Kuraklık stresi altındaki mısır (Zea mays L.) fidelerine dışarıdan uygulanan sodyum nitroprussid' in (SNP) kloroplastik antioksidan enzim aktiviteleri üzerine etkilerinin belirlenmesi
Tam metin
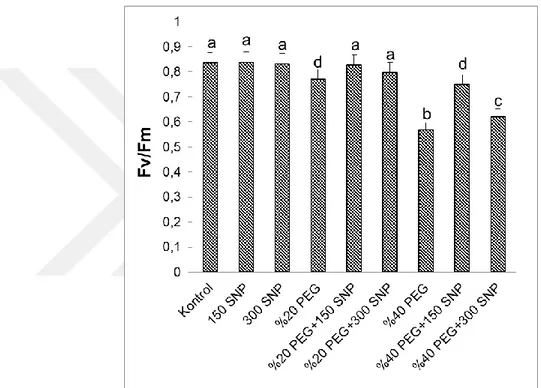
Şekil

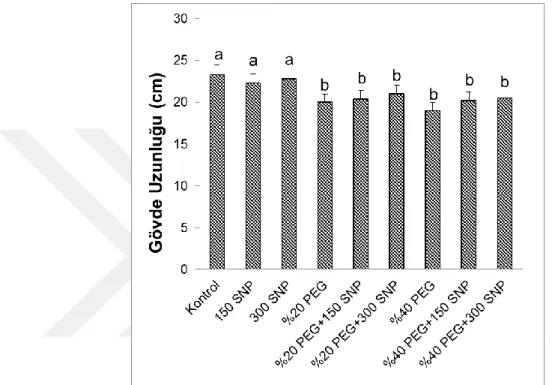
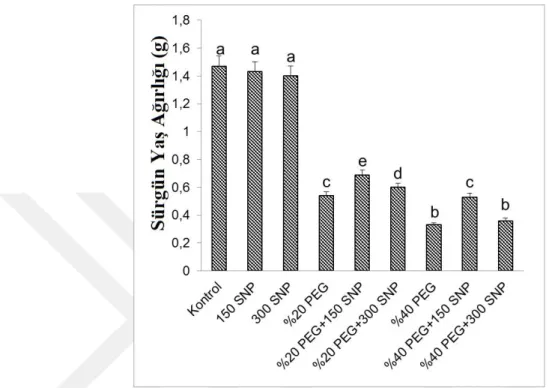
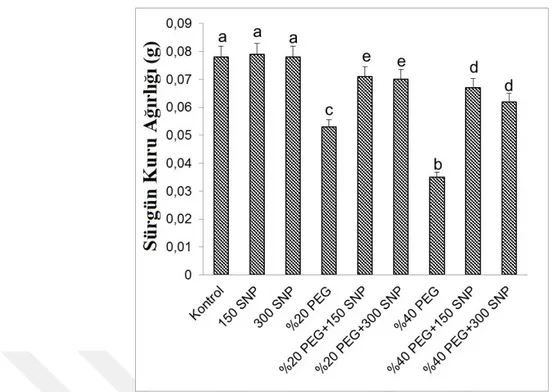
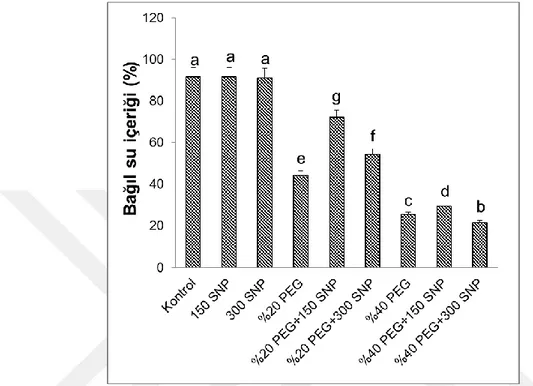
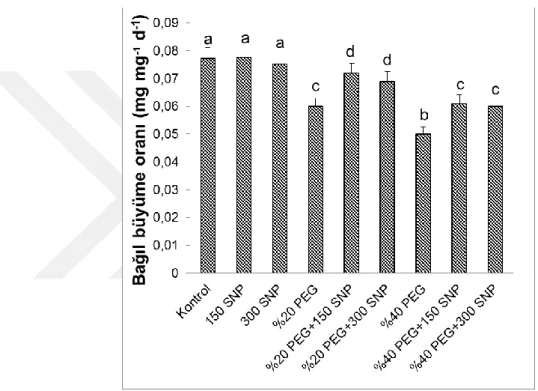
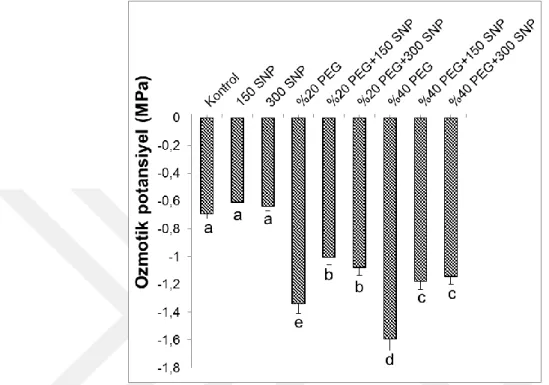
Outline
Benzer Belgeler
Kontakt dermatit cildi tahriş eden veya alerji oluşturan bir maddeyle doğrudan temas edilmesi sonucunda ortaya çıkar.. Sebo- reik egzamada yaralar genellikle yağ bezlerinin
Ancak Nieder ve arkadafllar›n›n, yanal al›n lobundaki nöronlar›n %31’inin, yanal kortekstekilerininse %14’ünün belirli say›lara tepki verdi¤ini öne
Redüksiyonsuz disk deplasman› hastalar›n›n muayenesinde ilk dikkat edilecek husus, açma s›ras›nda çene ucunun rahats›z- l›k olan tarafa do¤ru bariz bir
In this study, as a result of that fi- nancial crisis, evolution process and content of Basel III criteria is going to be handled after proposed and predicted changes for
Boş geçen zamanlarımızı olumlu biçimde değerlendirme, gö nüllü hizmet yapma, çevremizdekileri iyiye, doğruya, güzele yönelt me yolları o kadar çoktur
Sözlerimi yazarın şu çağrısıyla bitirm ek istiyorum : «Yalnız eğitimle uğraşan kişiler olarak değil, tüm toplum olarak, insan yetiştirm enin o büyük
Bu araştırmanın amacı, Spor yönetimi disiplininde yapılmış olan tezlerin araştırma eğilimlerinin kapsamlı bir şekilde analiz edilmesidir. Araştırma betimsel bir
UZ, a.g.e. Đhale sürecindeki konuların itirazen şikayet başvurusu dışında herhangi bir yolla Kamu Đhale Kurumunun gündemine alınması sonucu bir karar verilmiş
