots from shoot tip explants were quite late compared to the control group. This delay was determined as 8, 7, and 5 days for Cd, Cr, and Pb, respec-tively. 100% shoot occurrences were determined in low-metal culture media, while the percentage of shoot regen-eration in high metal-containing culture media decreased significantly. These results indicate that the formation and growth of shoots decreased in the presence of heavy metals. In addition, an inverse relationship was determined between the increase in metal concentration and the percentage of shoot regeneration. This may be due to the effect of Cd, Cr, and Pb in the regeneration pattern in cells (totipotency), oxidative stress, inhibition of enzyme activity, and inhibi-tion of photosynthesis activity (Chen et al. 2009; Gajewska et al. 2009; Yadav 2010; Amari et al. 2017). Similarly, Xu et al. (2009) reported a decrease in growth parameters with increased Cd concentration in Solanum nigrum L., which was cultured on tissue culture media with Cd.
In the present study, the number of shoots per explants under heavy metal effect was evaluated and they ranged from 34.24 to 9.58, 34.91 to 14.72, and 33.71 to 18.64 for Cd, Cr, and Pb, respectively. In contrast, many tissue culture studies conducted with B. monnieri in a heavy metal-free medium reported a higher number of regenerated shoots than our study (Tiwari et al. 2001; Banerjee and Modi 2010; Yusuf et al. 2011; Jain et al. 2013; Karatas et al. 2013). Similarly, Behera et al. (2015) obtained the number of shoots per explant at 102.00–30.26 in heavy metal-free culture medium (BAP + Gibberellic acid) in B. monnieri. The results showed that even low concentrations of heavy metals had a negative effect on the number of shoots. The results of this study are also in agreement with earlier reports on Brassica juncea L. (John et al. 2009) Lemna minor (Zhang et al. 2017) and Cer-atophyllum demersum L. (Dogan et al. 2018), where addition of heavy metals to media was found to be harmful for plant growth and development, proliferation and root elongation. Similarly, Shukla et al. (2007) reported that the biomass of B. monnieri under the effect of Cd and Cr decreased due to increased heavy metal concentration and exposure duration.
Even if the concentrations are at the same dose, the effects of heavy metals on plants may vary. In the present study, increases in Cd concentrations in terms of the number of shoots per explant have more inhibitory effect than Cr and Pb. Cd is a non-redox metal and causes ROS directly via Haber–Weiss actions. Due to the toxic effect of Cd, exces-sive ROS production and oxidative stress occur in plants. This excess ROS production causes disturbance antioxidant system, electron transport chain and the basic elements of the metabolism (Nahar et al. 2016). Excessive ROS in cells may provoke oxidation and modification of cellular amino acids, proteins, membrane lipids, and DNA (Emsen and Dogan 2018). These changes lead to oxidative injuries and lead to reduced plant growth and development (Ogawa and Iwabuchi 2001; Yadav 2010). Furthermore, it has been reported in many papers that exposure of the plant to the Cd may cause many effects such as reduction in carbon produc-tion, preventing chlorophyll synthesis, decreasing nutrient uptake, inhibiting photosynthetic activity, oxidative stress, regression of plant growth, and even cell death resulted from excessive or prolonged exposure (Das et al. 1997; Baryla et al. 2001; Pietrini et al. 2003; Hu et al. 2013; Ovečka and Takáč 2014).
The photosynthetic pigment contents of B. monnieri, which was treated to heavy metals, decreased significantly. In comparison to control, the lowest contents of the photo-synthetic pigment in B. monnieri (chlorophyll a, b, a + b and carotenoid) were determined in the highest Cd, Cr, and Pb treatments (1.00 mg/kg). These results showed that the con-centration of chlorophylls and carotenoids decreased with increasing heavy metal concentration. Similarly, Duman and Koca (2014), Ling et al. (2017) and Chen et al. (2015)
reported that the lowest chlorophyll and carotenoid contents were obtained at the highest heavy metal concentrations. The main reasons for the decrease in chlorophyll and carot-enoid levels in plants were the increase in ROS production in cells due to heavy metal origin. In addition, carotenoids play an important role in the prevention of several degenerative stresses in plants due to their antioxidant functions. Carot-enoids are phyto-inhibitors in reducing high levels of ROS as chain-breaker antioxidants (Sytar et al. 2013).
As a result of toxic effects of heavy metals, many symp-toms such as chlorosis, necrosis, softening were observed in B. monnieri. These toxic symptoms vary according to the type of metal. The lowest chlorophyll values were recorded in the plants treated with Pb; therefore, the most toxic metal on photosynthetic pigmentation. These results showed that the effects of heavy metal types on photosynthetic pigments could change. Similarly, Fargašová (2004) investigated the ecotoxic effects of Cd, Cu, Pb, Se, and Zn in Sinapis alba L. and determined the most toxic metal on photosynthetic pigment contents (chlorophyll a, b, total carotenoids) as Pb. In contrast, Burzynski and Klobus (2004) examined the toxic effects of Cu, Cd, and Pb in the leaves of Cucumis sativus L. and reported that Cu is more toxic than other metals in terms of photosynthetic activity and chlorophyll a + b con-tents. Tauqeer et al. (2016) reported that the photosynthetic pigment content of Alternanthera bettzickiana increased in medium containing low concentrations of Cd and Pb but decreased in photosynthetic pigments with higher Cd and Pb applications. These results show that the effect of heavy metals on photosynthetic pigments changes depending on both metal type and plant species.
The most typical symptoms of Pb toxicity are to inhibit the photosynthetic pigment content and photosynthetic activity (Singh et al. 2010). The decrease in photosynthetic activity in plants exposed to Pb is due to the deterioration of chloroplast infrastructure, inhibition of chlorophyll, plastoquinone and carotenoid synthesis, inadequate intake of essential elements such as Mn and Fe, substitution of divalent cations with Pb and disruption of electron transport system (Sharma and Dubey 2005; Pourrut et al. 2011). In addition, Pb reduces the rate of photosynthetic activity by inhibiting the activities of the Calvin cycle enzymes and the opening and closing of the stomas (Romanowska et al. 2006; Mishra et al. 2006; Liu et al. 2008).
According to the results, the second-toxic metal on the photosynthetic pigment was Cr. The Cr-induced decrease in photosynthetic pigments may be related to the deterioration of the activities of various enzymes in chlorophyll biosyn-thesis, such as δ-aminolevulinic acid dehydratase (ALAD or porphobilinogen synthase) and protochlorophyllide reductase (Singh et al. 2013). Cr stress has been reported to decrease chlorophyll content by increasing the content of δ-aminolevulinic acid (ALA) in Nymphaea alba (Vajpayee
et al. 2000). In addition, Cr inhibits the uptake of Mg and N from the plant, resulting in a decrease in photosynthetic pigments (Singh et al. 2013), and interferes with the elec-tron transport system and reduces the photosynthetic activity (Nichols et al. 2000).
Many negative effects of Cd on chlorosis, necrosis, colour fading, deterioration in mineral uptake, imbalance in carbo-hydrate metabolism and reduction in biomass amount have been reported in plants (Azevedo et al. 2005). Moreover, photosynthetic activity and elements of photosynthesis are very sensitive to Cd. Cadmium interferes with photosyn-thetic activity by inhibiting the synthesis of the protochlo-rophyll reductase and δ-aminolevulinic acid in chloprotochlo-rophyll biosynthesis (Zengin and Munzuroglu 2005). It affects pho-tosynthetic electron transport especially with Fe deficiency (Arena et al. 2017), which reduces the photosynthetic rates, PSII efficiency, chlorophyll, and carotenoid contents (Larbi et al. 2002). Manganese is an important element in chloro-plasts and it has been reported that Cd toxicity may cause reduction of manganese and thus reduce photosynthetic activity (Lagriffoul et al. 1998). Similarly, Lactuca sativa exposed to Cd showed that chlorophyll contents decreased significantly and chlorophyll a was more affected than chlo-rophyll b (Dias et al. 2012).
The protein contents of B. monnieri exposed to Cd, Cr, and Pb decreased significantly compared to the control group. However, the lowest metal concentration (0.25 mg/ kg) had a low increase in protein contents compared to the control and they decreased significantly in higher metal applications (p < 0.05). It has been reported that short-term protein increase can be seen in plants under stress of metal with induction of stress proteins such as antioxidant enzymes, heat-shock proteins and enzymes in GSH and PC biosynthesis (Srivastava et al. 2005; Dogan et al. 2018). In addition, antioxidants such as glutathione, ascorbic acid, proline, and cysteine and antioxidant enzymes such as superoxide dismutase, catalase, ascorbate peroxidase, and glutathione reductase, which result from metal toxicity, affect the amount of protein (Gupta et al. 2009). Similar to the findings, low levels of protein increase in heavy metal treated plants were also determined by Sinha et al. (2005), Mishra et al. (2008) and Dogan et al. (2018).
Although all metals decreased the protein level compared to the control group, the lowest protein contents were meas-ured in Cd-treated plants (41.40% lower than the control). Protein synthesis associated with growth parameters is sig-nificantly decreased by Cd applications. The reason for this decrease may be due to the damage of many proteins as a result of the increase in the amount of ROS and the dena-turation of proteins with heavy-metal toxicity and the deg-radation of proteins by protease or other catabolic enzymes (RNAase and DNAase enzymes). In addition, Cd toxicity can disrupt the protein synthesis mechanism by inhibiting
functional sulfhydryl groups of enzymes (Duman et al. 2010; Bavi et al. 2011).
Metal stress can cause cell-membrane damage by sig-nificantly increasing ROS such as superoxide radicals (O2−) and H2O2 in plants (Hsu and Guo 2002). Lipid peroxida-tion is defined as the oxidative degradaperoxida-tion of membrane lipids. Malondialdehyde (MDA) content is accepted as a determinant of lipid peroxidation (Yildiz et al. 2011). In the study, changes in MDA and H2O2 levels of B. monnieri which were affected by different concentrations of Cd, Cr, and Pb were determined Fig. 5b, c. The results showed that the concentration of MDA in the plant increased with the increase of metal level in the media. Li et al. (2015) have found higher MDA content in Arachis hypogaea L. under Cd stress compared to control. Mishra et al. (2006) reported that MDA content of C. demersum exposed to 1–100 µM Pb was increased compared to control. MacFarlane and Bur-chett (2001) reported an increase in lipid peroxidation of Avicennia marina (Forsk.) Vierh under heavy metal stress and explained that photosynthetic pigment content and lipid peroxidation data can be considered as biological indicators of stress.
When the cellular effects of heavy metals such as Cd, Cr, Pb, and Hg are examined, it was observed that heavy metals produced superoxide ion, hydrogen peroxide, and hydroxyl radicals by consuming protein-bound sulfhydryl groups and glutathione (Stohs and Bagchi 1995). This toxic ROS can lead to oxidation of membrane proteins and lipids, and even to cell death (Khaliq et al. 2015). In the present study, a positive relationship was determined between lipid peroxi-dation (MDA contents) and hydrogen peroxide. Both values increased with metal concentration. Similarly, Anjum et al. (2015b) reported that the MDA ve H2O2 content of Zea mays L. exposed to different concentrations of Cd (0–375 µM) increased compared to control. Ulusu et al. (2017) reported increased levels of H2O2 in Phaseolus vulgaris L. due to increased Cd concentration.
5 Conclusion
As a result of anthropogenic activities, heavy metals released to the environment show negative effects on all living things, especially plants. With the rapid population growth and industrialization, the heavy-metal types and levels that the plants are exposed to also increase. In this study, the effects of Cd, Cr, and Pb on the production of B. monnieri by tissue culture techniques were evaluated. In addition, the phytotox-icity of these heavy metals in the plant was investigated. The negative effects of Cd, Cr, and Pb on the in vitro production of B. monnieri were clearly demonstrated. The number of shoots per explant was significantly decreased compared to the control. Likewise, heavy metals damage the plant by
affecting vital parameters such as photosynthetic pigment content, protein content, and lipid peroxidation. This study is very important in terms of showing the effects of heavy metals spreading to the environment with fast industrializa-tion on the producindustrializa-tion and biochemical parameters of plants. Compliance with ethical standards
Conflict of interest The author declares that he has no conflict of
inter-est.
References
Agır SU, Kutbay HG, Surmen B, Elmas E (2017) The effects of erosion and accretion on plant communities in coastal dunes in north of Turkey. Rend Fis Acc Lincei 28:203–224
Ahmad SS, Reshi ZA, Shah MA, Rashid I, Ara R, Andrabi SMA (2016) Heavy metal accumulation in the leaves of Potamogeton natans and Ceratophyllum demersum in a Himalayan RAMSAR site: management implications. Wetl Ecol Manag 24:469–475 Amari T, Ghnaya T, Abdelly C (2017) Nickel, cadmium and lead
phy-totoxicity and potential of halophytic plants in heavy metal extrac-tion. S Afr J Bot 111:99–110
Anjum NA, Sofo A, Scopa A, Roychoudhury A, Gill SS, Iqbal M, Lukatkin AS, Pereira E, Duarte AC, Ahmad I (2015a) Lipids and proteins-major targets of oxidative modifications in abiotic stressed plants. Environ Sci Pollut Res 22:4099–4121
Anjum SA, Tanveer M, Hussain S, Bao M, Wang L, Khan I, Ullah E, Tung SA, Samad RA, Shahzad B (2015b) Cadmium toxicity in Maize (Zea mays L.): consequences on antioxidative systems, reactive oxygen species and cadmium accumulation. Environ Sci Pollut Res 22:17022–17030
Arena C, Figlioli F, Sorrentino MC, Izzo LG, Capozzi F, Giordano S, Spagnuolo V (2017) Ultrastructural, protein and photosynthetic alterations induced by Pb and Cd in Cynara cardunculus L., and its potential for phytoremediation. Ecotoxicol Environ Saf 145:83–89
Azevedo H, Pinto CGG, Fernandes J, Loureiro S, Santos C (2005) Cadmium effects on sunflower growth and photosynthesis. J Plant Nutr 28:2211–2220
Banerjee M, Modi P (2010) Micropropagation of Bacopa monnieri using cyanobacterial liquid medium. Plant Tissue Cult Biotechnol 20:225–231
Baryla A, Carrier P, Franck F, Coulomb C, Sahut C et al (2001) Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cad-mium-polluted soil: causes and consequences for photosynthesis and growth. Planta 212:696–709
Bavi K, Kholdebarin B, Moradshahi A (2011) Effect of cadmium on growth, protein content and peroxidase activity in pea plants. Pak J Bot 43:1467–1470
Behera S, Nayak N, Shasmita Barik DP, Naik SK (2015) An efficient micropropagation protocol of Bacopa monnieri (L.) Pennell through two-stage culture of nodal segments and ex vitro accli-matization. J Appl Biol Biotechnol 3:16–21
Bhusari S, Wanjari R, Khobragade P (2013) Cost effective in vitro clonal propagation of Bacopa monnieri L. Penell. Int J Ind Med Plants 46:1239–1244
Burzyński M, Kłobus G (2004) Changes of photosynthetic parameters in cucumber leaves under Cu, Cd, and Pb stress. Photosynthetica 42:505–510
Chen C, Huang D, Liu J (2009) Functions and toxicity of nickel in plants: recent advances and future prospects. Clean Soil Air Water 37:304–313
Chen M, Zhang LL, Li J, He XJ, Cai JC (2015) Bioaccumulation and tolerance characteristics of a submerged plant (Ceratophyllum demersum L.) exposed to toxic metal lead. Ecotoxicol Environ Saf 122:313–321
Das P, Samantaray S, Rout G (1997) Studies on cadmium toxicity in plants: a review. Environ Pollut 98:29–36
Dias MC, Monteiro C, Moutinho-Pereira J, Correia C, Gonçalves B, Santos C (2012) Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol Plant 35:1281–1289 Dogan M, Emsen B (2018) Anti-cytotoxic-genotoxic influences of
in vitro propagated Bacopa monnieri L. Pennell in cultured human lymphocytes. Eurasian J Biol Chem Sci 1:48–53
Dogan M, Akgul H, Inan OG, Zeren H (2015) Determination of cad-mium accumulation capabilities of aquatic macrophytes Cerato-phyllum demersum, Bacopa monnieri and Rotala rotundifolia. Iran J Fish Sci 14:1010–1017
Dogan M, Karatas M, Aasim M (2018) Cadmium and lead bioaccumu-lation potentials of an aquatic macrophyte Ceratophyllum demer-sum L.: a laboratory study. Ecotoxicol Environ Saf 148:431–440 Duman F, Koca FD (2014) Single and combined effects of exposure
concentration and duration on biological responses of Cerato-phyllum demersum L. exposed to Cr species. Int J Phytoremediat 16:1192–1208
Duman F, Şahan S, Ceylan A, Koca FD (2010) Biological responses of Ceratophyllum demersum L. exposed to chromium (Cr+6). SDU
J of Sci 5:163–171
Emsen B, Dogan M (2018) Evaluation of Antioxidant activity of in vitro propagated medicinal Ceratophyllum demersum L. extracts. Acta Sci Pol Hortoru 17:23–33
Fargašová A (2004) Toxicity comparison of some possible toxic metals (Cd, Cu, Pb, Se, Zn) on young seedlings of Sinapis alba L. Plant Soil Environ 50:33–38
Gajewska E, Wielanek M, Bergier K, Skłodowska M (2009) Nickel induced depression of nitrogen assimilation in wheat roots. Acta Physiol Plant 31:1291–1300
Gay CA, Gebicki JM (2000) A critical evaluation of the effect of sorbi-tol on the ferric-xylenol orange hydroperoxide assay. Anal Bio-chem 284:217–220
Gupta DK, Nicoloso FT, Schetinger MR, Rossato LV, Pereira LB, Cas-tro GY (2009) Antioxidant defense mechanism in hydroponically grown Zea mays seedlings under moderate lead stress. J Hazard Mater 172:479–484
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Bio-chem Biophys 125:189–198
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. Circ Calif Agric Exp Stn 347:1–32
Hsu PC, Guo YL (2002) Antioxidant nutrients and lead toxicity. Toxi-cology 180:33–34
Hu YF, Zhou G, Na XF, Yang L, Nan WB, Liu X, Zhang YQ, Li JL, Bi YR (2013) Cadmium interferes with maintenance of auxin home-ostasis in Arabidopsis seedlings. J Plant Physiol 170:965–975 Jain R, Prasad B, Jain M (2013) In vitro regeneration of Bacopa
mon-nieri (L.): a highly valuable medicinal plant. Int J Curr Microbiol Appl Sci 2:198–205
Jain S, Singh A, Khare P, Chanda D, Mishra D, Shanker K, Karak T (2017) Toxicity assessment of Bacopa monnieri L. grown in biochar amended extremely acidic coal mine spoils. Ecol Eng 108:211–219
John R, Ahmad P, Gadgil K, Sharma S (2009) Heavy metal toxicity: effect on plant growth, biochemical parameters and metal accumu-lation by Brassica juncea L. Int J Plant Prod 3(3):65–76
Karatas M, Aasim M, Dogan M, Khawar KM (2013) Adventitious shoot regeneration of the medicinal plant water hyssop (Bacopa monnieri L. Pennell) using different ınternodes. Arch Biol Sci 65:297–303
Kashyap S, Kapoor N, Kale RD (2018) Micropropagation of B. mon-nieri using humin media in plant tissue culture. Ann Plant Sci 6:1625–1629
Kean JD, Downey LA, Stough C (2017) Systematic overview of Bacopa monnieri (L.) Wettst. dominant poly-herbal formulas in children and adolescents. Medicines 4(4):86
Khaliq A, Aslam F, Matloob A, Hussain S, Geng M, Wahid A, Rehman H (2015) Seed priming with selenium: consequences for emer-gence, seedling growth, and biochemical attributes of rice. Biol Trace Elem Res 166(2):236–244
Kılıç DD, Kutbay HG, Sürmen B, Hüseyinoğlu R (2018) The classi-fication of some plants subjected to disturbance factors (grazing and cutting) based on ecological strategies in Turkey. Rend Lincei Sci Fis 29:87–102
Lagriffoul A, Mocquot B, Mench M, Vangronsveld J (1998) Cadmium toxicity effects on growth, mineral and chlorophyll contents, and activities of stress related enzymes in young maize plants (Zea mays L.). Plant Soil 200:241–250
Larbi A, Morales F, Abadía A, Gogorcena Y, Lucena JJ, Abadía J (2002) Effects of Cd and Pb in sugar beet plants grown in nutri-ent solution: induced Fe deficiency and growth inhibition. Funct Plant Biol 29:1453–1464
Leung DWM (2017) Potential of plant tissue culture research contrib-uting to combating arsenic pollution. In: Gupta D, Chatterjee S (eds) Arsenic contamination in the environment. Springer, Cham, pp 187–194
Li CJ, Yan CX, Liu Y, Zhang TT, Wan SB, Shan SH (2015) Phytotoxic-ity of cadmium on peroxidation, superoxide dismutase, catalase and peroxidase activities in growing peanut (Arachis hypogaea L.). Afr J Biotechnol 14:1151–1157
Lichtenthaler HK, Wellburn AR (1983) Determinations of total carot-enoids and chlorophylls a and b of leaf extracts in different sol-vents. Biochem Soc Trans 11:591–592
Ling T, Gao Q, Du H, Zhao Q, Ren J (2017) Growing, physiological responses and Cd uptake of Corn (Zea mays L.) under different Cd supply. Chem Spec Bioavailab 29:216–221
Liu D, Li T, Jin X, Yang X, Islam E, Mahmood Q (2008) Lead induced changes in the growth and antioxidant metabolism of the lead accumulating and non-accumulating ecotypes of Sedum alfredii. J Integr Plant Biol 50:129–140
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein meas-urement with the Folin-phenol reagent. J Biol Chem 193:265–275 MacFarlane GR, Burchett MD (2001) Photosynthetic pigments and
peroxidase activity as indicators of heavy metal stress in the grey mangrove, Avicennia marina (Forsk.) Vierh. Mar Pollut Bull 42:233–240
Maleki M, Ghorbanpour M, Kariman K (2017) Physiological and anti-oxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene 11:247–254
Matheka JM, Magiri E, Rasha AO, Machuka J (2008) In vitro selection and characterization of drought tolerant somaclones of tropical maize (Zea mays L.). Biotechnology 7:641–650
Mejáre M, Bülow L (2001) Metal-binding proteins and peptides in bioremediation and phytoremediation of heavy metals. Trends Biotechnol 19:67–73
Mishra S, Srivastava S, Tripathi RD, Kumar R, Seth CD, Gupta DK (2006) Lead detoxification by coontail (Ceratophyllum demersum L.) involves induction of hytochelatins and antioxidant system in response to its accumulation. Chemosphere 65:1027–1039 Mishra S, Srivastava S, Tripathi RD, Dwivedi S, Shukla MK (2008)
demersum L.) plants under cadmium stress. Environ Technol 23:294–301
Mishra S, Srivastava S, Dwivedi S, Tripathi RD (2013) Investigation of biochemical responses of Bacopa monnieri L. upon exposure to arsenate. Environ Toxicol 28:419–430
Mohanpuria P, Rana NK, Yadav SK (2007) Cadmium induced oxida-tive stress influence on glutathione metabolic genes of Camellia sinensis (L.) O. Kuntze. Environ Toxicol 22:368–374
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Nagajyoti PC, Lee KD, Sreekanth TVM (2010) Heavy metals, occur-rence and toxicity for plants: a review. Environ Chem Lett 8:199–216
Nahar K, Hasanuzzaman Alam M, Rahman A, Suzuki T, Fujita M (2016) Polyamine and nitric oxide crosstalk: antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol Environ Saf 126:245–255 Naik PM, Godbole M, Nagella P, Murthy HN (2017) Influence of
dif-ferent media, medium strength and carbon sources on adventitious shoot cultures and production of bacoside A in Bacopa monnieri (L.). Ceylon J Sci 46:97–104
Nichols PB, Couch JD, Al-Hamdani SH (2000) Selected physiological responses of Salvinia minima to different chromium concentra-tions. Aquat Bot 68:313–319
Ogawa K, Iwabuchi M (2001) A mechanism for promoting the germi-nation of Zinnia elegans seeds by hydrogen peroxide. Plant Cell Physiol 42:286–291
Ogbonna PC, Nzegbule EC, Obasi KO, Kanu H (2018) Heavy metals in soil and accumulation in medicinal plants at an industrial area in Enyimba city, Abia State, Nigeria. Niger J Environ Sci Technol 2:89–95
Ovečka M, Takáč T (2014) Managing heavy metal toxicity stress in plants: biological and biotechnological tools. Biotechnol Adv 32:73–86
Pietrini F, Iannelli MA, Pasqualini S, Massacci A (2003) Interaction of cadmium with glutathione and photosynthesis in developing leaves and chloroplasts of Phragmites australis (Cav.) Trin. ex Steudel. Plant Physiol 133:829–837
Pourrut B, Shahid M, Dumat C, Winterton P, Pinelli R (2011) Lead uptake, toxicity, and detoxification in plants. Rev Environ Contam Toxicol 2011:113–136
Rai MK, Kalia RK, Singh R, Gangola MP, Dhawana AK (2011) Devel-oping stress tolerant plants through in vitro selection—an over-view of the recent progress. Environ Exp Bot 71:89–98
Rezende ALSR, Joyce Doria RS, Heloisa Oliveira S, Moacir P, Roberto Alves B, Reis RO, Filipe Almendagna R, Jose Darlan R (2017)
Effects of silicon on antioxidant enzymes, CO2, proline and
bio-logical activity of in vitro-grown cape gooseberry under salinity stress. Aust J Crop Sci 11:438–446
Romanowska E, Wróblewska B, Drozak A, Siedlecka M (2006) High light intensity protects photosynthetic apparatus of pea plants against exposure to lead. Plant Physiol Biochem 44:387–394 Russo A, Borrelli F (2005) Bacopa monniera, a reputed nootropic
plant: an overview. Phytomedicine 12:305–317
Safarnejad A (2004) Characterization of somaclones of Medicago sativa L. for drought tolerance. J Agric Sci Technol 6:121–127 Sarwar N, Imran M, Shaheen MR, Ishaque W, Kamran MA, Matloob
A, Rehim A, Hussaine S (2017) Phytoremediation strategies for soils contaminated with heavy metals: modifications and future perspectives. Chemosphere 171:710–721
Sharma P, Dubey RS (2005) Lead toxicity in plants. Braz J Plant Physiol 17:35–52
Sharma S, Kamal B, Rathi N, Chauhan S, Jadon V, Vats N, Gehlot A, Arya S (2010) In vitro rapid and mass multiplication of highly
valuable medicinal plant Bacopa monnieri (L.) Wettst. Afr J Biotechnol 9:8318–8322
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mecha-nism in plants under stressful conditions. J Bot 2012:1–26 Shukla OP, Dubey S, Rai UN (2007) Preferential accumulation
of cadmium and chromium: toxicity in Bacopa monnieri L. under mixed metal treatments. Bull Environ Contam Toxicol 78:252–257
Singh R, Tripathi RD, Dwivedi S, Kumar A, Trivedi PK, Chakra-barty D (2010) Lead bioaccumulation potential of an aquatic macrophyte Najas indica are related to antioxidant system. Bioresour Technol 101:3025–3032
Singh HP, Mahajan P, Kaur S, Batish DR, Kohli RK (2013) Chro-mium toxicity and tolerance in plants. Environ Chem Lett 11:229–254
Sinha S, Saxena R, Singh S (2005) Chromium induced lipid peroxida-tion in the plants of Pistia stratiotes L: role of antioxidants and antioxidant enzymes. Chemosphere 58:598–604
Smith E, Palethorpe HM, Tomita Y, Pei JV, Townsend AR, Price TJ, Young JP, Yool AJ, Hardingham JE (2018) The purified extract from the medicinal plant Bacopa monnieri, bacopaside II, inhibits growth of colon cancer cells in vitro by inducing cell cycle arrest and apoptosis. Cells 7(7):81
Snedecor GW, Cochran WG (1997) Statistical methods. The Iowa State University Press, Iowa
Srivastava S, Mishra S, Dwivedi S, Baghel VS, Verma S, Tandon PK, Rai UN, Tripathi RD (2005) Nickel phytoremediation potential of broad bean Vicia faba L. and its biochemical responses. Bull Environ Contam Toxicol 74:715–772
Srivastava P, Tiwari KN, Srivastava G (2017) Effect of different carbon sources on in vitro regeneration of Brahmi Bacopa monnieri (L.) an important memory vitalizer. J Med Plants Stud 5:202–208 Stohs SJ, Bagchi D (1995) Oxidative mechanisms in the toxicity of
metal ions. Free Radic Biol Med 18:321–336
Summart J, Thanonkeo P, Panichajakul S, Prathepha P, McManus MT (2010) Effect of salt stress on growth, inorganic ion and proline accumulation in Thai aromatic rice, Khao Dawk Mali 105, callus culture. Afr J Biotechnol 9:145–152
Sytar O, Kumar A, Latowski D, Kuczynska P, Strzałka K, Prasad MNV (2013) Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol Plant 35:985–999
Tauqeer HM, Ali S, Rizwan M, Ali Q, Saeed R, Iftikhar GU (2016) Phytoremediation of heavy metals by Alternanthera bettzicki-ana: growth and physiological response. Ecotoxicol Environ Saf 126:138–146
Tiwari V, Tiwari KN, Singh BD (2001) Comparative studies of cyto-kinins on in vitro propagation of Bacopa monniera. Plant Cell Tissue Org Cult 66:9–16
Uabundit N, Wattanathorn J, Mucimapura S, Ingkaninan K (2010) Cog-nitive enhancement and neuroprotective effects of Bacopa mon-nieri in Alzheimer’s disease model. J Ethnopharmacol 127:26–31 Ulusu Y, Öztürk L, Elmastaş M (2017) Antioxidant capacity and cad-mium accumulation in parsley seedlings exposed to cadcad-mium stress. Russ J Plant Physiol 64:883–888
Vajpayee P, Tripathi RD, Rai UN, Ali MB, Singh SN (2000) Chro-mium (VI) accumulation reduces chlorophyll biosynthesis, nitrate reductase activity and protein content in Nymphaea alba L. Che-mosphere 41:1075–1082
Wang Z, Yao L, Liu G, Liu W (2014) Heavy metals in water, sediments and submerged macrophytes in ponds around the Dianchi Lake, China. Ecotoxicol Environ Saf 107:200–206
Wójcik M, Tukiendorf A (2004) Phytochelatin synthesis and cadmium localization in wild type of Arabidopsis thaliana. Plant Growth Regul 44:71–80
Xu J, Yin HY, Li X (2009) Protective effects of proline against cad-mium toxicity in micropropagated hyperaccumulator, Solanum nigrum L. Plant Cell Rep 28:325–333
Yadav SK (2010) Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress toler-ance of plants. S Afr J Bot 76:167–179
Yıldız M, Terzi H, Uruşak B (2011) Chromium toxicity and cellular responses in plants. Erciyes Univ J Inst Sci Technol 27:163–176 Yusuf A, Rajesh KT, Nikhilesh S, Rao PS (2011) Effects of antioxi-dants and gelling agents on regeneration, in vitro conservation and genetic stability of Bacopa monnieri (L.) Pennell. Int J Ayurvedic Herb Med 1:51–67
Zengin KF, Munzuroğlu Ö (2005) Effects of some heavy metals on content of chlorophyll, proline and some antioxidant chemicals
in bean (Phaseolus vulgaris L.) seedlings. Acta Biol Crac Bot 47:157–164
Zhang T, Lu Q, Su C, Yang Y, Hu D, Xu Q (2017) Mercury induced oxidative stress, DNA damage, and activation of antioxidative sys-tem and Hsp70 induction in duckweed (Lemna minor). Ecotoxicol Environ Saf 143:46–56
Publisher’s Note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.