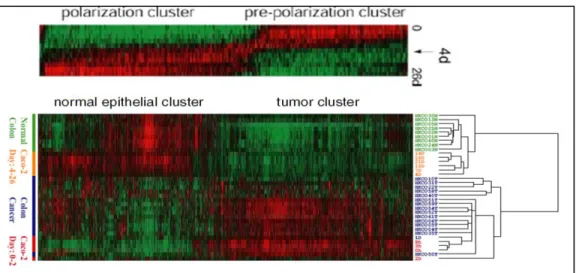
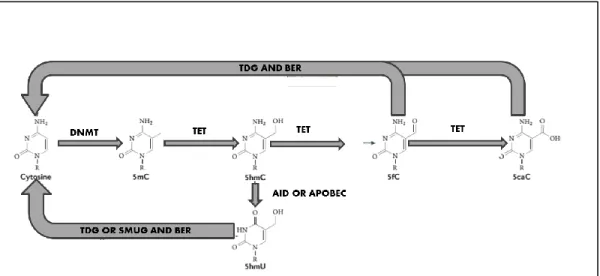
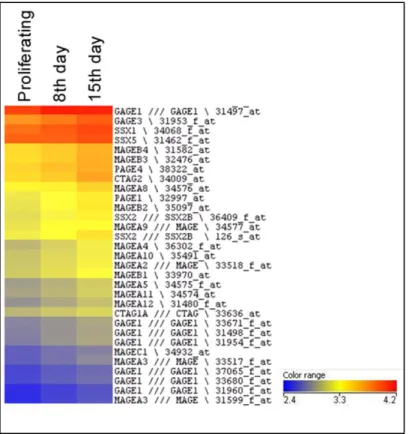
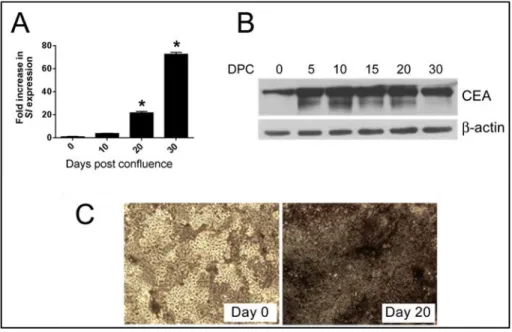
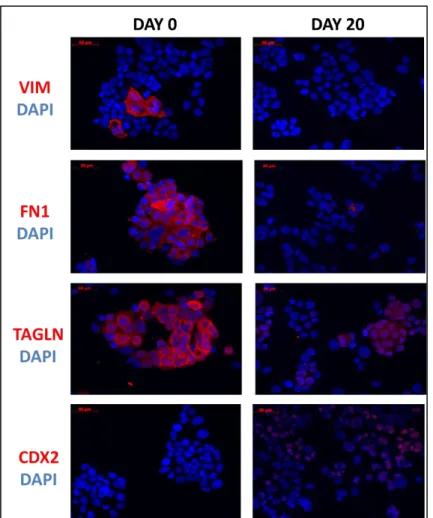
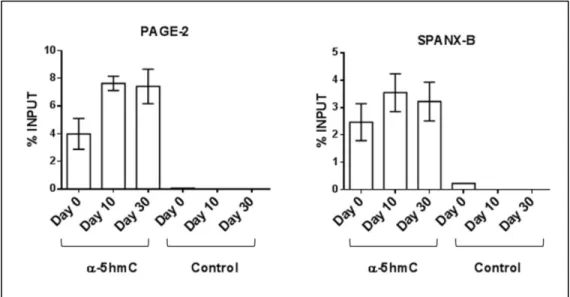
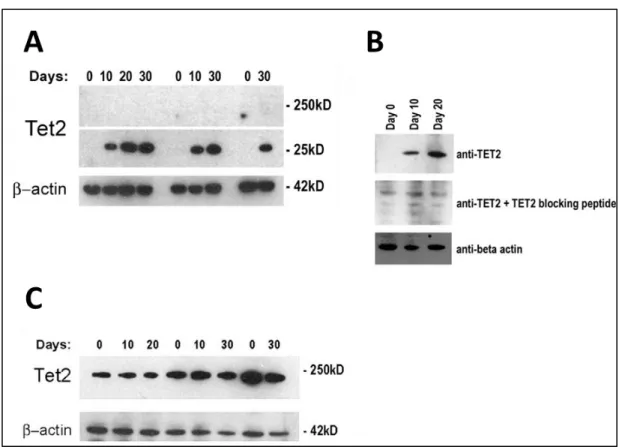
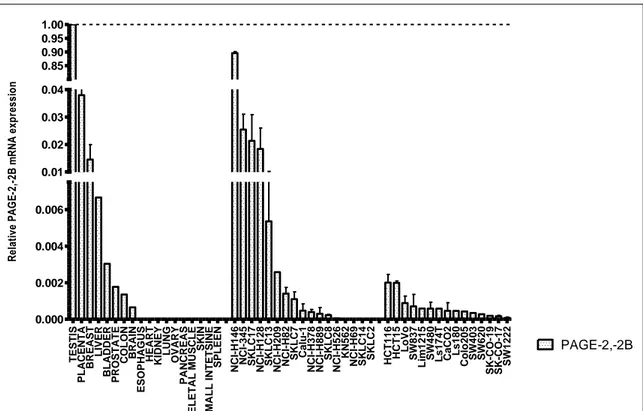
A novel role for 5-hmC in the regulation of cancer testis gene expression in cancer and mesenchymal to epithelial transition
Tam metin
Şekil


Benzer Belgeler
Kadın Dayanışma Vakfı'nm Ankara'daki gecekondularda yaşayan kadınlar arasında yaptığı bir araştırma, kadınların % 97'sinin kocalarının saldırısına uğradığı (1995);
Kaynaklarda bazı sahâbîlerin cuma öncesinde kıldıkları ifade edilen namaz, cuma ezanı ile kameti arasında kılınan cuma öncesi sünnetler değil zeval vaktinden önce
Thus, even if the products differ in their profit margins, the optimal firm assortments are such that a firm’s assortment is always a subset of another firm with an equal or
Finally, we use them to derive a num- ber of results in quantum information theory: from new bounds for quantum random access codes (QRACs) for both real and complex quantum systems
Mechanization and the so- called deskilling of many work processes, the continuing signifi cance of craft-based control and craft autonomy, the increas ing significance
In Shallow grating based experiments, we use copper sulfate solutions with different concentrations and the silver as a plasmon supporting layer. Although the
The MTT test did not indicate a significant growth inhibition in ZF4 cells following rapamycin treatment, however, rapamycin was observed to significantly downregulate zebrafish
This can be achieved by solving a multi-period optimization problem which minimizes the summation of production, setup, inventory holding, and regular and overtime capacity
