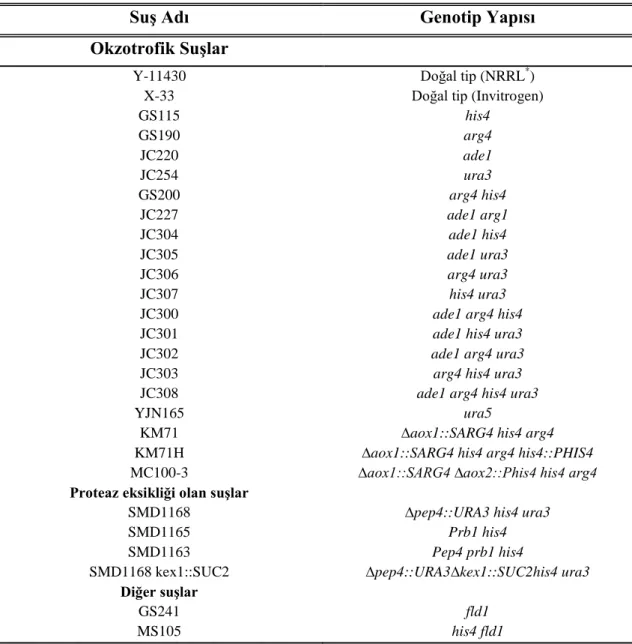
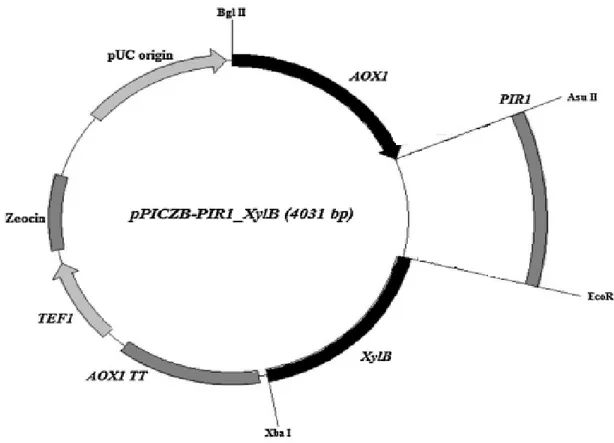
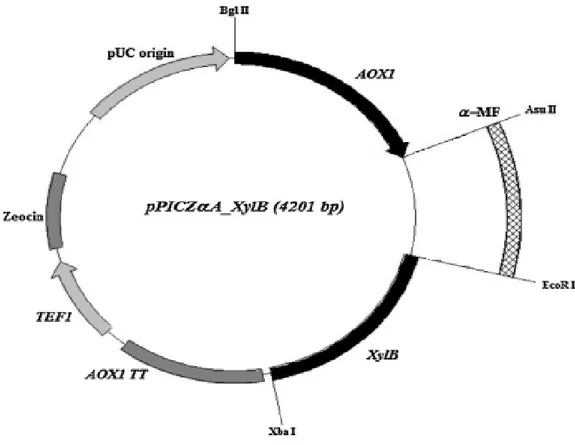
Pichia pastoris mayası ile rekombinant ksilanaz enzim üretiminin optimizasyonu
Tam metin
Şekil


Benzer Belgeler
Gebelerde s›k görülen postüral de¤ifliklikler ileri bafl, yuvar- lak omuz, bafl ve boynun toraksa göre anterior pozisyonu, dor- sal kifozda artma, lomber lordozda artma,
[r]
A descriptive survey study was conducted between January 2006 and December 2011 among 44 patients (18 women, 26 men) with a permanent stoma receiving care at the Kartal Training
Risk factors for acute kidney injury in critically ill patients receiving high intravenous doses of colistinmethanesulfonate and/or other nephrotoxic antibiotics: a
Yazımızda, gebelik döneminde düşük molekül ağırlıklı heparin kullanımı öyküsü olup laktasyon döneminde osteoporoz saptadığımız hastamızı sunarak, gebelik
Осы орайда Бес уақыт намаз: таң (Бамдат) (таң қылаң бере таң намазын оқитын кез), күннің еңкейген мезгілін бесін (бесін намазын оқитын кез) деп
İlgili kişi, nöbet sırasında işyerinde veya işveren tarafından belirlenmiş olan başka bir yerde bulunmayıp, imkanları ölçüsünde özel alanında serbest
Mozaik yapımında “opus seetile”, “opus vermiculatum”, “opus aleksandri- num” ve “opus tesselatum” deni len teknikler kullanılıştır.. Opus Seetile:
