Çeşitli kaynaklardan bakteriyal selüloz üreten bakterilerin izolasyonu ve karakterizasyonu
Tam metin
Şekil



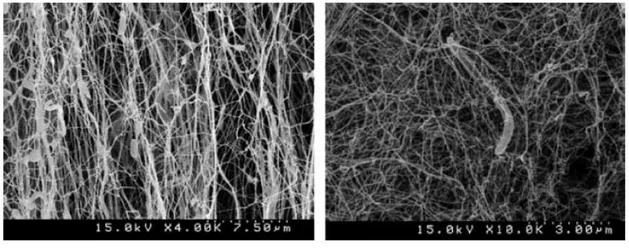




Benzer Belgeler
Türkler buraya «Beş | taş» admı vermişler, sonra uykuya yattığımız için olacak taşı bile I ninni söylemek için beşik yapmışız.. Ortaköy o zamanlar
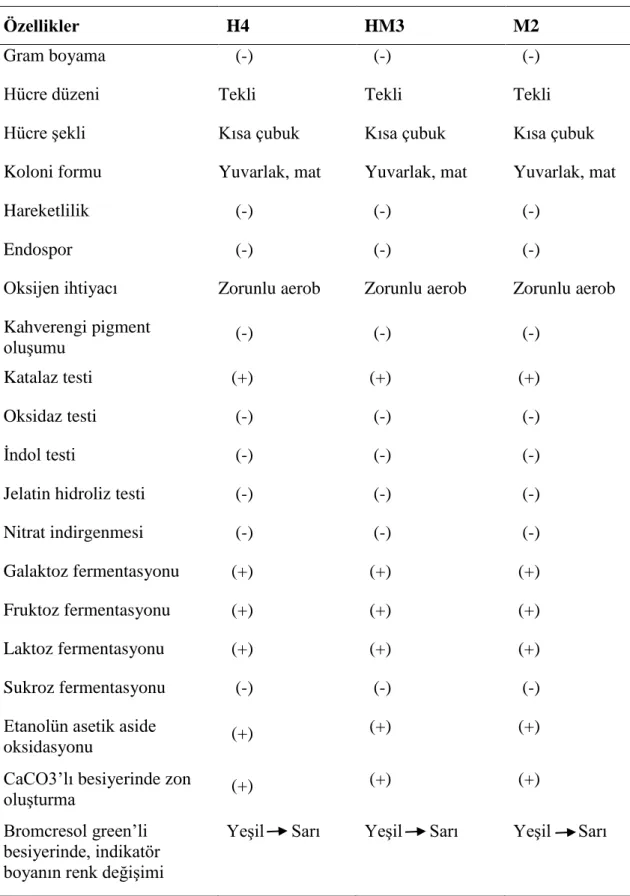
Uygun enzim üretim besi yerine ekilmiĢ olan izolatlar daha önceden belirlenmiĢ olan uygun pH ve uygun sıcaklığa ayarlanmıĢ etüve konularak inkübasyona bırakılır ve 6, 12,
Eğitim ve teknoloji ilişkisi; temel kavramlar; öğretim teknolojisinin tarihsel gelişimi; Türkiye’de 1980 ve sonrası öğretim teknolojilerinin gelişimi
Yıldız Zamanı ( = YZ) : Herhangi bir yıldızın S saat açısı ile
Bu düşük hata oranı, aminoasil-adenilat formasyonundan sonra da izolösil tRNA sentetaz enziminin iki amino asidi ayırdığına işaret etmektedir..
Sonuç olarak HBB-247 izolatının ürettiği bakteriyosin amonyum sülfat çöktürmesi ile kısmi olarak saflaştırılmıştır ve anyon değiştirici kromotografiden
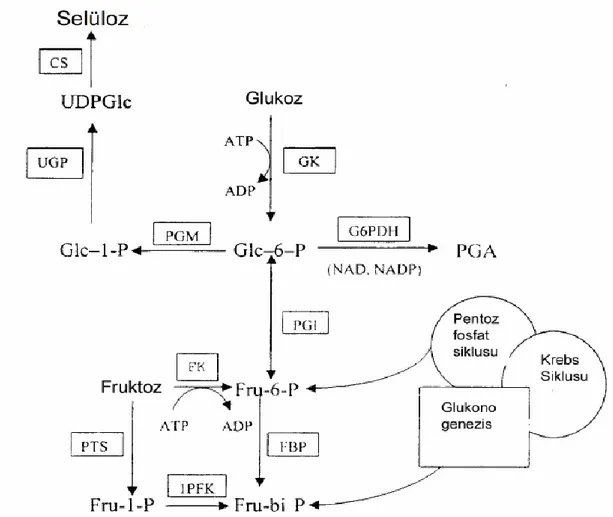
► Selüloz molekülleri hücre çeperinde düz Selüloz molekülleri hücre çeperinde düz.. paralel
Örn: turşu, sirke, süt ürünleri asit üreten bakterilerin; bira, şarap gibi alkollü içkiler ise alkol üreten mayaların fermantasyon ürünleridir.. İnsanoğlu ilk
