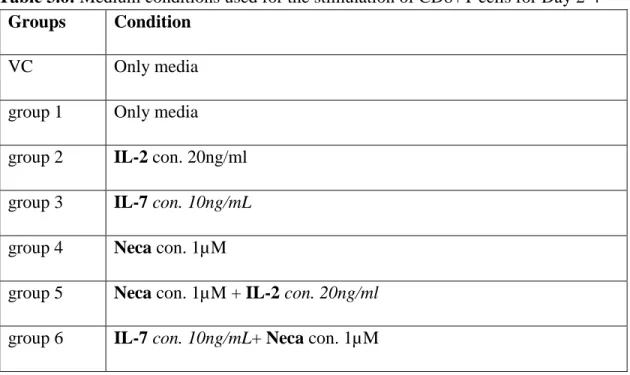
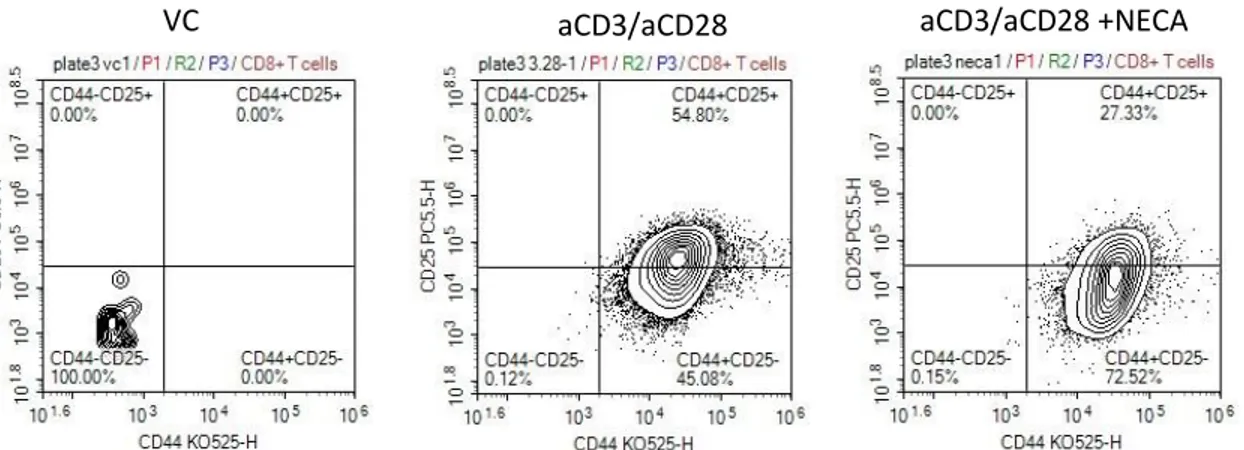
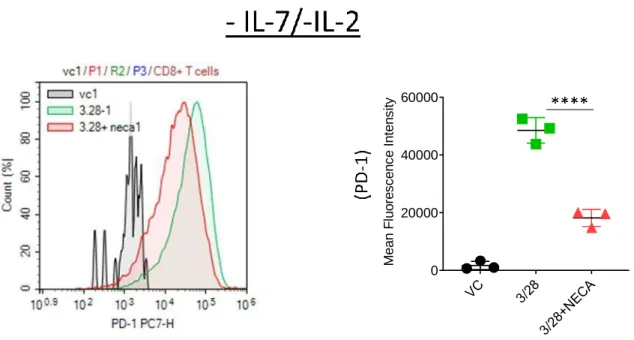
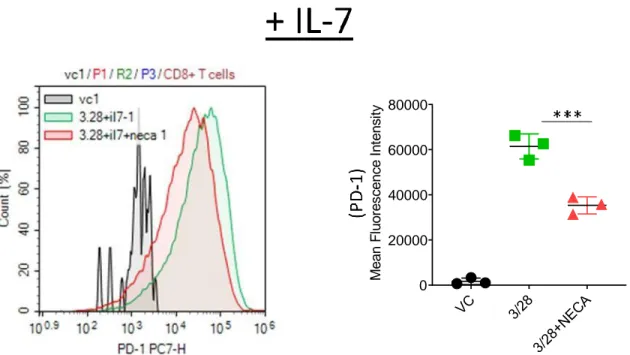
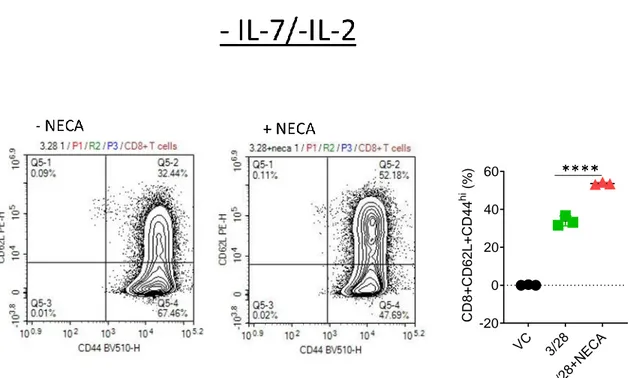
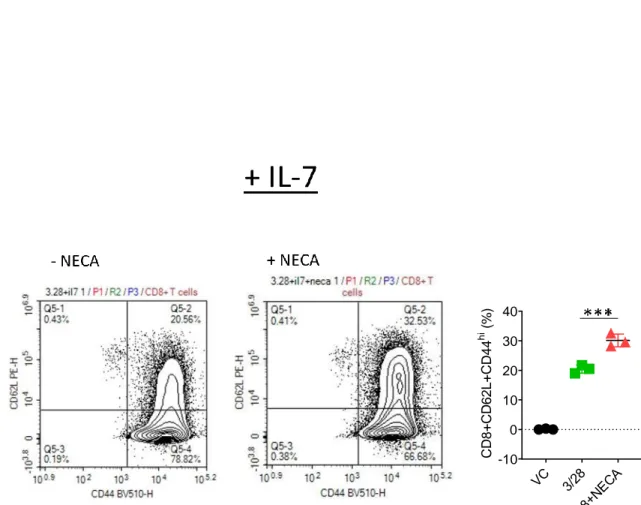
The effect of interleukin 7 on CD8+T cell accumulation and differentiation in the presence of adenosine signaling
Tam metin
Şekil



Benzer Belgeler
Timusta merkezi olarak kontrol noktaları bulunmakla beraber yeterli değildir ve periferde T hücrelerinin selfe karşı aktif efektör hücreler haline gelmesini engelleyen iki
In most cases, the clinical features of EATL are very si- milar to celiac disease in the initial stages, which usual- ly leads to a late diagnosis.. EATL is primarily observed during
Earlier theoretical studies have predicted so far two dif- ferent stable structures of 2D, free-standing SL GaN, namely, planar honeycomb structure with hexagonal lat- tice, 17 , 20
In the decision-level fusion scheme with global (sensor-independent) training model, a common classification model is used for the feature vectors extracted from
2013 年國際口腔雷射應用醫學會(SOLA)世界年會假北醫大盛大舉行,來自歐 美亞等國近 200 名專業人士與會
Ahidnamelere aykırı olarak yapılan bu hareketlerin Osmanlı devletine bildirilmesi sonucu, kanuna göre hareket edilmesini ve Fransa’nın kadim dost olması nedeniyle
BÜYÜK ÂFET, asıl adı Yoıjgiki olan b ü Hıtrvat delikanlısıydı; o zamanlar İstadbuMa Yorjgakisiz düğün olmaz gibiydi; tdk kusuru,R. •
In terms of location; Since the circulation of complicated surgical operations is intense, especially in the 3rd level public university hospitals it is more appropriate
