Başlık: Molecular characterization of some selected wild olive (Olea oleaster L.) ecotypes grown in TurkeyYazar(lar):OZKAYA, Mücahit Taha; ERGÜLEN, Erkan;ÜLGER, Salih;ÖZELBEY, Nejat Cilt: 15 Sayı: 1 Sayfa: 014-019 DOI: 10.1501/Tarimbil_0000001066 Yayın Ta
Tam metin
Şekil

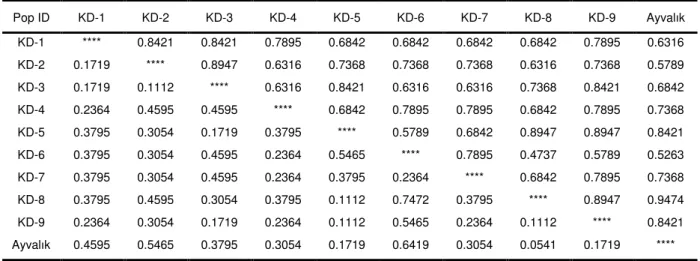

Benzer Belgeler
287] from a calculation dealing with coherent sheaves to the relatively better understood domain of vector bundles under suitable conditions, namely when the
• Bir problemle baş etmeye çalışırlar. Problem çözücü konumundadırlar. • Araştırma ve problem çözme sürecine katılırlar. • Arkadaşlarıyla ve
Başka bir deyişle, öz yeterlilik algısı yüksek olan işgörenlerin lider-üye etkileşimi açısından iç grupta yer alacakları, dolayısıyla daha az tükenmişlik
Şeftali bahçesi tesisi için yapılacak yatırımın karlı olup olmadığını görmek amacıyla yapılan fizibilite etüdü sonucunda, 20 dekarlık bir şeftali
For the case that had various occupational ex- posures and cigarette anamnesis, local air trap- ping secondary to bronchial obstruction was suspected and a contrast-enhanced computed
Tablo1’de yer alan araştırmaların sonuçlarına göre, tüketicilerin izinli mobil reklam mesajlarını kabul etme, izinsiz olarak gönderilen mobil reklamları ise kabul
Zorunlu dersler kapsamında ilk dönem elle teknik çizim yapmayı öğrenen öğrenci, ikinci dönem BDT I dersinde Autocad, BDT II dersinde Sketch Up, Mimari Uygulama I
The decay resistance of untreated and heat-treated Scotch pine (Pinus sylvestris), oak (Quercus petreae) and beech (Fagus orientalis) wood specimens against white rot
