A.
tJ.
Veteriner Fakültesi Bakteriyoloji ve Salgınlar Kürsüsü Prof. Dr. Mustafa ArdaENTERİK İNFEKSİYONLARDA YEREL BAGışmLIGIN ROLÜ
Ersİn İstanbulluoğlu.
The Role of Loeal Immunity in Enterle Infeetion Summary: During recent years a considerable amount of evidence has bem accumulated that gastro-intestina! system possess a collective immUlıe me-chanism which is composed of immunologic and nonimmunologic means against iı!fections. The predominant immUl:oglobulin type in intestinal secretions is secrotory dimeric IgA(SIgA). Experimental evidence also suggests that there an inCipendent cell-mediated immUlıity in intestinal tract.
Özet: Son yıllarda gastro-intestinal sistemin immunolojik ve immuno-lojik olmayr.n vasıtalarden oluşan kendine özgü bir sisteme sahip olduğu hak-kında yeterli deneysel veriler elde edilmiıtir. Sekrotorik IgA(SIgA) barsak kcnalında en yüksek orande bulunan immunoglobulin türüdür. Aynca, barsak kanalının lokal bir hücresel immunitc.;'e sahip olduğu da ortaya konmuştıır.
Giriş
Gastro-intestinal sistem, yakın bir zamana kadar yalnızca or-ganizmanın yaşam sürecini devam ettirebilmesi için gerekli enerji ve doku yenilemelerinde kullanılan temel maddelerin, alınan besin-lerden üretildiği bir organlar grubu olarak bilinmektc idi. Fakat, son yıllarda yapılan çalışmalarla bu sistemin yukarıda açıklanan a-na görevi yanı sıra, canlının yaşamını tehlikeye sokan çeşitli etken-lere karşı korunmasında da önemli roloynadığı ortaya konulmuştur (13, 23).
Gastro-intestinal sistem, alınan gıdalar nedeni ile devamlı çe-~itli toksik, allerjik ve infeksiyöz etkenlerle kar~ı kar~ıya gelmekte ve bu zararlı ajanlarla vücut dokuları arasında ilk koruyucu tabaka-yı olu~turmaktadır. Dı~arıdan sindirim yoluyla giren ve geçici bir süre bu sistemde barındıktan sonra dı~arı atılabilen etkenlerin yanı sıra çe~itli cins ve sayıda zararsız mikroorganizma gastro-intestinal bo~luklarda sürekli olarak bulunmakta, ya kendileri ya da salgıla-dıkları çe~itli metabolizma ürünleri antijenik karaktere sahip bulun-maktadır.
Gastro-intestinal sistem sahip bulunduğu immunolojik ve im-immunolojik olmayan vasıtalarla gerek kendisini gerekse canlının dokularını zararlı etkenlerden korumaktadır. Immunolojik olmayan koruma vasıtaları tek tek zayıf bir koruma yeteneğine sahip iseler de bir bütün olarak, bilhassa barsaklardaki mukozal savunma sis-teminde, etkin bir yardımcı güçtürler (I4, 15)'
Çe~itli zararlı etkenlere kaqı korunmayı sağlayan immunolo-jik ve immunoIoimmunolo-jik olmayan mekanizmalar tablo- ı de toplu .9larak
gösterilmi~tir .
Tablo ı. Gastro-intestİnal sistemde bağışıklığı sağlayan mekanizmalar Immunolojik olanlar Immunolojik olmayanlar Hümoral bağışıklık
(IgG, IgM, IgA) Hücresel bağışıklık
Müköz tabaka Gastrik engel
Lizozim ve safra tuzları Perista!tik barsak hareketi Doğal barsak florası Hepatik engel
Gastro-intestinal sistemin epitel dokusunu kaplayan muköz ta-bakanın yapısında bulunan musinIer (glikoprotein ve glikolipidler) mikroorganizmaları veya bunların toksik ürünlerini tutma özelli-ğine sahip algaçlara (receptor) benzeyen moleküler yapıdadırlar. Böylece, birçok mikroorganizmanın veya organizma için zararlı olan maddelerin epitel tabakaya ula~ması engellenmi~ olmaktadır
(16, 18).
Asidik özelliği dolayısı ile mide sıvısı, birçok mikroorganizmayı veya bunların toksinlerini barsak kanalına ula~madan zararsız ha-le getirir. Goldstein ve ark. (ıo) yaptıkları çalı~malarla achlorhydric bireyIerde barsak infeksiyonlarının daha sık görüldüğünü ortaya koymu~lardır. Case ve ark. (2) 2gr. bikarbonat verildikten sonra
ko-26 Ersin İstanbulluoğlu
lera mikrobu (108) ile infekte edilen ~ahıslarda sürgünün yalnız
mik-roorganizma verilenlere oranla 8 misli fazla olu~tuğunu bildirmek-tedirler. Aynı sonuçlar enteropatojeni'k E. coli ile yapılan deneyiJi1~ lerde d~ elde edilmi~tir (6).
Lizozimler ile safra tuzlarınT.n bakterilerin çoğalmalarını önleyici etkisi in vitro çalı~malarda ortaya konmuş ise de konakçının barsak kanalındaki mikroorganizmalara karşı in vivo oynadıkları rol kesin olarak bel irlenememiştir (24)'
Barsak lumeninde doğalolarak, bulunan mikroorganizmaların türleri ve sayıları konakçının hastalık etkenlerinden korunmasına yardımcı olan bir husustur. Germ-free (Normal mikropsuz) fare-ler doğal çevreye çıkarıldıkları zaman çoğu kez kolayca şiddetli gast-roenteritis ve s:stemik infeksiyonlara yakalanırlar (5). Normal bar-sak florasında bulunan zararsız mikroorganizmalar patojenik o-lanlarla mevcut gıda için rekabet ederek sonuncuların konakçıya zarar verecek miktarda üremesine engelolmaktadırlar. Ayrıca, sap-rofitik bakterilcr patojenik olanlar için bakteris:dal veya bak't~rios-tatik olan birçok maddeleri (koli~in gibi) sentezleyebilmektedirler
(8).
Enterotoksinlei- bazen ince barsak engelini a~arak portal do-la~ı-ma ula~abilirler. Walker ve Isselbacher -(20)'e göre bu durumlarda infeksiyonun meydana gelmemesi karaciğerin böyle zararlı madde-leri dola~ımdan süzerek ayırma yeteneğine sahip olmasındandır.
İMMUNOLOJİK MEKANİzMALAR
Hümoral bağışıklık: Son on yıl içinde yapılan geni~ kapsamlı ça~ lı~malar sonucu bir çok memeli türünde pe-riferal kan dola~ımına veya diğer sistemik bağı~ıklık organlarına (dalak, timus gibi) bağım-lı olmayan bir immun sistemin varlığı ortaya konulmu~tur (7, 23)' Çeşitli deneysel verilerle desteklenenbir hipoteze göre, peyer plfı.k-ları veya diğer intcstinal lenfoid doku hücreleri lokal bir antijen ta-rafından uyarıldıkları zaman blastoid karakterde hücreler haline dö-nü~üp, bölgesel lenf yumrularına göç ederek buralarda olgunla~-makta ve olgunla~an hücreler kan yoluyla barsak dokusunun lami-na propria tabakası ile meme dokusuna. ve daha az. miktarlarda da diğer mukozal yüzeylere yerleşerek IgA (Şekil-ı) sentezleinektedir-ler (3, 22). Barsak lenfoid doku hücreleri in vitro üretildikleri
za-man immunolojik olarak olgun haıC gelmezler. Olgunlaşmaları için herhangi bir antijen tarafından uyarılmaları ve T-lenfositIer ile te-masları gereklidir (22). Guy-Grand ve ark. (ı ı) bildirdiğine göre, doğuştan timussuz olan farelerde peyer plaklarındaki germinal mer-kezleri çok az g~lişmiş olup ya hiç veya çok az sayıda lenfositlere sa-hiptirler. Timik doku graftları, T-lenfosit populasyonlarını yeniden oluşturmakla kalmayıp payer plaklarındaki germinal merkezlerin de gelişmesine neden olurlar. Bunun sonucu olarakta gerek serum ve gerekse barsak 19A konsantrasyonlarında önemli oranlarda yük-selmeler meydana gelir.
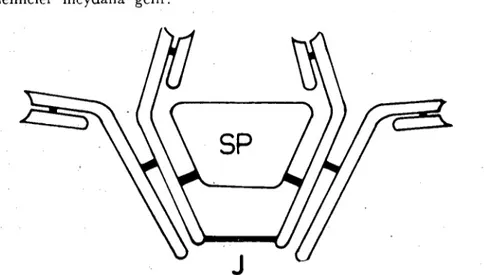
Şekil ı. IgA Yapısı-Aktarım parçası (SP), manamerleri bağlayan polipeptit zinciri (J).
Barsak epiteli yüzeyinde bulunan antijen veya antijenler mik-tarlarına göre, iki farklı şekilde lamina propria tabakasındaki im-mun yetenekli hücrelere ula~maktadırlar. Antijen, az miktarlarda ise !cimina propria'y;:ı morfolojik bakımdan kendine özgü bir yapı-ya sahip M-epitel hücreleri tarafından taşınmaktadır. Eğer antijen çok miktarda ise tüm epitel yüzeyi kaplayan basit bir yayılım (dif-fusion) olayı ile lamina propria'ya ulaşmaktadır (22).
İnce barsak ~ekresYGnunda en yüksek konsantrasyonda bulu-nan immuneglobulin türü dimerik bir yapıya ~ahip olan sekretorik 19A (SlgA) dır. SlgA,
J
halkası diye tanımlanan bir yapı ile birleş-tirilmiş iS lik 19A monomerinden yapılmıştır (Şekil-ı). Ayrıca, sck-retorik komponent (SC) diye bilineb, immunoglobulin karakterinde olmayan bir protein de SIgA'nın yapısında bulunur. SC, ayniza-28 Ersin İstanbulluoğlu
manda, bütün sekretorik hücrelerin içinde veya aralarında yer alır. SC'nin görevi SIgA yı diğer immunoglobulin türlerinin ince barsak-larda maruz kaldıkları enzimatik parçalanmadan korumaktır. Ay-rıca, SC, SIgA'nın ince barsak kriptlerindeki hücrelerden mukozal yüzeye aktarılmasına yardımcı olur (2ı).
SIgA İnce barsaklarda yerel olarak sentezlenir (3). Lamina propria'da bulunan plasma hücrelerinin yüzeylerinde IgA'nm var-lığı histoşimik yöntemlerle ortaya konmuştur (4). Ayrıca, in vitro barsak organ kültürlerine antijenik uyarı yapıldığı zaman IgA'nın sentezlenerek lumen boşluğuna salgılandığı Welliver ve Ogra (23)
tarafından saptanmıştır. Steel ve ark. (I7)'e göre SIgA sentezlerne yeteneğinden mahrum insanlarda ince barsaklarda yüksek miktar-da polimerik IgM türü immunoglobulinler bulunmaktadır.
Fonksiyonel olarak SIgA opsonik, komplemanı tutucu ve bak-terisidal etkilere sahip değildir (17). Yerel ve sistemik olarak sentez-lenen IgG, IgM ve IgE türü immunoglobulinler dokulardan veya kan dolaşımından ince barsak lumenine diffuse olabilirler. Fakat, SIgA'nın aksine bunlar villilerin uç kısmında yer alan epitel hücre-lerini geçerek mukozal yüzeye ulaşırlar ve SC tarafından
korunmaz-lar (22). Bu nedenle de daima hücre içi enzimlerin parçalayıcı
et-kisinde kalırlar. SIgA'nın dışındaki immunoglobulin türleri barsak lumeninde düşük konsantrasyonlarda bulunurlar. Ancak barsak epitelinin yangısal bozukluklarında IgG ve IgM türü immunoglo-bilinlerin konsantrasyonlarında yüksek oranda artışlar meydana ge-lir (12).
Hücresd bağışıklık: İnce barsaklarda yerel bir hücresel bağışıklık mekanizmasının bulunduğuna dair deneysel veriler mevcuttur. T-tipi lenfosiderin fonksiyonel bozukluğu bulunan insanlarda çocuk felci aşısı yapıldığı zaman bunların devamlı olarak dışkıları ile virus çı-kardıkları halde normal bireyIerde aşılamadan kısa bir süre sonra virus atılımının durduğu saptanmıştır (23). Waldaman ve Ganguly (ıg) göre payer plaklannda bulunan öncü i (precursor) hücreler bar-saklardaki hücresel bağışıklık görevini üstlenirler.
Barsaklarda bulunan çeşitli tür antijenlere karşı yerel bir bağı-şıklık sisteminin varolduğunun saptanması, çeşitli hastalıkları (in-feksiyöz ve allerjik) kontrolünde yeni ufukların açılmasına ne-den olmuş; yakın zamana kadar pek başarılı sonuçlar alınamayan enterobakteriyel infeksiyonların' aşılama yöntemiyle kontrolünde ümit verici sonuçlar elde edilmiştir (I, 9).
Literatür
1- Anon. (1972): Ore! enterie baeterial vaeeines. WHO Tech. Rep. Ser. Geneva. 2-44.
2- Case, R. A., Music, S. I., and Libonan, j. P. (1974): Response of man to in/eetion with Vibrio .eholera. i. Clinical, serological and baeteriologie response to a known inoeulum.:
J.
Infeet. Dis. 129: 45-52.3- Cebra; j.
J.
(1973): Secret07Y Immunoglobulin A. The -moleeule and its bios..ynthesis, in Mergenhagen, S. E., aııd Sharp, R.A. (Ed). Compe-mtive immunology of the oral eavity. Philadelphia. DHEW, PubL. No: 73-438: 3-38.4- Crabbe,P. A.,andlleremans, j. F. (1966): The distribution of immunoglobü.lin eoııtaining eells along the human intestinal traet. Gast-roenterology. 51:3 305-316.
5- Dubos, R. j., and Schaedler, R. W. (1960):Effeet of intestinal flora on growth rate in miee and on their suspeetibili(v to experimental
infeeıians.
J.
Exp. Med. i i i: 4°7-417.6- Dupont, H. L., Formal, S. B., and Homick, R. S. (1971): Pathogenesis of E. eoli diarrhea N. Engl. J.M. 285: 1-9
7- Frederick, G. T., and Bohl, E. H. (1976): Loeal and Systemie eell-mediated immunity against transmissible gastroenteritis, an intestinal viral infeeti on of swine.
J.
ImmunoL. 116: 1000-1004.8- Freter, R. (I962): In vivo and vitro antogonism of intestinal bacteria against Shilgella flaxneri i I.Inhibitioıı mechanism.
J.
Infeet. Dis. i i i : 37-48.9- Fubara, E. S., and Freter, R. (1973):Protetion against enterie bae-terial infection by seeretory IgA antibocies.
J.
ImmunoL. i i i: 395-403I,cr Goldstein, F., Wirtis, C. W., andjosephs, L., (1962): Bacte-rial flora of small in tes/ine. Gastroenterology. 42: 755-772. 11- Guy-Grand, D., Griscelli, C., and Vassalli, P. (1975):Payer's
patehes gut IgA plasma eells and thymic funetion: Study in nude miee bearing thymie grafls.
J.
ImmunoL. 115: 361-364.12- Haneberg, B., and Aarksog, D. (1975):Humanfaecal immunog-lobulin in healty infants and in some with disease effeeting the intestinc! tract or the immun s..Ystem.Clin. Exp. Immuno!. 22: 210-222.
30 '. Ersin İstanbulluoğlu
13- Husband, A.
J.,
and Watson, D. L. (1978): Immunity in the intestine. Ye't. BuJI. 48 :91 1-9,10.14.-Laseelles, A. K., and MeDowell, G. H. (1974): Loealized humo-ral immunity with partieul(!r referenee to ruminants. TranspJant Rev.
19: 170-208.
15- Porter, P. (1973): Intestinal defence in the young pig. Vet. Ree. 92: 658-664.
16- Springer, G. F. (197°): Importanee of blooe group substances in interaetions between man ene mierobes. Ann. X.Y. Aead. Sci. 169:
134-141.
17- Steele, E.
J.,
Chaieumpa, W., and Rowley, D. (1974): lsolc-tion cnt. biologiecl properties of three classes of rabit antibor:y to vibrio c!lOlerae.J.
Infeet. Dis. 130: 93-~°3.18- Strombeek, D. R., and Harrold, D. (1974)' Bindingof Choleree toxin to mueins and inhibition by gestrie muein. Infeet.' Immun. 10:
1266-1272.
19- Waldman, R.H., and Gangu:ly,R. (1974) ImmUlıity in infeetions on seeretoıy surfcee.
J.
Infeet. Dis. 130: 93-103.20- Walker, W. A., aıidlsselbaeher,K.
J.
(1974): Uptake and.trans-port of maeromoleeules by the intestine: Possible role in clinical disor-cers. GastroenteroJogy. 61=, 531-55°.21- Walker, W. A. (I 976): Host defence me'chanisms in the gaStrointes-tinal traet. Pediatrres. 57: 9°1-916.
22- Walker, W. A., and Isselbaeher, K.
J.
(1977): Intestindı cnti-bodies. N. Engl.J.
Med. 297: 767-773.23- Welliver, R. C., and Ogra, P. L. (1978): Importanee of loee! immunil)' iıı enteric infeetion. J..A.Y.M.A. 173: 160-'164.
24- Williams, R. C., Showalter, R:, and Kern, F. (1975): In vivo effect of bile salts andcholestyramine on' intestinal anaerobie baeteria; Gastroenterology. 69: 48 5~490.