Biofortification and Localization of Zinc in Wheat Grain
Tam metin
Şekil
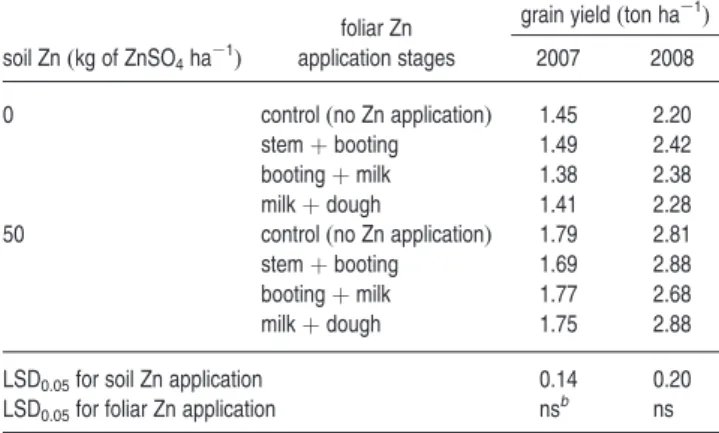
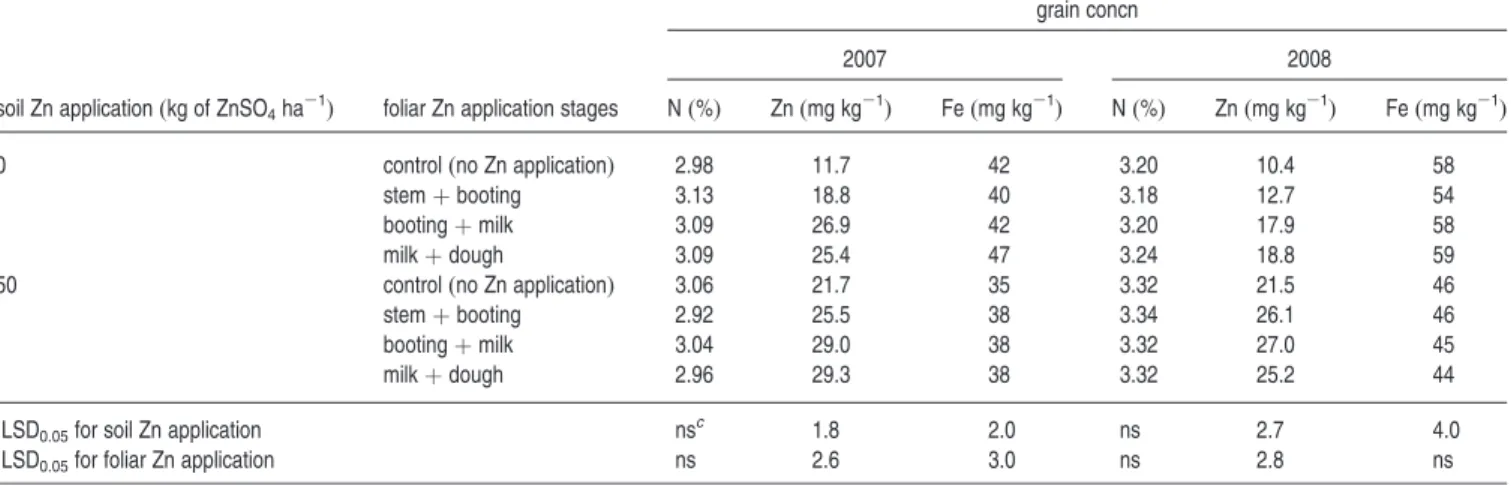
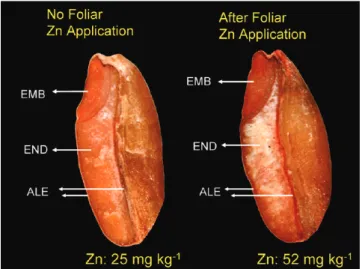
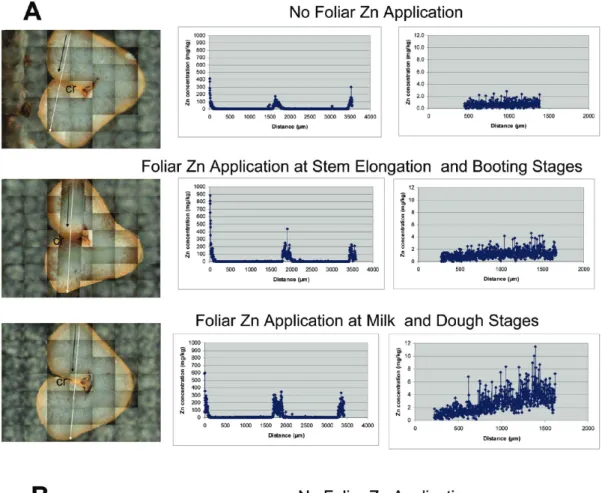
Benzer Belgeler
Kanlıca’nm BizanslIlar zamanında da mevcut olduğu bulunan eserlerden anlaşılmaktadır.. Bizansm zenginleri Kanlıea’da
The activation of epidermal growth factor receptor gene (EGFR), located on chromosome 7, the homozygous deletion of the p16, located on the short arm of chromosome 9, deforming of
To assess daily intake of metal, consumption of seafood gets more quantity of metal than other kinds of food and Zn is the highest.. To assess THQ and the risk of
The Prevalence of Violence and Violent Characteristics in Alcohol and Drug Abusers Compared with Controls Objective : To identify the prevalence of violence in alco- hol and
https://yazilidayim.net/ ZÜBEYDE HANIM MESLEKİ VE TEKNİ ANADOLU LİSESİ DİN KÜLTÜRÜ VE AHLAK BİLGİSİ 9 .SINIF 1.DÖNEM 1.YAZILI
[r]
“Kapasiteler ve Kapsayıcılık” başlığı altında ise “kapasite” kavramının, en az demokrasi ve eşitlik kadar kalkınma olgusu için önemli olduğu
Sonuç olarak, düşük doğum ağırlığı, erken doğum ve postnatal yoğun bakım tedavisi alma, tarama ABR sonuçları ile en çok ilişkili risk faktörleri olup, yenidoğan
