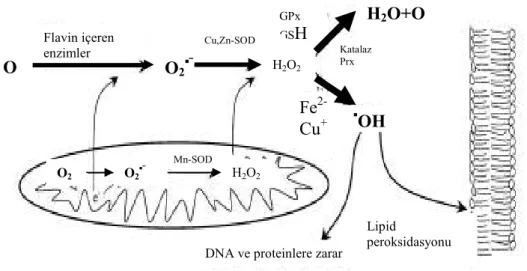
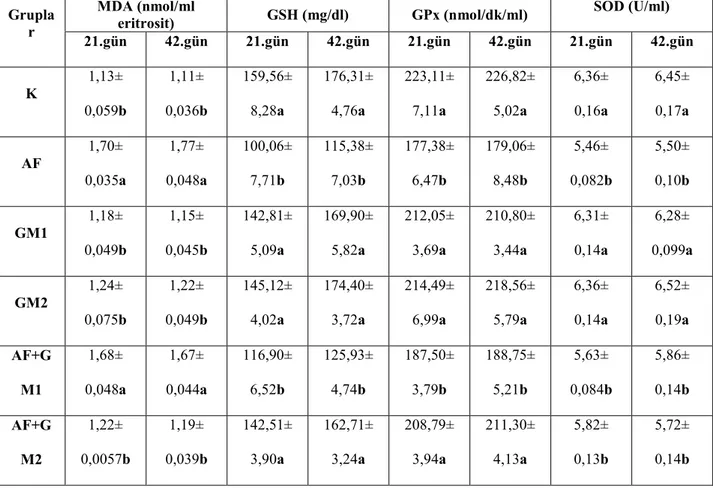
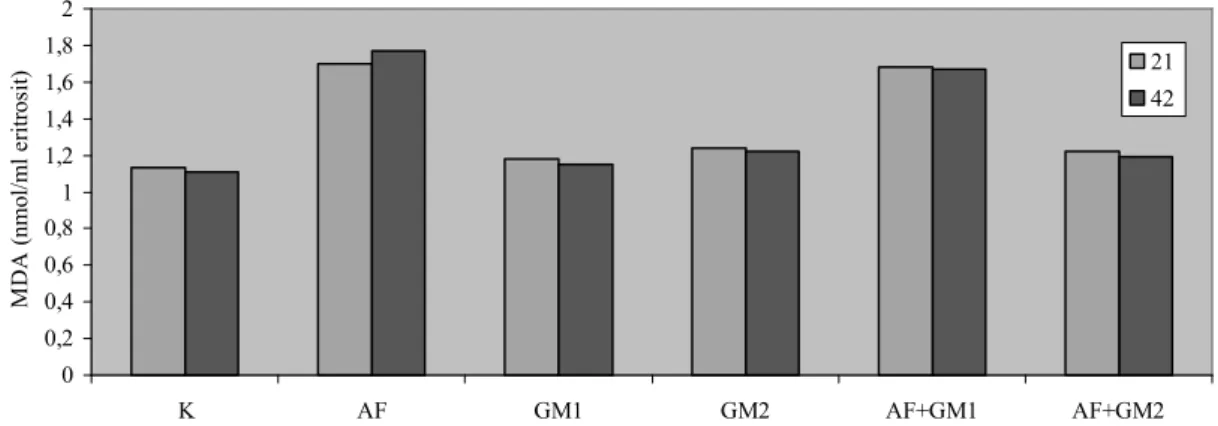
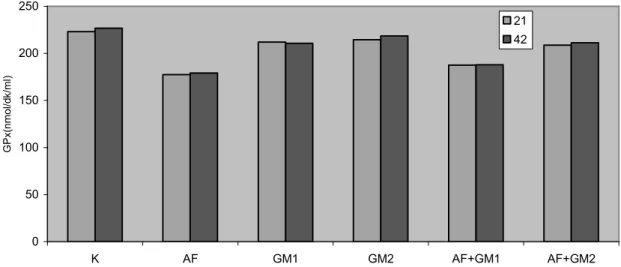
Japon bıldırcınlarında (coturnix coturnix japonica) deneysel olarak oluşturulan aflatoksikozis'te yeme katılan maya glukomannaninin bazı biyokimyasal parametreler ve antioksidan enzimler üzerine olan etkileri
Tam metin
Şekil
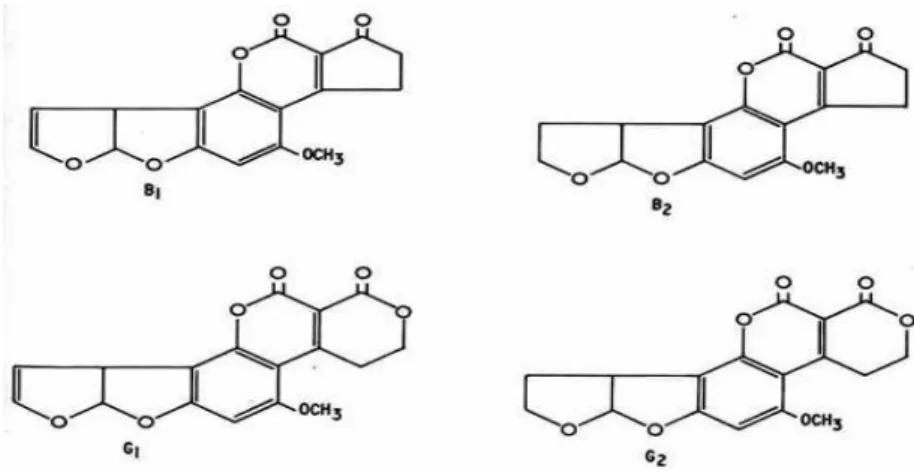
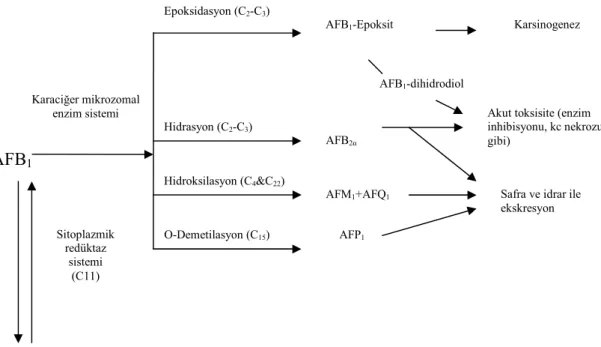
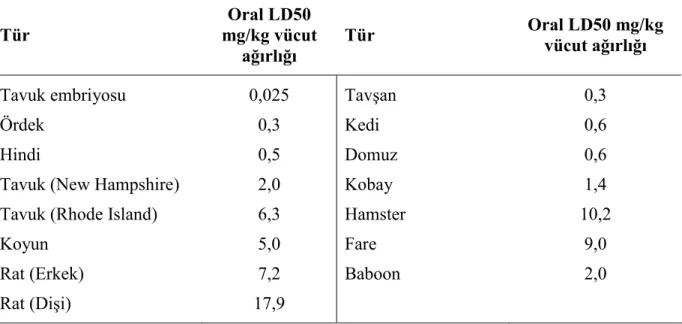
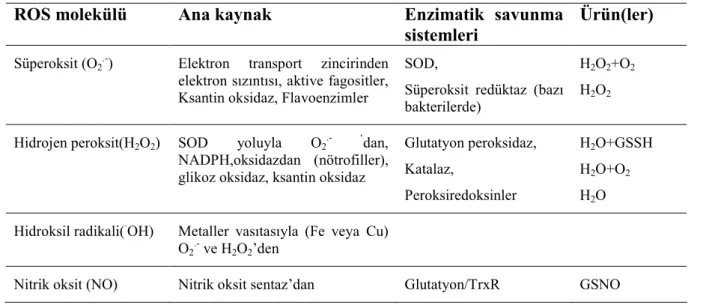
Outline
Benzer Belgeler
As a result of this study, Kepler’s laws the various alternative concepts of teaching students to have pre-determined and made for conceptual change students' alternative
Kendisinin y›l- lard›r incelemekte oldu¤u, Californi- a’daki Long Valley kalderas›n›n da 1997 y›l›nda 6 ay içinde 10 cm kadar yükseldi¤ini ve bu yükselmeye bir
İnsan, hür olarak doğarmış, fakat bugün her yanından bir çok bağ için de bulunuyormuş. Bu münasebetle söylenmiş kâfirce bir sözü
İlköğretimi tamamlamış öğrencilerin, fen ve teknoloji öğretmenlerine göre sürdürülebilir çevreye yönelik kavramsal anlamaları daha iyi düzeyde olduğu
Çalışmanın ilerleyen bölümlerinde, ülkemizdeki sivil hava taşımacılığında çağdaş eğilimler, stratejik planlama ilkeleri; havayolu işletmelerindeki düşük
Kan kültürü altın standart olarak kabul edilirken kültür sonuçlarının 24-48 saatten önce alınamaması, yanlış negatif ve pozitif sonuçların
5- Yandaki devrede zil çalarken 3 ve 4 nolu anahtarlar kapatılırsa hangi
la kitaplaştırılan Kıdwai’nin eser- lerinde genel olarak Osmanlı Devleti’nin son dönemleri ve özellikle de yakın tarihimiz için önemli bilgi ve ipuçları verilmek-
