T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
BOR STRESĠNĠN BAZI HAVUÇ
GENOTĠPLERĠNĠN TEMEL FĠZYOLOJĠK VE BĠYOKĠMYASAL ÖZELLĠKLERĠNE
ETKĠLERĠ
AYġE HÜMEYRA OMAY YÜKSEK LĠSANS TEZĠ
Toprak Bilimi ve Bitki Besleme Anabilim Dalı
Aralık-2019 KONYA Her Hakkı Saklıdır
TEZ KABUL VE ONAYI
AyĢe Hümeyra OMAY tarafından hazırlanan “Bor Stresinin Bazı Havuç
Genotiplerinin Temel Fizyolojik ve Biyokimyasal Özelliklerine Etkileri” adlı tez
çalıĢması …/.../…. tarihinde aĢağıdaki jüri tarafından oy birliği / oy çokluğu ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü Toprak Bilimi ve Bitki Besleme Anabilim Dalı’nda YÜKSEK LĠSANS olarak kabul edilmiĢtir.
Jüri Üyeleri Ġmza
BaĢkan
Prof. Dr. Erdoğan EĢref HAKKI ………..
DanıĢman
Doç. Dr. Mehmet HAMURCU ………..
Üye
Doç. Dr. Ceyda ÖZFĠDAN KONAKÇI ………..
Yukarıdaki sonucu onaylarım.
Prof. Dr. Mustafa YILMAZ FBE Müdürü
Bu tez çalıĢması TÜBĠTAK tarafından 218O137 nolu proje ve S.Ü. BAP kordinatörlüğü tarafından 1940166 nolu proje ile desteklenmiĢtir.
TEZ BĠLDĠRĠMĠ
Bu tezdeki bütün bilgilerin etik davranıĢ ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalıĢmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all material and results that are not original to this work.
AyĢe Hümeyra OMAY 03.12.2019
iv
ÖZET
YÜKSEK LĠSANS TEZĠ
BOR STRESĠNĠN BAZI HAVUÇ GENOTĠPLERĠNĠN TEMEL FĠZYOLOJĠK VE BĠYOKĠMYASAL ÖZELLĠKLERĠNE ETKĠLERĠ
AyĢe Hümeyra OMAY
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Toprak Bilimi ve Bitki Besleme DanıĢman: Doç. Dr. Mehmet HAMURCU
2019, 81 Sayfa Jüri
Doç. Dr. Mehmet HAMURCU Prof. Dr. Erdoğan EĢref HAKKI Doç. Dr. Ceyda ÖZFĠDAN KONAKÇI
ÇalıĢmanın amacı, bor noksan ve bor toksik koĢullara adaptasyon problemi yaĢamayan genotipleri belirlemek için havuç bitkisinde fizyolojik ve biyokimyasal değiĢimlerin araĢtırılmasıdır. ÇalıĢmada bitki materyali olarak Türkiye’de ekimi yapılan ve ticari öneme sahip 4 farklı havuç genotipi kullanılmıĢtır. Genotiplere 5 farklı bor dozu (kontrol (0.0033 mM), 0 mM B, 0.5 mM B, 1 mM B, 2 mM B ) uygulanmıĢtır. Deneme tam kontrollü iklim odasında, su kültürü ortamında dört tekerrürlü olarak kurulmuĢtur.
Havuç genotipleri, bor noksan ve bor toksik uygulamalarından önemli ölçüde etkilenmiĢtir. Uygulamalara bağlı olarak gövde ve kök temel büyüme parametreleri (boy, yaĢ ağırlık ve kuru ağırlık) kontrol ile kıyaslandığında azalmaların meydana geldiği tespit edilmiĢtir. Genotipler içerisinden 6 numaralı genotipin büyüme parametre değerlerindeki azalmaların daha düĢük oranlarda olduğu belirlenmiĢtir. Genotiplerin bor içeriklerinde, bor noksan koĢullarda azalmalar olduğu, artan bor dozlarına bağlı olarak ise artıĢlar meydana geldiği tespit edilmiĢtir. Bor noksan ve bor toksik koĢullarda genotiplerde strese bağlı olarak EC düzeylerinde ve MDA içeriklerinde artıĢlar olduğu belirlenirken, 13 numaralı genotipte bu artıĢların minumum seviyede olduğu tespit edilmiĢtir. SOD, APOX, POX, GR enzim aktivite değerlerinde genel olarak artıĢlar meydana gelirken, 12 numaralı genotipte uygulamalara bağlı olarak azalmalar meydana geldiği tespit edilmiĢtir.Yapılan analiz sonuçları göz önüne alındığında elde edilen ortalama değerlere göre, bor noksan koĢullara karĢı 13 numaralı genotipin, bor toksik koĢullara karĢı 6 numaralı genotipin toleranslılık seviyelerinin yüksek olduğu gözlemlenmiĢtir.
Anahtar Kelimeler: Antioksidan enzimler, bor noksanlığı, bor toksisitesi, havuç, serbest radikaller.
v
ABSTRACT MS THESIS
THE EFFECTS OF BORON STRESS ON BASIC PHYSIOLOGICAL AND BIOCHEMICAL PROPERTIES OF SOME CARROT GENOTYPES
AyĢe Hümeyra OMAY
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY
THE DEGREE OF MASTER OF SCIENCE
IN DEPARTMENT OF SOIL SCIENCE AND PLANT NUTRITION Advisor: Assoc. Prof. Dr. Mehmet HAMURCU
2019, 81 Pages Jury
Assoc. Prof. Dr. Mehmet HAMURCU Prof. Dr. Erdoğan EĢref HAKKI
Assoc. Prof. Dr. Ceyda ÖZFĠDAN KONAKÇI
The aim of this study was to investigate the physiological and biochemical changes in carrot in order to determine the genotypes without adaptation problems to boron deficient and boron toxic conditions. In this study, the four carrot genotypes which are planted in Turkey and have commercially the most important were used as plant material. Five different boron doses (control (0.0033 mM), 0 mM B, 0.5 mM B, 1 mM B, 2 mM B) were applied to the genotypes. The experiment was set up with four replications in a fully controlled climate chamber in a water culture medium.
Carrot genotypes were significantly affected by boron deficient and boron toxic applications. Depending on the applications, it was determined that the decreases occurred in stem and root growth parameters (height, fresh weight, and dry weight) when compared with the control. Among the genotypes, the decrease in the growth parameter values of genotype 6 was found to be lowest. It was determined that boron content of genotypes decreased in boron deficient conditions, however depending on the increasing boron doses, the increases occurred. In boron deficient and boron toxic conditions, the increases in EC levels and MDA contents of genotypes were determined due to stress, while these increases in genotype number 13 was found to be at a minimum level. While SOD, APOX, POX, and GR enzyme activity values generally increased, in genotype number 12, it was determined that the decreases occurred due to applications. When the results of the analysis were considered, it was observed that the tolerance levels of genotype 13 against boron-deficient conditions and genotype 6 against boron-toxic conditions were high according to the obtained values.
vi
ÖNSÖZ
Tezimin planlanmasında ve yürütülmesinde bilgi ve deneyimlerini esirgemeyen ve bana her zaman yol gösteren danıĢmanım sayın Doç. Dr. Mehmet HAMURCU hocama çok teĢekkür ederim. Ayrıca çalıĢmamda bilgilerinden faydalandığım Prof. Dr. Sait GEZGĠN hocama ve desteklerinden dolayı diğer Toprak Bilimi ve Bitki Besleme Bölüm hocalarıma teĢekkür ederim.
Tez çalıĢmam boyunca hep yanımda olan değerli ekip arkadaĢlarım; Z. Zeynep AVġAROĞLU’na, Ġsmail KAYA’ya, Canan ÜSTÜN’e ve Fevzi ELBASAN’a yardımlarından dolayı teĢekkür ederim.
Manevi desteklerinden dolayı değerli hocam ArĢ. Gör. Vildan ERCĠ’ye, değerli ablam Öznur YALÇIN’a, kıymetli arkadaĢlarım Nazife Sultan EFE’ye ve Huriye TÜRKOĞLU’na çok teĢekkür ederim.
Eğitim hayatım boyunca, maddi manevi desteklerini asla esirgemeyen canım annem Fatma OMAY’a, canım babam Mehmet OMAY’a, canım kız kardeĢim Amine Rüveyha OMAY’a, eğitim hayatım boyunca, her zorlukta yanımda olan canım kız kardeĢim Makbule Rumeysa OMAY’a çok teĢekkür ederim.
Ayrıca tez çalıĢmamı finansal olarak destekleyen TÜBĠTAK (Proje No: 218O137) ve S.Ü. BAP kordinatörlüğüne (Proje No: 1940166) teĢekkür ederim.
AyĢe Hümeyra OMAY KONYA-2019
vii ĠÇĠNDEKĠLER ÖZET ... iv ABSTRACT ... v ÖNSÖZ ... vi ĠÇĠNDEKĠLER ... vii SĠMGELER VE KISALTMALAR ... ix ÇĠZELGELER DĠZĠNĠ ... xi ġEKĠLLER DĠZĠNĠ ... xiii 1. GĠRĠġ ... 1 2. KAYNAK ARAġTIRMASI ... 4
2.1. Bor Elementi ve Bitkiler Üzerine Etkileri ... 5
2.2. Bitkilerde Bor Stresi ve Savunma Sistemleri ... 9
3. MATERYAL VE YÖNTEM ... 14
3.1. Materyal ... 14
3.2. Yöntem ... 14
3.2.1. Deneme serilerinin hazırlanması ... 14
3.2.2. Tohum dikimi ... 15
3.2.3. Örnekleme ... 16
3.3. Laboratuvar Analiz Yöntemleri ... 17
3.3.1. Bitki örneklerinde fizyolojik analizler ... 17
3.3.2. Biyokimyasal parametrelerin ölçümü ... 18
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 23
4.1. Büyüme Parametreleri Bulguları ... 23
4.1.1. Gövde ve kök yaĢ ağırlığı ... 24
4.1.2. Gövde ve kök kuru ağırlığı ... 28
4.1.3. Gövde ve kök boyu ... 32
4.2. Gövde B Konsantrasyonu ... 38
4.3. Bağıl Su Ġçeriği ... 41
4.4. Prolin Seviyeleri ... 43
4.5. Lipid Peroksidasyon Düzeyleri ... 46
4.6. Elektrolit Sızıntısı ... 48
4.7. Reaktif Oksijen Türlerine Ait Sonuçlar ... 50
4.7.1. Hidroksil radikali (·OH-) süpürülme aktivitesi ... 50
4.7.2. Hidrojen peroksit (H2O2) süpürülme aktivitesi ... 54
4.8. Antioksidan Aktivite Sonuçları ... 56
viii
4.8.2. Peroksidaz enzim aktiviteleri (POX) ... 59
4.8.3. Katalaz enzim aktiviteleri (CAT) ... 61
4.8.4. Glutatyon redüktaz enzim aktiviteleri (GR) ... 63
4.8.5. Askorbat peroksidaz enzim aktiviteleri (APX) ... 65
4.8.6. Gulutatyon miktarı (GSH) ... 68
5. SONUÇLAR VE ÖNERĠLER ... 70
5.1 Sonuçlar ... 70
5.2 Öneriler ... 73
ix
SĠMGELER VE KISALTMALAR Simgeler
Kısaltmalar
B : Bor
B(OH)4- : Borat Ġyotları
DAB : 3'-3'-Diaminobenzidin Tetrahidroklorit
EDTA : Etilen Diamin Tetra Asetik Asit
Fe : Demir
FeSO4 : Demir Sülfat
H3BO3 : Borik Asit
HCl : Hidrojen Klorür
HNO3 : Nitrik Asit
H2O : Su
H2O2 : Hidrojen Peroksit
NaOH : Sodyum Hidroksil
NADPH : Nikotinamid Adenin Dinükleotit Fosfat
Na : Sodyum
NBT : Nitro Blue Tetrazolium
O2 : Oksijen
O2- : Süperoksit .
OH- : Hidroksil
TBA : Thiobarbituric Acid
APX : Askorbat Peroksidaz
CAT : Katalaz cm : Santimetre o
C : Santigrat Derece
EC : Elektrolit Sızıntısı
Genotip 6 : Mor Havuç
Genotip 9 : Mor Havuç
Genotip 12 : Turuncu-Nantes
Genotip 13 : Sarı-Nantes
GR : Glutatyon Redüktaz
GSH : Glutatyon
g : Gram
g bitki-1 : Gram/ Bitki
KA : Kuru Ağırlık MDA : Malondialdehit mM : Milimolar ml : Mililitre mg L-1 : Miligram/Litre mg kg-1 : Miligram/Kilogram N : Normal nm : Nanometre
x
nmol : Nanomol
POX : Peroksidaz
ppm : Milyonda bir
PVPP : Polivinilpolipirolidon
ROT : Reaktif Oksijen Türleri
RWC : Bağıl/Nispi Su Ġçeriği
SOD : Süperoksit Dismutaz
YA : YaĢ Ağırlık
xi
ÇĠZELGELER DĠZĠNĠ
Çizelge 4.1. B noksan ve B toksik koĢullarda yetiĢtirilen havuç (6, 9, 12, 13)
genotiplerinin temel büyüme parametrelerine ait varyans analiz sonuçları …………...23
Çizelge 4.2. Havuç (6, 9, 12, 13) genotiplerinin gövde yaĢ ağırlık değerleri (g bitki
-1)………...25
Çizelge 4.3. Havuç (6, 9, 12, 13) genotiplerinin kök yaĢ ağırlık değerleri (g bitki-1 ) ……….27
Çizelge 4.4. Havuç (6, 9, 12, 13) genotiplerinin gövde kuru ağırlık değerleri (g bitki -1
)………...29
Çizelge 4.5. Havuç (6, 9, 12, 13) genotiplerinin kök kuru ağırlık değerleri (g bitki -1
)………...……31
Çizelge 4.6. Havuç (6, 9, 12, 13) genotiplerinin gövde boy değerleri (g bitki-1 ). …..……….………..33
Çizelge 4.7. Havuç (6, 9, 12, 13) genotiplerinin kök boy değerleri (g bitki -1
)………...35
Çizelge 4.8. B noksan ve B toksik koĢullarda yetiĢtirilen havuç (6, 9, 12, 13)
genotiplerinin gövde B konsantrasyonu, RWC, EC, MDA ve prolin değerlerine ait varyans analiz sonuçları ………..38
Çizelge 4.9. Havuç (6, 9, 12, 13) genotiplerinin gövde B konsantrasyon değerleri (mg g
-1)………...39
Çizelge 4.10. Havuç (6, 9, 12, 13) genotiplerinin nispi su içeriği (%).…………...…...42 Çizelge 4.11. Havuç (6, 9, 12, 13) genotiplerinin prolin seviyeleri (nmol g-1 YA).……….44
Çizelge 4.12. Havuç (6, 9, 12, 13) genotiplerinin gövde MDA düzeyleri (nmol g-1 YA)………..47
Çizelge 4.13. Havuç (6, 9, 12, 13) genotiplerinin EC değerleri (%)………...49 Çizelge 4.14. B noksan ve B toksik koĢullarda yetiĢtirilen havuç (6, 9, 12, 13)
genotiplerinin OH- ve H2O2 değerlerine ait varyans analiz sonuçları………50
Çizelge 4.15. Havuç (6, 9, 12, 13) genotiplerinin gövde ·OH
süpürülme aktivitesi (nmol g-1 YA)………...52
Çizelge 4.16. Havuç (6, 9, 12, 13) genotiplerinin gövde H2O2 süpürülme aktivitesi (nmol g-1 YA)………...55
Çizelge 4.17. B noksan ve B toksik koĢullarda yetiĢtirilen havuç (6, 9, 12, 13)
genotiplerinin SOD, POX, APOX, CAT, GR ve GSH değerlerine ait varyans analiz sonuçları………...………56
Çizelge 4.18. Havuç (6, 9, 12, 13) genotiplerinin gövde SOD aktiviteleri (Ünite mg-1 protein).………... 58
Çizelge 4.19. Havuç (6, 9, 12, 13) genotiplerinin gövde POX aktiviteleri (Ünite mg-1 protein).…………..………...60
Çizelge 4.20. Havuç (6, 9, 12, 13) genotiplerinin gövde CAT aktiviteleri (Ünite mg-1 protein)………...62
xii
Çizelge 4.21. Havuç (6, 9, 12, 13) genotiplerinin gövde GR aktiviteleri (Ünite mg-1 protein)……….64
Çizelge 4.22. Havuç (6, 9, 12, 13) genotiplerinin gövde APX aktiviteleri (Ünite mg-1 protein). ………...……66
Çizelge 4.23. Havuç (6, 9, 12, 13) genotiplerinin gövde GSH aktiviteleri (Ünite mg-1 protein). ………..……….69
xiii
ġEKĠLLER DĠZĠNĠ
ġekil 2.1. Antioksidan tarafından serbest radikalin nötralize edilmesi………...10 ġekil 3.1. AraĢtırmanın yapıldığı bitki fizyoloji laboratuvarı, tam kontrollü bitki
yetiĢtirme kabini ve hidroponik sistem………...14
ġekil 3.2. Deneme planı………..15 ġekil 3.3. Sterilizasyon ve çimlendirme aĢamaları ile ilgili görseller (a,b,c,d,e )……...16 ġekil 3.4. Bitkilerin kabine ekimi ve hasata kadar geliĢimi ile ilgili görseller
(a,b,c,d,e)……….………16
ġekil 4.1. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin gövde yaĢ
ağırlık değerleri (g bitki-1
)………...25
ġekil 4.2. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin kök yaĢ
ağırlık değerleri (g bitki-1
).………...27
ġekil 4.3. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin gövde
kuru ağırlık değerleri (g bitki-1
)………...29
ġekil 4.4. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin kök kuru
ağırlık değerleri (g bitki-1)………...………...31
ġekil 4.5. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin gövde boy
değerleri (cm)………...………...33
ġekil 4.6. Kontrol (C), B noksan ve B toksik koĢullarda havuç (6, 9, 12, 13)
genotiplerinin kök boy değerleri (cm)………....……….35
ġekil 4.7. 6. ve 9. havuç genotiplerinin kontrol (C) , B noksan ve B toksik koĢullarda bitlilerin tepkilerini gösteren resimler………..………...36
ġekil 4.8. 12. ve 13. havuç genotiplerinin kontrol (C) , B noksan ve B toksik koĢullarda
bitlilerin tepkilerini gösteren resimler……….………36
ġekil 4.9. 6. genotipte; kontrol (C) ve B uygulamalarına ait görseller [a (C x 0 mM B), b
(C x 0,5 mM B), c (C x 1 mM B), d (C x 2 mM B)]……….………..37
ġekil 4.10. 9. genotipte; kontrol (C) ve B uygulamalarına ait görseller [a (C x 0 mM B),
b (C x 0,5 mM B), c (C x 1 mM B), d (C x 2 mM B)]………37
ġekil 4.11. 12. genotipte; kontrol (C) ve B uygulamalarına ait görseller [a (C x 0 mM
B), b (C x 0,5 mM B), c (C x 1 mM B), d (C x 2 mM B)].………...37
ġekil 4.12. 13. genotipte; kontrol (C) ve B uygulamalarına ait görseller [a (C x 0 mM
xiv
ġekil 4.13. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin gövde B
konsantrasyonları (mg kg-1)……….39
ġekil 4.14. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin bağıl su
içeriği (%)………42
ġekil 4.15. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin gövde
prolin değerleri (nmol g-1
YA)……….44
ġekil 4.16. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin gövde
MDA düzeyleri (nmol g-1 YA)………...47
ġekil 4.17. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin EC
değerleri (%)………...49
ġekil 4.18. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin ·OH -radikali değerleri (nmol g-1
YA)………..52
ġekil 4.19. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin H2O2 değerleri (nmol g-1
YA)………...55
ġekil 4.20. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin SOD
activite değerleri (Ünite mg-1
protein)………...58
ġekil 4.21. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin POX
activite değerleri (Ünite mg-1
protein)………...60
ġekil 4.22. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin CAT
activite değerleri (Ünite mg-1 protein)……….62
ġekil 4.23. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin GR
activite değerleri (Ünite mg-1
protein)………...64
ġekil 4.24. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin APX
activite değerleri (Ünite mg-1
protein)………...66
ġekil 4.25. Kontrol (C), B noksan ve B toksik koĢullarda havuç genotiplerinin GSH
activite değerleri (Ünite mg-1
1. GĠRĠġ
Bitkiler büyüyüp geliĢebilmek ve yaĢamlarını idame ettirebilmek için temelde besin elementlerine gereksinim duymaktadır. Bitkilerdeki miktarlarına göre besin elementleri genel olarak “makro” ve “mikro” olmak üzere ikiye ayrılmaktadır. Mikro elementler, bitki bileĢimlerinde ve topraklarda makro elementlere oranla daha küçük konsantrasyonlarda bulunmasına rağmen (Bolat ve Kara, 2017) bitkiler için büyük önem taĢımaktadır. Son yıllarda yapılan çalıĢmalarda, dünya ve Türkiye topraklarında mikro besin elementleriyle ilgili beslenme problemlerinin yaygınlık gösterdiği ortaya konulmuĢtur. Mikro elementlerin hem fazlalığı hem de noksanlığı bitki geliĢimini sınırlandırmaktadır. Bor (B) bitkilerin eser miktarda da olsa gereksinim duydukları bir mikro elementtir. Türkiye, dünya B rezervlerinin %72’sine sahiptir. Bölgeden bölgeye topraklarda bulunan besin element miktarları değiĢiklik göstermektedir. Orta-Güney Anadolu tarım bölgesi topraklarının (Konya, Afyon, Karaman, Aksaray, Niğde, NevĢehir ve Kayseri) %26’sında B noksanlığı ve %18’inde ise bazı bitkiler için B toksisitesi mevcut olduğu bildirilmiĢtir (Gezgin ve ark., 2002). Bitkilerde stres faktörlerinin en önemlilerinden biri olan mikro element eksikliği veya toksisitesi; metabolizmanın iĢlevini engellemekte ve bitkide hasarlara neden olabilmektedir. Canlılar, doğal ortamlarında meydana gelen çevresel farklılıklara karĢı çeĢitli içsel ve mekanik tepkiler gösterirler (Edreva, 1998). Herhangi bir stres faktörü ile karĢılaĢan bitkiler biyokimyasal ve fizyolojik olarak çeĢitli tepkiler oluĢturdukları gibi, B fazlalığı ve B noksanlığı ile baĢ edebilmek için bazı biyokimyasal ve moleküler mekanizmalarını da harekete geçirebilmektedirler. Abiyotik stres faktörleri ve genlerin fizyolojik etkileri; hücresel metabolizmada değiĢimler, büyüme oranlarında ve ürün miktarlarında farlılıklara neden olurlar (Bray, 2000). Bundan dolayı, oluĢan stres koĢullarına karĢı dayanıklı bitki türlerinin belirlenmesi, tolerans mekanizmalarının açıklanması, bitkilerin B noksan ve B toksisitesine karĢı dayanıklılığını arttıran veya etkileyen unsurların belirlenmesi, giderek artıĢ sağlayan dünya nüfusuna, yeterli ve kaliteli ürünler sunabilme açısından önemli rol oynayacaktır.
Bitkisel üretim için tercih edilen genotiplerin abiyotik stres koĢullarına karĢı tolerans seviyelerinin yetersiz olması bitkilerde birçok metabolik olayı olumsuz yönde etkileyerek, ürün kalitesi ve verim kayıplarına sebep olmaktadır. Kültür bitkilerinin, abiyotik stres koĢullarına karĢı farklı seviyelerde dayanıklılık gösteren genotiplere sahip
oldukları bilinmektedir. Havuç, maydanozgiller (Umbelliferae) familyasına mensup
Daucus carota L. türüne ait kökleri etli (yumru) kültür bitkisidir. Besin değerlerinin
yüksek olması, çeĢitli mineralleri bünyesinde barındırması ve antioksidan özelliklere sahip olması nedeniyle dünyada ve Türkiye’de yaygın olarak tüketilmektedir. Havuç bitkisi, sofralık olarak tüketildiği gibi (A vitamini kaynağı olarak), kozmetik sanayinde (vücut ve güneĢ yağı), havuç konservesi, havuç dondurması, havuç reçeli ve havuç suyu olarak birçok alanda değerlendirilmektedir. Havuç, üretildiği alan, üretim miktarı, aynı zamanda pazar değeri açısından dünyada önemli ilk 10 sebze arasında yer almaktadır (Rubatzky ve ark., 1999). TÜĠK verilerine bakıldığında, Türkiye’de havuç üretimi yıllara göre artıĢ sağlamıĢtır. 2016 yılında 554.7 ton olan havuç üretimi 2017 yılında 569.5 ve 2018 yılında 642.8 tona ulaĢmıĢtır (Tüik, 2019). Konya KaĢınhanı bölgesi havuç üretimi açısından Türkiye’de ilk sıralarda yer almaktadır.
Bitkisel üretim yapılırken birçok stres etmeniyle karĢılaĢılmaktadır. Bu streslerden biri element noksanlığı ve toksisitesidir. Yukarıda da belirtildiği gibi B noksanlığı ve B toksisitesi bitki geliĢimini olumsuz yönde etkileyerek hem üretimi sınırlandırmakta hem de ürün kalitesini düĢürmektedir. Çevresel strese karĢı toleransı da kapsayan birçok hücresel süreçte reaktif oksijen türleri (ROT) ikincil haberciler olarak da rol oynamaktadırlar. Reaktif oksijen türleri, oksijenin elektron transferi esnasında fazla enerji kullanarak reaktif olan singlet oksijeni ve süperoksit radikali olarak kloroplastlarda üretilir. Hidrojen peroksit ve hidroksil radikalleri, süperoksitin reaksiyonlarının bir sonucu olarak oluĢabilir. Bütün bu reaktif türler potansiyel olarak zararlıdır. Bitkilerde özellikle abiyotik stres altında ROT üretiminin artıĢı lipitlerin peroksidasyonuna, proteinlerin oksidasyonuna, nükleik asit hasarına, enzim inhibisyonuna ve hücrelerin ölümüne kadar birçok hasara yol açabilir. ROT normalde kloroplastlarda ve diğer hücre altı kompartmanlarında bulunan çeĢitli antioksidanlar ve enzimler tarafından temizlenirler. Antioksidanlar düĢük konsantrasyonlarda oksidasyon yapabilen ve diğer bir substratın oksidasyonunu azaltan veya engelleyen yani oksidasyona karĢı mücadele eden maddelerdir (Smirnoff, 1993; Büyük ve ark., 2012).
Havuçta ve diğer tüm bitkilerde yüksek verim ve kalite için, bor stresine karĢı bitkinin direncinin arttırılması gerekmektedir. B stresine karĢı direncin arttırılabilmesi için bitkinin büyümesi sırasında bu stres faktörlerinin meydana getirdiği fizyolojik ve biyokimyasal değiĢimlerin bilinmesi ve beraber yorumlanması zorunluluktur. B stresine
dayanıklı bitki türlerinin belirlenmesi ve geliĢtirilmesi, tolerans mekanizmaları ve bunda etkin rol oynayan unsurların açıklanması ile mümkün olacaktır. Tezin amacı; elde edilen sonuçlar doğrultusunda bor problemi olan alanlara uyum sağlayabilen havuç çeĢitlerinin belirlenmesi ve geliĢtirilmesidir. B noksan ve B toksik koĢullara maruz bırakılan havuç genotiplerinin geliĢtirdikleri savunma mekanizmaları üzerine etkileri, belirli büyüme evresinde yapılan örneklemelerle ele alınmıĢ ve temel büyüme parametreleri, gövde B analizleri, iyon sızıntısı (EC) ve malondialdehit (MDA) miktarındaki değiĢiklikler yardımıyla hücresel membranlardaki hasar oranı belirlenmiĢtir. Ayrıca, süperoksit dismutaz (SOD), askorbat peroksidaz (APX), katalaz (CAT), peroksidaz (POX) ve glutatyon redüktaz (GR) gibi antioksidan enzim aktiviteleri, glutatyon (GSH) miktarı ve hidrojen peroksit (H2O2) ile hidroksil (.OH-) radikallerinin süpürülme miktarları belirlenmek suretiyle havuç genotiplerindeki tepkiler ortaya konulmaya çalıĢılmıĢtır.
2. KAYNAK ARAġTIRMASI
Havuç, maydonozgiller (Umbelliferae) familyasına mensup Daucus carota L. türüne giren kökleri etli (yumru) kültür bitkisidir. Daucus carota’nın 13 alt türünün olduğu öne sürülmüĢtür (Simon ve Goldman, 2006). Daucus carota’nın alt türlerden on ikisi tanesi yabani formda olup geriye kalan biri ise kültür havucudur. Havuç mor, siyah, kırmızı, beyaz ve sarı çeĢitler mevcut olmasına rağmen, genellikle turuncu renkli bir kök sebzesidir.
Havuç, Daucus carota'nın Avrupa'ya Güneybatı Asya'ya özgü yerli biçimidir. Kültür havuçları Doğu Asya ve Batı olmak üzere iki farklı kısımda yetiĢmektedir. Doğu Asya havuçları erken çiçeklenme eğilimi gösteren, antosiyanin içeren, kırmızımtırak mor veya sarı renklerde olup, genellikle dallı köklere sahiptirler. Bu havuçlarda yaygın olarak görülen mor renk antosiyanin pigmentlerinden gelmektedir. Batı havuçları ise uzun süreli düĢük sıcaklığa maruz kalmadıklarından çiçeklenmeye daha az eğilim gösterirler. Batı havuçları kök tipleri bakımından kendi içlerinde dörde ayrılırlar; turuncu, sarı, kırmızı veya beyaz kök renklerine sahiptirler (Rubatzky ve ark., 1999). Nantes havuçları karoten bileĢiğinden dolayı turuncu renktedirler. Beyaz kök rengine sahip olan havuçlar karoten nedeniyle normal turuncu pigmentten yoksundur.
Bazı yazılı belgelere göre kırmızı, mor ve sarı havuçlar 10. yy’da Kuzey Arabistan’da ve Ġran’da, 11. yy’da Suriye’de, 12. yy’da Ġspanya’da, 13. yy’da Ġtalya’da, 14. yy’da Fransa, Almanya ve Hollanda’da ve 15. yy’da Ġngiltere’de yetiĢtirilmeye baĢlandığı öne sürülmektedir. Havuç 13. yy’ın sonlarına doğru Çin’e, 17. yy’da Avrupalı tüccarlar yoluyla Amerika’ya gitmiĢtir. Çin’de de ilk olarak sarı ve mor renkli havuçlar yetiĢtirilmiĢtir (Banga, 1963; Rubatzky ve ark., 1999; Simon, 2000). Yüzyıllar ve kıtalar boyunca yaptığı yolculukta, sayısız botanikçi eski havuçların kompozisyonunu, görünüĢünü, tadını ve büyüklüğünü geliĢtirmeyi baĢarmıĢ ve 17. yüzyılda ilk Hollanda'da ortaya çıkan modern turuncu renkli havuç üretilmiĢtir (Anonim).
Havuç, tüm dünyada en popüler gıda bileĢenlerinden biri olarak, yaygın kullanılan bir kök sebzesidir. Besin değerlerinin yüksek olması, beta-karoten, diyet lifi, antioksidanlar, minerallerin varlığı açısından; havuç üretildiği alan, üretim miktarı, aynı
zamanda pazar değerinin yüksek olmasından dolayı dünyada önemli ilk 10 sebze arasındadır (Simon, 2000).
2.1. Bor Elementi ve Bitkiler Üzerine Etkileri
Bitkiler büyüme ve geliĢim süreçlerini tamamlayabilmek için birtakım besin elementlerine gereksinim duymaktadır. Mineral olan besin elementleri makro ve mikro olmak üzere iki gruba ayrılırlar. B değiĢken ölçülerde, birçok bitkinin ihtiyaç duyduğu mikro besin elementidir. Bütün bitkiler normal geliĢmeleri için bir miktar bora ihtiyaç duyarlar (DemirbaĢ, 2005).
Yeryüzünde yaygın olan elementler arasında yer alan B toprakta, kayalarda ve suda yaygın olarak bulunan bir elementtir. Yüksek konsantrasyonda ve ekonomik düzeydeki B kaynakları, B’nin oksijenle bağlanmıĢ bileĢikleri olarak daha çok Türkiye (rezervlerinin yaklaĢık %70’i) ve Amerika'nın kurak, volkanik ve hidrotermal aktivitelerinin yüksek olduğu bölgelerde bulunmaktadır (Ediz ve Özdağ, 2001). Bor; deri renklendirici, böcek öldürücüler, elektronik rafinasyon, kozmetik, ilaç, çimento korozyon önleyiciler, yapıĢtırıcılar, dezenfekte ediciler, naylon, gübre, nükleer yakıt teknolojisi, tıp sanayi, antiseptikler, borik asit üretimi, fotoğrafçılık, boya, tekstil, ateĢe dayanıklı olan malzeme, deterjan sanayinde, kaplama sanayinde, cam sanayisi gibi birçok alanda kullanılmaktadır.
Bitkilerin büyüme ve geliĢmesinde B’nin önemli bir element olduğu ilk olarak Warington tarafından kanıtlanmıĢtır (Ho, 2000). Yapılan bazı çalıĢmalarda, dünya ve Türkiye topraklarında mikro besin elementleriyle ilgili beslenme problemlerinin yaygınlık gösterdiği ortaya konulmuĢtur (Gezgin ve ark., 2002). Türkiye tarım arazilerinin yaklaĢık %25’inde B noksanlığı gözlemlenirken, %20’sinin üzerinde ise B toksisitesi görülmektedir (Gezgin ve ark., 2002). Mutlak gerekli besin elementlerinden birisi olan B’a karĢı tepkilerde bitkiler arasında büyük faklılıklar olduğu (Gezgin ve ark., 2005), B noksanlığı koĢullarında, B uygulamasına sebzeler içerisinde en fazla olumlu tepkinin ise havuç tarafından gösterildiği bildirilmiĢtir (Sakal ve Singh, 1995).
Bitkilerde, B’nin büyüme ve geliĢmesindeki önemi fizyolojik ve biyokimyasal olaylarda yer almasından kaynaklanmaktadır (Cakmak ve Römheld, 1997). B, bitki fizyolojisinde hücre duvarının dayanıklılığında, fenol metabolizmasında, karbonhidratların taĢınmasında, solunumda, indol asetik asit metabolizmasında, membran bütünlüğünde ve generatif organların oluĢumunda, polen çimlenmesinde ve polen tüpü büyümesinde rolü önemlidir (Parr ve Loughman, 1983; Marschner, 1995).
Topraktaki B, iletim demetli bitkilerde köklerden transpirasyonal akıntı ile gövde ve yaprakların büyümekte olan bölgelerine taĢınır ve buralarda biriktirilir. B bitki köklerine yüksüz haldeki borik asit Ģeklinde girer (Woods, 1996). Bitkiler temelde B’yi borik asit B(OH)3 olarak pasif absorbsiyon, borat iyonları B(OH)4- Ģeklinde ise aktif absorbsiyon yoluyla alırlar (Kacar ve Katkat, 2010).
Stres, önemli fizyolojik ve metabolik değiĢimlere yol açarak bitkilerde büyüme ve geliĢmeyi olumsuz Ģekilde etkilerken, ürün kalitesinin ve miktarının azalmasına, bitkinin veya organlarının ölümüne yol açabilmektedir. Bitkiler, bulundukları çevrelerde yaĢamlarını ve geliĢme oranlarını kısıtlayıcı bazı stres faktörlerine maruz kalabilmektedir. Çevresel stres faktörleri; gen(ler)in fizyolojik etkileri ile hücresel metabolizma değiĢimlerinin, büyüme oranları ve ürün miktarlarının değiĢimine kadar çok çeĢitli tepkilere neden olurlar (Bray, 2000). Bitkilerde stres faktörlerinin en önemlilerinden biri olan besin elementlerinin eksikliği veya toksisitesidir. Bitkiler tarafından eser miktarda gereksinim duyulan, eksiklik ile toksisite arasındaki eĢiğin dar olduğu bir mikro besin maddesi olarak B benzersiz bir elementtir (Brown ve ark., 2002; Yau ve Ryan, 2008). B’nin noksanlık ve toksisitesi belirtileri bitkilerde yaygın ve belirgin olarak görülmektedir (Gezgin ve ark., 2005).
Mahsul verimliliğini sınırlayan önemli bir tarımsal sorun olan B toksisitesi dünyanın farklı bölgelerinde; B bakımından zengin topraklarda veya sulama sularında veya tutarsız gübre uygulamaları sonucunda topraklarda bor birikimi sonucunda meydana gelebilir. B toksisitesi, kurak ve yarı kurak ortamlardaki topraklarda bitki büyümesini sınırlandırabilen önemli bir sorundur (Nable ve ark., 1997).
B toksisitesi sonucunda bitkilerde kuru madde kaybı, meyve çürümesi, yapraklarda öncelikle uç ve kenarlarda oluĢan kahverengi lekeler ilerleyen aĢamalarda nekroza dönüĢümü gibi bozulmalar meydana gelir (Brown ve Shelp, 1997).
B eksikliği, birçok mahsülün verimliliğini önemli derecede etkileyen en belirgin mikro besin elementidir. B eksikliği durumunda bitkide büyümenin bozulması, büyüyen meristemlerin ölümü ve damar demetlerinin geliĢiminin engellenmesi gibi semptomlara neden olur. Bu semptomlar, B'nin hücre duvarı yapısındaki önemli rolünün ve bitkinin floemindeki ara mobiliteye sahip olması sonuçlarındandır (Marschner, 1995; Brown ve Shelp, 1997; Brown ve ark., 2002).
B eksikliğinin belirgin ilk görsel semptomları köklerdeki azalmalardır: ana kök uzunluğu ve primer kökün uzunluğu baĢına lateral kök sayısı artmasıdır. B eksikliği olan kökler daha kısa, daha kalın ve açık kahverengi renkte olur. Yapraklarda tipik B eksikliği belirtileri ilk önce genç yapraklarda ortaya çıkar ve yüzey alanında koyu yeĢil renk oluĢur ve yaprak lamina kalınlığı artar. Yapraklar, özellikle en küçük olan genç yapraklar, düzensiz olarak yaprak kenar boĢlukları kıvrılma gösterir. Ġlerleyen aĢamalarda ise semptomlar daha da belirginleĢir, genç yapraklar dıĢındaki yapraklarda da noksanlık belirtileri görülebilir (Hajiboland ve Farhanghi, 2010).
Torun ve ark. (1999), Konya koĢullarında yetiĢtirilen 22 buğday çeĢidinin B toksisitesine duyarlılığını sera ve tarla koĢullarında yaptıkları araĢtırmada B toksisitesinden kaynaklanan verim kayıplarının büyük boyutlarda olduğu bildirilmiĢtir. B’den kaynaklanan verim kayıplarının ekmeklik çeĢitlerle karĢılaĢtırıldığında, makarnalık buğdaylarda oldukça fazla olduğu görülmüĢ, kuru madde veriminde azalmaların makarnalık buğday çeĢitlerinde farklı olmamasına karĢılık ekmeklik buğday çeĢitleri arasında oldukça farklı olduğu saptanmıĢtır. Topraktaki B fazlalığında bitkilerin farklı reaksiyon gösterme nedenlerinin, çeĢitten çeĢide farklılık göstermesinden kaynaklı olduğu saptanmıĢtır.
Günes ve Alpaslan (2000), yaptıkları araĢtırmada 8 tane mısır çeĢidinin B toksisitesine duyarlılıklarını belirlemek amacıyla yürüttükleri çalıĢmada, toprağa 0, 10 ve 30 mg kg-1 düzeylerinde H3BO3 formunda B uygulamıĢlardır. ÇalıĢma sonunda bitkilerin yaĢ ve kuru ağırlıkları ile B konsantrasyonları belirlenmiĢtir. YaĢ ve kuru bitki
ağırlıkları ile bitkilerin B konsantrasyonları arasındaki iliĢkilerden yararlanılarak mısır çeĢitlerinin B toksisitesine duyarlılıkları ortaya konulmuĢtur. Genel olarak yüksek bora duyarlılıkları düĢük olan çeĢitlerin, yüksek olan çeĢitlere göre daha fazla B içerdikleri belirlenmiĢtir.
Hamurcu ve ark. (2016b), B toksisitesine toleranslı (RX 770) ve hassas (TTM 81.19) mısır çeĢitlerine 1/5 oranında seyreltilmiĢ hoagland çözeltisi ile kontrol, 2.5, 25, 50 ve 100 mg L-1 B uygulayarak kök ve gövde anatomisindeki etkileri belirlemek amacıyla yürüttükleri çalıĢmada, bitkide gövde ve kök B konsantrasyonları artan B dozlarına bağlı olarak artıĢ gösterdiği saptanmıĢtır. Mısır çeĢitleri kendi aralarında değerlendirildiğinde, artıĢların toleranslı RX 770 çeĢidinde TTM 81.19 çeĢidinden daha yüksek olduğu belirlenmiĢtir. Elde edilen sonuçlarda kontrole göre B uygulamalarında her iki mısır çeĢidinde de kök metaksilem, gövde trake ve korteks çaplarının önemli düzeylerde azaldığı, bu azalmaların B toksitesine toleranslı RX 770 çeĢidinde, hassas TTM 81.19 çeĢidine göre daha fazla olduğu belirlenmiĢtir.
Harite (2008), yapmıĢ olduğu çalıĢmada perlit kum karıĢımında yetiĢtirilen pamuk çeĢitlerinin farklı düzeylerde uygulanan B (0.5, 7.5, 15, 22.5 mg L-1
B ) dozlarına karĢı olan reaksiyonları incelemiĢtir. ÇalıĢmada 8 pamuk çeĢidi kullanılmıĢtır. B uygulama dozlarına paralel olarak B toksisitesinden zarar görmüĢ yaprak sayısı, kök, gövde ve yaprak B içerikleri artmıĢ buna karĢın bitkilerde taze ağırlık, kuru ağırlık, boy ve yaprak sayıları azaldığı bildirilmiĢtir.
Lordkaew ve ark. (2011), tarafından yürütülen çalıĢmada dane olgunlaĢma döneminde B uygulaması yapılmadığı durumlara göre, B uygulamasının yapıldığı durumlarda dane verimi ve koçan baĢına tane sayısı, üzerine daha fazla bir etkisinin olduğu bildirilmiĢtir. Bitki baĢına dane verimi B0 uygulamasında 0.5 g, B20 uygulamasında 72.3 g olduğu tespit edilmiĢtir. Sap, kök ve bitki boyunda gerçekleĢen farlılıkların dane verimi ve koçan baĢına tane sayısı kadar belirgin olmadığı belirlenmiĢtir.
Karabal ve ark. (2003), B toksisitesine duyarlı (Hamidiye) ve toleranslı (Anadolu) arpa çeĢitleri 8 gün boyunca topraksız ortamda fideler halinde yetiĢtirmiĢlerdir. Fideler, 5 gün boyunca, 5 ve 10 mM borik asit iĢlemlerine tabi
tutulmuĢtur. Uygulamalar kontrole göre kıyaslandığında büyüme parametre (kuru-yaĢ ağırlık) değerlerinde azalmalar meydana geldiği belirtilmiĢtir.
Hamurcu ve ark. (2006), bezelye bitkisine, kontrollü sera koĢullarda farklı seviyelerde B (0, 1, 20 mg kg-1 B) ve Fe (0, 6, 60 mg kg-1 Fe) dozları uygulamıĢtır. B toksisitesinden kaynaklanan kuru madde miktarının uygulanan B dozlarının ortalaması dikkate alındığında B uygulanmayan saksılara göre azaldığı, 1 mg kg-1 B uygulamasında ise artıĢ olduğu belirlenmiĢtir. Bezelye bitkisine uygulanan B miktarındaki artıĢla birlikte, bitki boyunda da artıĢlar gözlemlenmiĢtir. Bitkiye uygulanan B miktarı arttıkça içsel B konsantrasyonunun arttığı belirlenmiĢtir.
Omay (2019), tam kontrollü sera koĢullarında yürüttüğü çalıĢmada, ekmeklik buğday çeĢit ve hatlarına artan dozlarda B (0, 0.75, 1.5 mM H3BO3) ve tuz (0, 100 mM NaCI) uygulamaları yapmıĢtır. Artan B dozlarına bağlı olarak, tüm genotiplerin B konsantrasyonlarında ve EC içeriklerinde artıĢ, kuru ağırlık ve bitki boylarında ise azalma meydana geldiğini bildirmiĢtir.
2.2. Bitkilerde Bor Stresi ve Savunma Sistemleri
Bitkiler diğer streslerde olduğu gibi B stresiyle de baĢ edebilmek için bazı mekanizmalar geliĢtirmiĢlerdir. Bu geliĢtirmiĢ oldukları mekanizmalar fizyolojik ve biyokimyasal olmak üzere ikiye ayrılır. Biyokimyasal mekanizmalar; seçici iyon birikimi veya dıĢlanması, köklerle alınan iyonların kontrolü ve yapraklara taĢınımı, tüm hücre düzeyinde veya hücresel boyutta iyonların dağılımı, uyumlu bileĢiklerin sentezi, fotosentetik yolun değiĢmesi, membran yapısındaki değiĢimler, antioksidan enzimlerin ve bitki hormonlarının indüksiyonu olarak sıralanabilirler (Seckin ve ark., 2009).
Stresin Ģiddeti, süresi, stresten etkilenmiĢ olan bitkilerin geliĢim aĢaması, doku tipi ve birçok stresin etkileĢimlerine göre bitkilerin gösterdikleri tepkiler farklı farklı olmaktadır (Koca ve ark., 2006). Bitki dokularında stres faktörlerinin azaltılması veya önlenmesine yönelik tepkiler, bitkilerin streslere karĢı oluĢturdukları mekanizmalar ile mümkün olmaktadır. Bitkilerde strese tepki olarak; yaprak ayasının kalınlığı, stomaların büyüklüğü ve sıklığı ve kütikulanın kalınlığı değiĢmektedir (Acar ve ark., 2001).
Stressiz koĢullarda, bitkilerde yer alan antioksidan savunma sistemleri, reaktif oksijen türlerine (ROT) karĢı gerekli korunmayı sağlamaktadırlar (Cakmak ve ark., 1993). Fakat yaĢamsal reaksiyonların sonucu veya oluĢan herhangi bir stres faktörünün ortaya çıktığı ve koruyucu mekanizmaların da iĢlev görmediği durumlarda, bitki dokularına zarar veren aktif oksijen türlerinin üretimi artmaktadır. Bunun sonucunda aktif oksijen türlerinin, bitkiler veya herhangi bir aerobik organizmanın, hücre yapısına ve iĢlevlerine zarar verebildiği bildirilmiĢtir (Sairam ve ark., 2002; Demiral, 2003)
Oksijen bitkiler için temel bileĢenlerden biridir. Moleküler oksijenin (O2) suya (H2O) indirgenmesi ile bitkiler için enerji kaynağı oluĢmaktadır. Oksijenin suya indirgenememesi halinde ise ROT oluĢmaktadır (Seçkin, 2005; Çağlar, 2012). Reaktif oksijen türleri; hidrojen peroksit (H2O2), süperoksit (O2-) radikalleri, hidroksil radikalleri (.OH-) ve tek değerlikli oksijendir (O2). Bunlar oksidatif reaksiyonlarla bitki hücrelerinde kloroplastlarda, mitokondrilerde ve peroksizomlarda üretilebilmektedirler. Bu radikaller membran proteinleri ve lipidler için çok toksiktir, hücresel zarların parçalanmasına ve dengesizleĢmesine vede zar sızıntısına neden olur (Apel ve Hirt, 2004).
Stres altında bitkiler yaĢamlarını devam ettirebilmek ve stresle baĢa çıkabilmek için ROT’un kontrolünü veya kendi bünyesinden temizlenmesini sağlayan çeĢitli antioksidanlara sahiptirler (Büyük ve ark., 2012). Korunma amacıyla ROT’lara karĢı, bitkiler bünyelerinde süperoksit dismutaz (SOD), katalaz (CAT), askorbat peroksidaz (APX), glutatyon redüktaz (GR) veya peroksidaz (POX) gibi antioksidan enzimler ve çeĢitli enzimatik olmayan antioksidanlar (alfa-tokoferol, glutatyon, askorbat vb.) bulundururlar (Matthews ve ark., 1984; Mittler, 2002; Blokhina ve ark., 2003).
ġekil 2.1. Antioksidan tarafından serbest radikalin nötralize edilmesi (Büyük ve ark.,
Liu ve Yang (2000), düĢük miktarlarda B’nin soya fasulyesinde SOD, APX ve CAT aktivitelerini azalttığı bildirmiĢlerdir. Narenciye fidelerinde ise B eksikliğinin APX aktivitesini arttırdığı ancak CAT aktivitesini azalttığı bildirilmiĢtir (Han ve ark., 2008).
Dube ve ark. (2000), ayçiçeği ( Helianthus annuus ) bitkisi farklı iki B dozunda (0.017 ila 0.66 mg L-1 B) kum ortamında yetiĢtirilmiĢtir. B eksikliği (0.017 mg L-1 B) durumunda bitkideki APX aktivite değerlerini arttırdığı, ancak CAT'ı düĢürdüğü ve SOD aktivitesini etkilemediği bildirilmiĢtir.
Farklı bir çalıĢmada, materyal olarak kullanılan ayçiçeği bitkisinde glutatyon ve glutatyon redüktaz seviyelerinin B eksikliğine cevap olarak azaldığı belirlenmiĢtir (Cakmak ve Römheld, 1997).
Cervilla ve ark. (2007), yaptıkları çalıĢmada 0.5 ve 2.0 mM B'de yetiĢtirilen domates bitkisindeki Kosaco çeĢidinde (kontrole kıyasla) SOD aktivitesinde bir artıĢ gözlemlemiĢlerdir. Buna karĢılık, aynı durumda yetiĢen Josefina kültür bitkileri, yalnızca 0.5 mM B'da SOD aktivitesinde bir yükselme, 2.0 mM B konsantrasyonunda ise kontrole kıyasla SOD aktivitesinde önemli ölçüde bir değiĢme olmadığı bildirilmiĢtir. Ayrıca turunçgillerde, hem B eksikliği hem de fazlalığında SOD aktivitesinde bir artıĢ gerçekleĢtiğini belirlemiĢlerdir (Han ve ark., 2008).
Hajiboland ve Farhanghi (2010) tarafından, Ģalgam (Brassica rapa L.) bitkileri yeterli (25 μmol L−1 B) veya düĢük (<2.5 μmol L−1 B) B konsantrasyonlarında 1 ay boyunca besin çözeltisinde yetiĢtirilmiĢtir. Antioksidan, savunma enzimlerin aktivitesi B eksikliği altında yüksek olmasına rağmen, daha yüksek malondialdehit (MDA) birikimi üzere, bu enzimler oksidatif (H2O2) hasardan yeterli koruma sağlayamadığı ve dokularda sızıntı olduğu bildirilmiĢtir. Radikaller membran proteinleri ve lipidler için çok toksiktir ve hücresel zarların parçalanmasına ve dengesizleĢmesine neden olur ve zar sızıntısına neden olur (Apel ve Hirt, 2004). Karabal ve ark. (2003), tarafından yürütülen çalıĢmada artan B dozuna (5 ve 10 mM B) bağlı olarak sürgünlerde MDA içerikleri ve elektrolit sızıntısında bir artıĢ olduğu belirlenmiĢtir. B eksik ortamda yetiĢtirilen ayçiçeği bitkisinde doymamıĢ yağ asitlerindeki azalmaya paralel olarak malondialdehit (MDA) içeriğinde artıĢ gözlemlenmiĢtir (El-Shintinawy, 2000).
Karabal ve ark. (2003), Hamidiye ve Anadolu arpa çeĢitlerinde, antioksidan tepkisi üzerine etkilerini araĢtırmıĢlardır. Her iki çeĢidinin sürgün dokusundaki toplam SOD, CAT ve GR aktivitelerinde anlamlı bir değiĢim gözlemlenmediği, fakat 10 mM borik asit iĢlemlerinde belirgin bir Ģekilde, kontrole göre yüksek APX aktiviteleri gözlemlendiği bildirilmiĢtir. Gunes ve ark. (2006) B stres altındaki marul ve asmada, güçlü bir CAT artıĢı ve B stresinde APX aktivitesinde eĢ zamanlı bir azalma olduğunu bildirmiĢlerdir.
ÇeĢitli bitkilerde (arpa, nohut gibi) bor toksisitesi altında GR aktivitesinde artıĢlar olduğu görülmüĢtür (Karabal ve ark., 2003; Ardıç ve ark., 2009).
Wang ve ark. (2011)’de armutta yaptıkları çalıĢmada 10 M B'de yetiĢtirilen kontrol bitkilerine kıyasla 100 M ve 300 M B'de APX ve CAT antioksidan enzim aktivitelerinde bir artıĢ olduğunu, 500 M B’de yetiĢtirilen bitkilerde ise daha düĢük APX ve CAT aktivitesi olduğu bildirmiĢlerdir. Aynı çalıĢmada B dozları MDA ve H2O2’nin yapraklarda arttığı görülmüĢtür.
Hamurcu ve ark. (2013), yaptıkları çalıĢmada soya fasulyesinde B toksisitesinin antioksidan tepkileri üzerine etkisini araĢtırmıĢlardır. Ġki farklı B dozu (2 mg kg−1 B ve 12 mg kg−1 B) uygulanmıĢtır. Yaprak B konsantrasyonlarında 12 mg kg −1 B uygulaması sonucunda SOD, CAT ve APX aktivitelerindeki önemli artıĢlar meydana geldiği, POX aktivite değerlerinin değiĢmediği, GR aktivitelerinin ise azaldığı gözlemlenmiĢtir. SOD ve GR aktiviteleri kontrol gruplarına göre 2 mg kg −1 B konsantrasyonda azalırken, POX aktivitesinin arttığı ve CAT aktivitesinin değiĢmediği bildirilmiĢtir.
Cervilla ve ark. (2007), domates bitkisinin B toksisitesine karĢı enzimatik tepkisini incelemek amacıyla yapraklardaki antioksidan enzimler analiz edilmiĢtir. Uygulanan bor dozlarına göre CAT ve APX enzim aktivitelerinin kontrole göre arttığı bildirilmiĢtir.
Esim ve ark. (2012), yaptıkları çalıĢmada mısır fidelerine uyguladıkları B dozları (2 mM ve 4 mM B) sonucunda POX aktivitesinin azaldığını belirlemiĢlerdir. Bu sonuca bağlı olarak B toksisitesi altında mısır bitkilerinde oksidatif stres meydana geldiği bildirilmiĢtir. Uğurlar (2019), yaptığı çalıĢmada Capia tipi kırmızı tatlı biber bitkisine kontrol (0.05 mM B) ve 2 mM B uygulamıĢtır. B toksisitesi sonucunda POX aktivite içeriğinin arttığı belirlenmiĢtir.
Ayçiçeği bitkisinde B toksisitesinin glutatyon sentezini inhibe ettiğini bildirmiĢlerdirRuiz ve ark. (2003). Armut bitkisinde B toksisitesi koĢullarında GSH seviyelerinin önce arttığı ve sonra azaldığı görülmüĢtür (Wang ve ark., 2011).
Eraslan ve ark. (2007), artan B seviyelerinde domates ve biber bitkilerinde, prolin birikiminin en yüksek B düzeyi ile arttığını bildirmiĢlerdir. Domates bitkisindeki prolin miktarının biber bitkisinden oran olarak daha yüksek bulunduğu belirlenmiĢtir. Gunes ve ark. (2006), Grapevinin (Vitis vinifera L.)’e artan dozlarda B (0, 10, 20 ve 30 mg kg-1 ) uygulamıĢlardır. Bu çalıĢmada B toksisitesi artıĢına bağlı olarak, prolin miktarının azaldığı bildirilmiĢtir. Farklı bir çalıĢmada ise B toksisitesinde (10 mM), hidroponik olarak yetiĢtirilen arpa fidelerinin yapraklarında ve köklerinde, prolin içeriğinde önemli değiĢikliklere neden olmadığı bildirilmiĢtir (Karabal ve ark., 2003).
Karabal ve ark. (2003), tarfından farklı arpa çeĢitlerindeki oluĢan elektrolit sızıntısı, sürgünlerde doza bağlı bir artıĢ sergilerken, kök dokularında önemli bir değiĢiklik göstermediği bildirilmiĢtir. B problemi varlığında yetiĢtirilen ayçiçeği bitkisinin yapraklarında tespit edilen %EC değerlerine bakıldığında kontrole göre artıĢlar olduğu gözlemlenmiĢtir (Yıldıztekin ve Tuna, 2015).
Mukhopadhyay ve Mondal (2015), yaptıkları araĢtırmada çay bitkilerinin (Kamelya sinensis (L.)), B yeterli bitkilere göre B eksikliğinde %11 ve %18 oranında RWC oranlarının azaldığı, strese bağlı olarak bitkinin su alımının azalmasından dolayıda turgor kaybının yaĢandığı bildirmiĢlerdir. Mısır (Zea mays L.) bitkisinde 0.05 ve 2 mM B’ye maruz bırakıldıklarında bitkilerde nispi su içeriğinin (RWC) azaldığı gözlemlenmiĢtir (Kaya ve ark., 2018).
3. MATERYAL VE YÖNTEM
3.1. Materyal
AraĢtırmada daha önceki yapılmıĢ ön çalıĢmalarda bor toksisitesine tepkileri belirlenmiĢ olan bor toksisitesine duyarlı 9 (Mor Havuç) ve 12 (Nantes-Turuncu
Havuç) ve bor toksisitesine toleranslı 6 (Mor Havuç) ve 13 (Nantes-Sarı Havuç)
numaralı genotipler kullanılmıĢtır. AraĢtırmada kullanılan bitki materyalleri Yalova Tarımsal AraĢtırma Enstitüsü’nden temin edilmiĢtir.
AraĢtırma materyali olan havuç, ekim nöbeti için uygun bir tür olabilmesi, hasat kolaylığı, kültürel iĢlemlerin büyük oranda makine ile yapılabilmesi, hastalık ve zararlılar açısından fazla sıkıntıya yol açmaması gibi nedenlerden dolayı son dönemlerde yetiĢtiriciliği yaygınlaĢan bir türdür.
3.2. Yöntem
Havuç genotipleri iklim odasında kontrollü koĢullarda su kültürü ortamında, yetiĢtirilmiĢtir. Gerekli ısı, nem, ıĢık ve ayrıca sterilizasyon kontrolleri yapılmıĢtır.
ġekil 3.1. AraĢtırmanın yapıldığı bitki fizyoloji laboratuvarı, tam kontrollü bitki
yetiĢtirme kabini ve hidroponik sistem
3.2.1. Deneme serilerinin hazırlanması
ÇalıĢmada havuç materyaline, kontrol (0.0033 mM B) grubuna göre, mukayese amaçlı 0 mM B, 0.5 mM B, 1 mM B, 2 mM B (H3BO3) dozları uygulanmıĢtır. ÇalıĢma her bir gurupta 4 tekerrür olacak Ģekilde kurulmuĢtur. Buna göre; çalıĢma 5 uygulama x 4 çeĢit x 4 tekerrür olarak toplam 80 saksı Ģeklinde yürütülmüĢtür. ÇalıĢmada B dozları
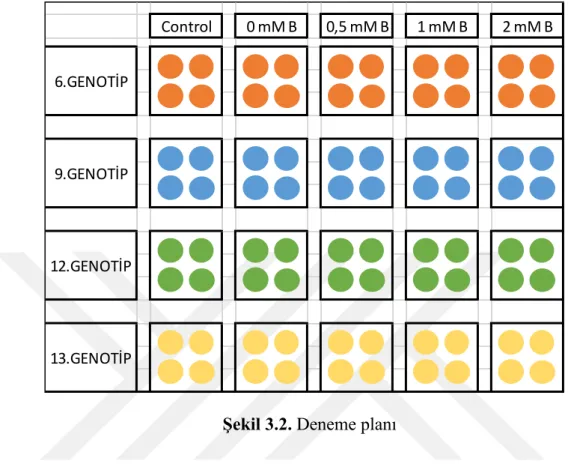
hoagland çözeltisi ile uygulanmıĢtır. Besin solüsyonları her üç günde bir yenisiyle değiĢtirilerek tazelenmiĢtir. Deneme planı aĢağıda özetlenmiĢtir.
Control 0 mM B 0,5 mM B 1 mM B 2 mM B 6.GENOTİP 9.GENOTİP 12.GENOTİP 13.GENOTİP
ġekil 3.2. Deneme planı
3.2.2. Tohum dikimi
ÇalıĢmada kullanılmıĢ olan havuç genotiplerinin tohumları önce %1’lik sodyum hipoklorit ile muamele edilmiĢ sonra steril saf suyla yıkanarak yüzey sterilizasyonu tamamlanmıĢtır. Steril hale getirilen tohumlar 2-3 saat saf suda bekletilmiĢtir. 6 gün boyunca karanlıkta 22°C’de iklim odasında çimlendirildikten sonra elde edilen fideler 1/5 oranında hoagland solüsyonu içeren (kontrol) steril edilmiĢ saksılarda her saksıda 5 bitki olacak ve her uygulamada 20 bitki olacak Ģekilde dikim iĢlemi gerçekleĢtirilmiĢtir. Genç fidecikler büyüme ve geliĢme süresince %45-55 nem, 16 saat aydınlık ve 8 saat karanlık fotoperyod, 21±1 0C sıcaklık ile 16000 Lüx/Gün ıĢık intensitesi olan Toprak Bilimi ve Bitki Besleme Bölümü bünyesinde bulunan Su Kültürü Laboratuvarı’nda kontrollü koĢullar altında yetiĢtirilmiĢtir.
ġekil 3.3. Sterilizasyon ve çimlendirme aĢamaları ile ilgili görseller (a,b,c,d,e )
ġekil 3.4. Bitkilerin kabine dikimi ve hasata kadarki geliĢimleri ile ilgili görseller
(a,b,c,d,e)
3.2.3. Örnekleme
Bitkiler ilk üç yapraklı evreye geldiklerinde uygulamaların ilk baĢlangıç günü kontrol (0.0033 mM B) grubuna göre mukayese amaçlı 0 mM B, 0.5 mM B, 1 mM B, 2 mM B uygulamaları ile beslenmeye baĢlanmıĢtır. Besin solüsyonları her üç günde bir yenisiyle değiĢtirilerek tazelenmiĢtir. Bitkiler B noksanlığı ve toksisitesi koĢullarında
a b c d e
e d
b
morfolojik değiĢimler göstermeye baĢladığında örneklemeler yapılmıĢtır. Örneklemelerde saksılardaki bitkilerden eĢit sayıda kökleriyle birlikte bütün bitki alınıp, kökler ayrıldıktan sonra yaprak örneklerinin bir kısmı fizyolojik analizlerde kullanılmıĢtır. Aynı zamanda yaprak örneklerinin geriye kalan kısmları enzim analizlerinde kullanılmak üzere -80 oC’de derin dondurucuda saklanmıĢtır.
3.3. Laboratuvar Analiz Yöntemleri
B noksan ve B toksik koĢullarda, amaçta belirtilen hedeflere ulaĢabilmek için hasat edilen havuç genotiplerine ait örneklerde aĢağıda belirtilen çalıĢmalar yapılmıĢtır.
3.3.1. Bitki örneklerinde fizyolojik analizler
3.3.1.1. Büyüme parametreleri
Belirtilen örnekleme dönemlerinde bitki numuneleri alınarak kökleri ve gövdeleri birbirinden ayrılmıĢtır. Kök ve gövdenin uzunlukları ile yaprak sayısı ve yaprak alanı ölçülerek, yaĢ ve kuru ağırlıkları tartılmıĢtır. Örnekler 70 o
C de 72 saat etüvde kurutulduktan sonra kuru ağırlıkları belirlenmiĢtir.
3.3.1.2. Bağıl su içeriği
Örnekleme dönemlerinde, her bir gruptaki bitkilerin en genç sürgünlerinden sonra gelen lateral yaprakların uç kısımlarından seçilen 6 adet yaprak örneği alınarak (6 ayrı bitki örneğinden) yaĢ ağırlıkları ölçülmüĢtür. 6 saat boyunca dI-H20 içinde petri kaplarında bekletilerek turgor haline gelmeleri sağlanmıĢtır. 70 oC’ de 72 saat etüvde kurutulduktan sonra kuru ağırlıkları alınmıĢtır. Her bir gruba ait yaprak örneklerinin bağıl su içeriği aĢağıdaki formüle göre % olarak hesaplanmıĢtır;
Bağıl Su Ġçeriği (%) = [(YA – KA) / (TA – KA)]x100 YA=YaĢ Ağırlık
KA=Kuru Ağırlık TA=Turgorlu Ağırlık
3.3.2. Biyokimyasal parametrelerin ölçümü
ÇalıĢmanın amaçladığı hedeflere ulaĢabilmesi için aĢağıda belirtilen analizler yapılmıĢtır.
Bitki yaprak örneklerinin B konsantrasyon düzeylerinin belirlenmesi
Prolin içeriğinin belirlenmesi
Lipid peroksidasyonunun belirlenmesi
Hücre zarı geçirgenliği (Elektrolit sızıntısı) tayini
Reaktif oksijen türlerinin (ROT) analizi
o Hidrojen peroksit (H2O2) süpürülme aktivitesinin belirlenmesi o Hidroksil radikali (OH•) süpürülme aktivitesinin belirlenmesi
Total protein miktarının belirlenmesi
Enzim ekstraktlarının hazırlanması,
Antioksidan enzim aktivite analizleri
o Süperoksit dismutaz (SOD) enzim aktivitesinin belirlenmesi o Peroksidaz (POX) enzim aktivitesinin belirlenmesi
o Katalaz (CAT) enzim aktivitesinin belirlenmesi
o Glutatyon redüktaz (GR) enzim aktivitesinin belirlenmesi o Askorbat peroksidaz (APX) enzim aktivitesinin belirlenmesi
Glutatyon (GSH) miktarının belirlenmesi
3.3.2.1. Bitki yaprak örneklerinin B konsantrasyon düzeylerinin belirlenmesi
Örnekleme sonrası bitkilerin vejetatif aksamı ve kök aksamı tamamen temizleninceye kadar musluk suyu ile yıkandıktan sonra sırasıyla bir kez saf su, 0.2 N HCl çözeltisi, iki kez saf su ve bir kez de deiyonize su ile yıkanıp, kaba filtre kağıdı üzerinde fazla suları alınmıĢtır. Daha sonra kese kâğıtlarına ayrı ayrı konulan bitki kısımları hava sirkülasyonlu kurutma dolabında 70 oC’ de sabit ağırlığa gelinceye kadar kurutulmuĢtur. Kuruyan bitki örneklerinin ağırlıkları belirlendikten sonra tungsten kaplı bitki öğütme değirmeninde öğütme iĢlemi gerçekleĢtirilmiĢtir. Polietilen kavanozlara konulan öğütülmüĢ bitki örnekleri analizde kullanılmadan önce 70 oC’de sabit ağırlığa gelinceye kadar kurutma dolabında bırakılmıĢtır.
Kurutulan ve öğütülen bitki örneklerinden 0.2 g tartılarak 5 ml HNO3 ile yüksek sıcaklık (210 oC) ve yüksek basınç (200 PSI) altında mikrodalga cihazında (CEM Mars 5) çözündürülme iĢlemi gerçekleĢtirilmiĢtir. Daha sonra örnekler 25 ml’ lik balonjojeye aktarılarak soğutulmuĢ, deiyonize su ile derecesine tamamlanmıĢtır. Bu süzükler hemen ince gözenekli filtre kağıdı ile süzülerek 25 ml’lik polietilen ĢiĢelere aktarılmıĢ ve süzükte B içerikleri ICP-AES (Inductively Coupled Plasma Atomic Emission Spectrometer) cihazı ile fakültemiz bünyesinde bulunan Elementel Analiz Laboratuvarında yapılmıĢtır.
3.3.2.2. Prolin seviyesinin belirlenmesi
Serbest prolin seviyesinin belirlenmesi Bates ve ark. (1973)’na göre yapılmıĢtır. Sıvı fazdan aspire edilen toluen fraksiyonunun 520 nm’deki absorbansı spektrofotometrede okunmuĢtur. Prolin seviyesinin, kalibrasyon eğrisi kullanılarak hesaplanmıĢ ve µmol prolin g-1
taze ağırlık olarak ifade edilmiĢtir.
3.3.2.3. Lipid peroksidasyonun belirlenmesi
Lipit peroksidasyonunun belirlenmesi TBARS reaksiyonu sonucu oluĢan malondialdehit (MDA) miktarının belirlendiği, Madhava Rao ve Sresty (2000) tanımladıkları yönteme göre yapılmıĢtır. Aktivite için 532-600 nm aralığında absorbans değiĢimlerine bakılmıĢtır.
3.3.2.4. Hücre zarı geçirgenliği (Elektrolit Sızıntısı)
Hücre zarı geçirgenliği elektrolit sızıntısının ölçümüyle Dionisio-Sese ve Tobita (1998)’nın tanımladıkları yönteme göre belirlenmiĢtir. Bunun için 100 mg yaprak örnekleri 5 mm uzunluğunda kesilerek 10 ml deiyonize su içeren deney tüplerine transfer edilmiĢtir. Tüpler plastik kapaklarla kapatıldıktan sonra 32oC’lik su banyosunda 2 saat tutulmuĢtur. Ortamın elektrik iletkenliği EC metre ile ölçülerek (EC1) örnekler 121oC’de 20 dk boyunca tüm dokuların ölmesi ve elektrolitlerin dıĢa çıkması için otoklava bırakılmıĢtır. Sonra örnekler 25oC’ye kadar soğutularak bu ortamdaki elektrik iletkenlikleri ölçülmüĢtür (EC2). Elektrolit sızıntısı (ES) aĢağıdaki formüle göre hesaplanmıĢtır.
ES = EC1/EC2×100
3.3.2.5. Reaktif oksijen türlerinin (ROT) analizi
3.3.2.5.1. Hidrojen peroksit (H2O2) içeriğinin ölçülmesi
Hidrojen peroksit ölçümü için kullanılacak olan örnekler, hasattan hemen sonra sıvı azotta dondurularak depolanmıĢtır ve H2O2 miktarı Liu ve ark. (2010)’nın tanımladığı metoda göre ölçülmüĢtür.
3.3.2.5.2. Hidroksil radikal (OH•) süpürülme aktivitesinin belirlenmesi
Bütün solüsyonlar analiz boyunca taze olarak hazırlanmıĢtır. Deoksiriboz ve OH•’in, Fe3+⁄askorbat ⁄EDTA⁄H2O2 sisteminden ürettiği örnek arasındaki rekabet OH• süpürülme aktivitesinin belirlenmesi için ölçülmüĢtür. Reaksiyon karıĢımı 0.30 ml 20 mM sodyum fosfat tamponu (pH 7.0), 0.15 ml 10 mM 2-deoksiriboz, 0.15 ml 10 mM FeSO4, 0.15 ml 10 mM EDTA, 0.15 ml 10 mM H2O2, 0.525 ml H2O ve 0.075 ml örnek içermektedir. KarıĢım 37°C’de 2 saat boyunca inkübe edilmiĢtir. Ardından 50 mM NaOH içerisinde hazırlanan 750 µl %2.8 triklor asetik asit ve 0.75 ml %1.0 TBA, test tüplerine eklenerek 20 dakika boyunca tüpler kaynatılmıĢtır. Sonrasında karıĢım soğutularak 520 nm absorbansta ölçüm yapılmıĢtır.
3.3.2.6. Total protein miktarının belirlenmesi
Total protein miktarı analizleri, Bradford (1976)'a göre BSA (Bovine Serum Albumine) standartları kullanılarak yapılmıĢtır. Örnekler köre karĢı Shimadzu UV spektrofotometre cihazı ile 595 nm’de okunmuĢtur. Çözünebilen total protein miktarı, mg yaĢ ağırlık olarak belirlenmiĢtir.
3.3.2.7 Antioksidan enzim aktivite analizleri
Enzim ekstraktlarının hazırlanması :
Antioksidan enzimlerin ekstraksiyonu için derin dondurucuda saklanmıĢ olan 1’er g yaprak örneği, soğutulmuĢ havanda sıvı azotta %2 w/v polivinilpolipirolidon (PVPP) ve 1 mM EDTA içeren pH 7.8'de 50 mM Na-fosfat tamponuyla homojenize edilmiĢtir. Ardından +4°C’de, 14 000 rpm’de 30 dakika santrifüj edilmiĢtir. Elde edilen süpernatant, enzim aktivitesi analizlerinde kullanılmıĢtır. Ekstraksiyon prosedürünün tümü ±4°C’de gerçekleĢtirilmiĢtir.
3.3.2.7.1. Süperoksit dismutaz (SOD) enzim aktivitesinin belirlenmesi
Süperoksit dismutaz (SOD) enzim aktivitesi, Beauchamp ve Fridovich (1971) tarafından tanımlanan metoda göre yapılmıĢtır. SOD aktivitesi, 560 nm’de spektrofotometrede ölçülen elektron alıcısı olan NBT’nin fotokimyasal indirgenmesinin %50 inhibisyonuna neden olan enzim miktarı olarak belirlenmiĢtir. Spesifik enzim aktivitesi U/mg protein olarak belirlenmiĢtir. 1 ünite SOD aktivitesi, 25°C’de 1 dakikada 1 µmol substratı ürüne dönüĢtüren enzim (SOD) miktarını göstermiĢtir.
3.3.2.7.2. Peroksidaz (POX) enzim aktivitesinin belirlenmesi
Peroksidaz (POX) enzim aktivitesi, Herzog ve Fahimi (1973) tanımladığı metoda göre yapılmıĢtır. Köre karĢı 465 nm’de H2O2 varlığında okside olan DAB (3'-3'-diaminobenzidin tetrahidroklorit) oluĢum miktarına bağlı olarak 3 dakika boyunca absorbans değiĢimleri okunmuĢtur. Spesifik enzim aktivitesi, dakikada tüketilen µmol/ml H2O2 olarak ifade edilmiĢtir.
3.3.2.7.3. Katalaz (CAT) enzim aktivitesinin belirlenmesi
Katalaz (CAT) enzim aktivitesi, Bergmeyer (1970) tanımladığı metoda göre yapılmıĢtır. Analiz UV ıĢığı bölgesinde köre karĢı 240 nm’de H2O2’nin azalma oranının belirlenmesi ile yapılmıĢtır. Bu enzim aktivitesi dakikada tüketilen µmol H2O2 olarak
ifade edilmiĢtir. Reaksiyon boyunca absorbansta oluĢan düĢüĢ 180 sn boyunca takip edilmiĢtir. CAT aktivitesi dakikada harcanan µmol H2O2 olarak ifade edilmiĢtir.
3.3.2.7.4. Glutatyon redüktaz (GR) enzim aktivitesinin belirlenmesi
Glutatyon redüktaz (GR) aktivitesi, 340 nm’deki absorbans azalmasından yola çıkılarak hesaplanmıĢtır (Foyer ve Halliwell, 1976). NADPH varlığında, okside glutasyon miktarındaki azalma, kuvartz küvette köre karĢı 180 sn boyunca takip edilmiĢtir. Hesaplamalar glutatyon redüktaz enziminin ekstinksiyon katsayısı kullanılarak yapılmıĢtır. Spesifik enzim aktivitesi, dakikada indirgenen 1 mmol ml-1 GSSG miktarı olarak ifade edilmiĢtir.
3.3.2.7.5. Askorbat peroksidaz (APX) enziminin aktivitesinin belirlenmesi
Askorbat peroksidaz (APX) aktivitesinin tayini Nakano ve Asada (1981) göre yapılmıĢtır. Askorbat okside oldukça, spektrofotometreden 290 nm’deki absorbansta oluĢan azalma okunmuĢ ve hesaplamalar askorbatın ekstinsiyon katsayısı kullanılarak yapılmıĢtır. Askorbatın oksidasyonu, enzim ekstraktının katılmasıyla baĢlatılarak, absorbanstaki azalma 180 sn boyunca takip edilmiĢtir. Bir birim APX aktivitesi dakikada okside olan 1 mmol ml-1 askorbat olarak ifade edilmiĢtir.
3.3.2.8. Glutatyon (GSH) miktarının belirlenmesi
Toplam glutatyon (redükte GSH ve okside GSSG) Hawrylak ve Szymanska (2004)’nın metoduna göre sıvı azotta dondurulmuĢ yaprak örneklerinden ekstrakte edilmiĢtir. Elde edilen örneklerin absorbans değĢimlerini 412 nm’de spektrofotometrik analiz sonucunda elde edilmiĢtir.
4. ARAġTIRMA SONUÇLARI VE TARTIġMA
AraĢtırmada, ön çalıĢmalar sonucunda bor stresine karĢı tepkileri belirlenen 4 farklı genotip (6, 9, 12 ve 13) kullanılmıĢtır. AraĢtırmaya ait bulgular alt baĢlıklar halinde verilmiĢtir.
4.1. Büyüme Parametreleri Bulguları
B noksan ve B toksik koĢullarda havuç genotiplerinin temel büyüme parametre (gövde-kök yaĢ ve kuru ağırlık değerleri ile gövde-kök uzunluğu) değerlerine ait varyans analiz sonuçları Çizelge 4.1’de, çalıĢmadan elde edilen ortalama büyüme parametreleri Çizelge 4.2, 4.3, 4.4, 4.5, 4.6 ve 4.7’de, bu değerlere ait grafikler ġekil 4.1, 4.2, 4.3, 4.4, 4.5 ve 4.6’da verilmiĢtir.
Çizelge 4.1. B noksan ve B toksik koĢullarda yetiĢtirilen havuç (6, 9, 12, 13)
genotiplerinin temel büyüme parametrelerine ait varyans analiz sonuçları
Uygulamalar Kareler Ortalaması
GYA GKA G Boy KYA KKA K Boy
Genotip (A) 32,28** 1,26** 74,35** 11,29** 0,15** 22,48**
Bor (B) 27,51** 1,27** 155,93** 21,11** 0,16** 221,82**
A x B Ġnt. 7,87** 0,17** 22,89** 3,47** 0,01** 13,46**
Hata 1,20 0,02 2,26 0,68 0,01 2,57
C.V. (%) 12,25 9,73 5,34 13,58 3,26 5,46
4.1.1. Gövde ve kök yaĢ ağırlığı
Farklı dozlarda B uygulamasının 14. günde, havuç genotiplerinin gövde ve kök yaĢ ağırlıklarına ait ölçülen ortalama değerler Çizelge 4.2 ve 4.3’de verilmiĢtir. Varyans analiz sonuçlarına göre, gövde ve kök yaĢ ağırlığı değerleri, B noksan ve B toksik koĢullarda 14. günlerde alınan örneklerde %1 seviyesinde önemli bulunmuĢtur (Çizelge 4.1).
Havuç genotiplerinin ortalama değerlerine bakıldığında uygulamalara bağlı olarak genotipler içerisinde en yüksek gövde yaĢ ağırlığının 12. genotipte olduğu tespit edilmiĢtir. Farklı B dozu uygulamaları ele alındığında ise bitki için yeterli düzeyde olan B (C) uygulaması sonucunda, gövde yaĢ ağırlık değerlerinin en yüksek seviyeye ulaĢtığı tespit edilmiĢtir.
AraĢtırmada kullanılan havuç genotiplerinin gövde yaĢ ağırlık değerleri incelendiğinde B noksan ve B toksik koĢullarda bitkilerde farklılıklar meydana geldiği belirlenmiĢtir. B noksan koĢullarda genotiplerde azalmalar meydana gelmiĢtir. Bu azalma oranlarının birbirine yakın olduğu ve en fazla azalmanın %39 oranında 12. genotipte olduğu tespit edilmiĢtir. Artan B dozlarıyla (0.5 ve 1 mM B) birlikte oluĢan toksisite koĢullarında, 9, 12 ve 13 numaralı genotiplerde uygulamalara bağlı olarak gövde yaĢ ağırlık değerlerinde azalmalar görüldüğü en fazla azalmanın her iki B dozunda da 9. genotipte sırasıyla %37 ve %34 oranlarında olduğu belirlenmiĢtir. Diğer genotiplerin (9, 12, 13) aksine 6. genotipte, 0.5 ve 1 mM B uygulamarında sırasıyla %28 ve %17 oranında gövde yaĢ ağırlık değerlerinde artıĢlar olduğu tespit edilmiĢtir. Toksisite oluĢturmak amacıyla uygulanan yüksek B (2 mM B) dozunda ise tüm genotiplerde azalmalar meydana geldiği en fazla azalmanın 9. Genotipte %53 oranında olduğu, 6. genotipte ise %5 oranında azalma ile diğer genotiplere oranla toksik B koĢullarında gövde yaĢ ağırlığının daha az etkilendiği tespit edilmiĢtir (Çizelge 4.2, ġekil 4.1).








![ġekil 4.9. 6. genotipte; kontrol (C) ve B uygulamalarına ait görseller [a (C x 0 mM B), b (C x 0.5 mM B), c (C x 1 mM B), d (C x 2 mM B)]](https://thumb-eu.123doks.com/thumbv2/9libnet/4950020.99760/51.892.193.747.130.272/ġekil-genotipte-kontrol-uygulamalarına-ait-görseller-mm-b.webp)
