T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
KÜTAHYA VĠġNE ÇEġĠDĠNDE TOMURCUK VE ÇĠÇEKLERĠN SOĞUĞA DAYANIKLILIK
DÜZEYLERĠNĠN BELĠRLENMESĠ Omar Muhammed Ghanim ALNUAĠMĠ
YÜKSEK LĠSANS TEZĠ Bahçe Bitkileri Anabilim Dalını
Nisan-2019 KONYA Her Hakkı Saklıdır
TEZ KABUL VE ONAYI
Omar Mohammed Ghanim ALNUAĠMĠ tarafından hazırlanan ―Kütahya ViĢne ÇeĢidinde Tomurcuk ve Çiçeklerin Soğuğa Dayanıklılık Düzeylerinin Belirlenmesi‖ adlı tez çalıĢması 19/04/2019 tarihinde aĢağıdaki jüri tarafından oy birliği ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü Bahçe Bitkileri Anabilim Dalı’nda YÜKSEK LĠSANS TEZĠ olarak kabul edilmiĢtir.
Jüri Üyeleri Ġmza
BaĢkan
Prof. Dr. Ahmet EġĠTKEN
DanıĢman
Prof. Dr. Lütfi PIRLAK
Üye
Prof. Dr. Halil Ġbrahim OĞUZ
Yukarıdaki sonucu onaylarım.
Prof. Dr. Mustafa YILMAZ FBE Müdürü
TEZ BĠLDĠRĠMĠ
Bu tezdeki bütün bilgilerin etik davranıĢ ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalıĢmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all material and results that are not original to this work.
Omar Mohammed Ghanim ALNUAĠMĠ Tarih: 19.04.2019
iv
ÖZET
YÜKSEK LĠSANS TEZĠ
KÜTAHYA VĠġNE ÇEġĠDĠNDE TOMURCUK VE ÇĠÇEKLERĠN SOĞUĞA DAYANIKLILIK DÜZEYLERĠNĠN BELĠRLENMESĠ
Omar Muhammed Ghanim ALNUAĠMĠ Selçuk Üniversitesi Fen Bilimleri Enstitüsü
Bahçe Bitkileri Anabilim Dalı DanıĢman: Prof. Dr. Lütfi PIRLAK
2019, 49 Sayfa
Jüri
DanıĢman: Prof. Dr. Lütfi PIRLAK BaĢkan: Prof. Dr. Ahmet EġĠTKEN Üye: Prof. Dr. Halil Ġbrahim OĞUZ
Bu çalıĢma, 2018 yılında Konya Ģartlarında Kütahya viĢne çeĢidinde çiçek tomurcukları ve çiçeklerin kıĢ ve ilkbahar dönemlerinde düĢük sıcaklıklara dayanıklılık düzeylerinin belirlenmesi amacıyla yürütülmüĢtür. Çiçek tomurcukları ve çiçeklerin zarar gördüğü sıcaklıkların belirlenmesi için farklı dönemlerde alınan örnekler kademeli olarak sıcaklığı düĢürülen kabine yerleĢtirilip test edilmiĢtir. Testler, tomurcuk ve çiçekler 0 ile -22oC arasındaki sıcaklıklarda bekletilerek yapılmıĢ, uygulama sonrasında % 70 zarar gören tomurcuk ve çiçekler ölü olarak kabul edilmiĢtir. Bunun dıĢında alınan örneklerde toplam, bağlı ve serbest su, kuru madde, toplam karbonhidrat, protein, prolin ve besin elementi miktarları tespit edilmiĢtir.
Farklı sıcaklıklarda tomurcuk ve çiçeklerin ölüm oranları arasındaki farklar istatistiki olarak önemli bulunmuĢtur. KıĢ mevsiminde dona dayanıklılığı yüksek olan tomurcuklar ilkbahar mevsimine doğru bu dayanıklılığını kaybetmeye baĢlamıĢtır. Tomurcuk ve çiçeklerin % 50’sinden fazla zarar meydana gelen sıcaklıklar 29.01 tarihinde -16°C; 19.02 ve 05.03 tarihlerinde -15°C; 19.03 tarihinde -11°C; 04.04 ve 20.04 tarihlerinde ise -10°C olarak tespit edilmiĢtir. Buna göre Konya il merkezinde viĢne çiçek tomurcuklarının mart ayı sonuna kadar dondan zarar görme ihtimali bulunmakta, bu dönemden sonra ise zarar ihtimali azalmaktadır.
KıĢ mevsiminden çıkıĢla birlikte tomurcuklarda su içeriğin bariz bir Ģekilde artıĢ göstermiĢtir. Tomurcuklarda kıĢtan çıkıĢla birlikte bağlı su miktarında hızlı bir azalma,
v
buna karĢılık serbest su miktarında ise hızlı bir yükselme tespit edilmiĢtir. Kuru madde içerikleri ise 29.01–05.03 tarihleri arasında sabit kalırken, 19.03 tarihinde hızlı bir artıĢ meydana gelmiĢtir.
Örnek alma dönemleri arasında besin maddesi miktarları arasındaki farklar istatistiki olarak önemli bulunmuĢtur. Buna göre örneklerde N içeriği 29.01 ile 20.04 tarihleri arasında sürekli azalma göstermiĢtir. P, K, Mn, Zn ve S miktarları 29.01 tarihinden 19.03 tarihine kadar artmıĢ, bu tarihten sonra azalmaya baĢlamıĢtır. Mg ve Fe miktarları 19.03 tarihine kadar düzenli bir Ģekilde artmıĢ, bu tarihten sonra sabit kalmıĢtır. Ca ve Cu miktarları ise 05.03 tarihine kadar yükselmiĢ, daha sonra düĢmeye baĢlamıĢtır. B içeriği ise sürekli artıĢ göstermiĢtir. Zn miktarı 04.04 tarihine kadar düzenli olarak artıĢ göstermiĢ, daha sonra düĢmüĢtür. S içeriği ise farklı tarihlerde dalgalanma göstermiĢtir.
Örneklerde karbonhidrat içeriği bakımından tarihler arasında önemli farklar tespit edilmiĢtir. Buna göre karbonhidrat içeriği 29.01 tarihinden 04.04 tarihine kadar düzenli olarak azalmıĢ, 20.04 tarihinde ise tekrar yükselmeye baĢlamıĢtır. Protein içerikleri birbirini takip eden örnek alma dönemlerinde sürekli azalma göstermiĢtir. Prolin içerikleri arasındaki farklar istatistiki olarak önemli bulunmamıĢtır.
Anahtar Kelimeler: Kütahya viĢne çeĢidi, çiçek tomurcukları, çiçekler, soğuğa
vi
ABSTRACT
MS
Determination of Cold Resistance of Buds and Flowers on Kütahya Sour Cherry Cultivar
Omar Muhammed Ghanim ALNUAĠMĠ
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY
THE DEGREE OF MASTER OF SCIENCE
Advisor: Prof. Dr. Lütfi PIRLAK
2019, 49 Pages
Jury
Advisor: Prof. Dr. Lütfi PIRLAK President: Prof. Dr. Ahmet EġĠTKEN Member: Prof. Dr. Halil Ġbrahim OĞUZ
This study was carried out in 2018 in order to determine the resistance of flower buds and flowers to low temperatures in winter and spring periods in Kütahya sour cherry cultivar, in Konya conditions. In order to determine the temperature of the flower buds and flowers, the samples taken at different periods were gradually placed in the temperature-lowered cabinet and tested. Buds and flowers were tested, held at temperatures between 0 and -22oC. After the application, 70% or above of damaged buds were considered dead. In addition, total, bound and free water, dry matter, total carbohydrate, protein, proline and nutrient levels were determined.
Differences between the mortality rates of samples at different temperatures were statistically significant. In the winter season, buds with high frost resistance started to lose this resistance towards the spring season. Temperatures of more than 50% of buds and flowers are observed at 16°C on 29.01; 19.02 and 05.03 at 15°C; 19.03 at -11°C; it was determined as -10°C on 04.04 and 20.04. According to this, in the city center of Konya cherry flower buds are damaged by frost until the end of March and the probability of damage after this period is reduced. With the emergence of winter season, the water content of the buds has increased significantly. A rapid decrease in the amount of bound water was found in the buds with the emergence of winter and a rapid increase
vii
in the amount of free water. On the other hand, the dry matter contents remained stable between 29.01 and 05.03, and a rapid increase occurred on 19.03. The differences between the nutrient amounts were found to be statistically significant. According to this, the content of N in buds has decreased continuously between 29.01 and 20.04. The amounts of P, K, Mn, Zn and S increased from 29.01 to 19.03 and started to decrease after this date. Mg and Fe amounts increased regularly until 19.03 and remained stable after this date. Ca and Cu levels increased until 05.03, then began to fall. The B content increased continuously. The amount of Zn increased regularly until 04.04 and then decreased. S content fluctuated on different dates.
Significant differences were determined in terms of carbohydrate content in samples. According to this, the carbohydrate content decreased regularly from 29.01 until 04.04 and started to rise again in 20.04. Bud’s protein contents showed a steady decrease in successive sampling periods. The differences between the contents of the proline were not statistically significant.
viii
ÖNSÖZ
Yüksek lisans tezimin planlanıp yürütülmesinde yardımlarını gördüğüm danıĢman hocam Sayın Prof. Dr. Lütfi PIRLAK' a en içten Ģükranlarımı sunarım. ÇalıĢmamızda kullandığımız viĢne ağaçlarının temininde gerekli yardım ve kolaylığı sağlayan Sayın Prof. Dr. Ahmet EġĠTKEN ve çalıĢmamın her aĢamasında yardımlarını esirgemeyen Dr. Muzaffer ĠPEK, Dr. ġeyma ARIKAN, ArĢ. Gör. Murat ġAHĠN ve Bahçe Bitkileri Bölümü öğrencilerine ve öğrenim hayatım boyunca maddi ve manevi olarak sürekli yanımda olan aileme teĢekkürlerimi bir borç bilirim.
Omar Muhammed Ghanim ALNUAĠMĠ
ix ĠÇĠNDEKĠLER ÖZET ... iv ABSTRACT ... vi ÖNSÖZ ... viii ĠÇĠNDEKĠLER ... ix SĠMGELER VE KISALTMALAR ... x 1. GĠRĠġ ... 1 2. KAYNAK ARAġTIRMASI ... 3 3. MATERYAL VE YÖNTEM ... 9 3.1. Materyal ... 9 3.2. Metot ... 9 3.2.1.Arazi ÇalıĢmaları ... 9
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 16
4.1. Tomurcuk ve Çiçeklerin Öldüğü Sıcaklıkların Belirlenmesi ... 16
4.2. Örneklerin Toplam, Bağlı, Serbest Su ve Kuru Madde Miktarı ... 22
4.3. Bitki Besin Maddesi Tayini ... 24
4.4. Toplam Karbonhidrat Miktarı ... 27
4.5. Protein Miktarı ... 28
4.6. Prolin Miktarı ... 30
5. SONUÇLAR VE ÖNERĠLER ... 31
KAYNAKLAR ... 32
x SĠMGELER VE KISALTMALAR Simgeler °C Santigrad derece % Yüzde Kısaltmalar K Potasyum Cu Cakır Ca Kkalsiyum Zn Çinko P Fosfor Fe Demir N Azot S Kükürt B Brom Mg Magnezyum Mn Mangan g Gram mg Miligram kg Kilogram mM Milimolar mL Mililitre nm Nanometre
rpm Round per minute
µM Mikromolar
ÖS Öldürücü Sıcaklık
DTA Diferansiyel Termal Analiz NaOH Sodyum hidroksit
H2S04 Sülfürik asit AFP Antifriz Proteini
1. GĠRĠġ
ViĢne (Prunus cerasus L.) Rosaceae familyasına mensuptur. ViĢnenin anavatanı, Küçük Asya, Ġstanbul ve Hazar Denizi arasında uzanan Kuzey Anadolu Dağları'dır. ViĢne farklı iklim Ģartlarına iyi adapte olur. Anadolu'nun iç bölgelerinde hakim olan karasal iklim, sıcak ve kurak yazlara ve soğuk kıĢlara dayanmaktadır. Bu nedenle, çoğunlukla Anadolu'nun iç ve geçiĢ bölgelerinde yetiĢtirilmektedir (Özçağıran ve ark., 2003).
Dünya viĢne üretimi 1.378.216 ton olup, 192.500 ton üretim ile Türkiye, Rusya ve Polonya'dan sonra 3. sırada yer almaktadır (Anonim, 2018). Ülkemizde viĢne üretiminde ilk sırada yer alan il Afyonkarahisar olup, ardından Kütahya, Konya, Ankara ve Isparta gelmektedir (TUIK, 2018).
DüĢük sıcaklık bitki türlerinin yeryüzünde dağılımını belirleyen en önemli faktör olarak kabul edilir ve bahçe bitkilerinin verimini ve dağılımını sınırlayabilir (Kalberer ve ark., 2006). Bahçeciler yüksek kaliteli meyve üretimi ile ilgili olarak don problemiyle sürekli karĢı karĢıya kalmaktadır (Nybom, 1992). Meyve ağaçlarının soğuğa dayanıklılığı öncelikle genotip tarafından etkilenir. Üreme organlarına zarar veren kritik sıcaklıklar türler ve çeĢitler arasında farklıdır. Bununla birlikte, genotip etkilerini diğer faktörlerden ayırmak kolay değildir. Sonuç olarak, kritik sıcaklıkların sadece çiçek tomurcuklarının fenolojik aĢamasıyla değil; türler, çeĢitler, meyve bahçeleri ve hatta ağaçlar ile de değiĢtiği gözlemlenmiĢtir (Westwood, 1993).
Meyve ağaçlarının çiçek tomurcukları düĢük sıcaklık etkilerine karĢı dirençli iken, dinlenmeden sonra ilkbaharda dormansi dönemi sona erer, bitkiler büyümeye baĢlar ve soğuğa karĢı direnç azalır (Iezzoni ve Hamilton, 1985). Açık generatif tomurcukların ve yumurtalıkların hasarı sadece sıcaklığa (donma derecesine) değil, aynı zamanda donma süresine de bağlıdır. ViĢne generatif organlarının ilkbahar donlarına karĢı direnci ve bunu etkileyen faktörler hakkındaki veriler yetersizdir. Bu çalıĢmanın amacı viĢne çiçek tomurcuklarının kıĢ ve ilkbahar donlarına karĢı dayanıklılığını incelemektir.
Ülkemizin bulunduğu iklim bölgesi nedeniyle meyve türleri zaman zaman ilkbahar donlarından zarar görmektedir. Önceden bu zararlar erken çiçek açan badem, kayısı ve şeftali meyve türlerinde daha yaygın görülürken, son yıllarda küresel iklim değişikliği nedeniyle geç çiçek açan meyve türlerinde de zararlara neden olmaktadır. İç Anadolu Bölgesi ve Konya ilinde yaptığımız gözlemlerde son yıllarda ilkbahar
donlarının elma, armut, ayva ve vişne gibi geç çiçek açan meyve türlerinde de zarara yol açtığı tespit edilmiştir.
Bölgelere uygun meyve tür ve çeşitlerini belirleyerek üreticilere, ülke ekonomisine, tarımsal ürünleri sigortalayan kurum ve kuruluşlara finansal yük oluşturan don hasarını azaltmak mümkün olabilir. Bu amaçla, meyve tür ve çeşitlerinin farklı fenolojik dönemlerdeki soğuğa duyarlılığı belirlenerek, her bölgeye uygun meyve tür ve çeşitlerinin belirlenmesi ve önerilmesi mümkündür. Böylece, büyük verim kayıplarına yol açan don olaylarının etkisi belirli bir ölçüde azaltılabilir.
Bu çalıĢmanın amacı, ülkemiz tarımının önemli bir parçası olan vişnelerdeki çiçek tomurcuklarının soğuk toleransını belirlemek ve soğuk toleransı etkileyen bazı faktörleri incelemektir.
2. KAYNAK ARAġTIRMASI
Don zararının ana etkisi düĢük ortam sıcaklığından ziyade hücre suyunun donmasından kaynaklanır. Buzlanma meydana geldiğinde, kısa bir süre sonra hücreler arasındaki boĢlukta oluĢan buz kristalleri hücreler arası boĢluklardaki su potansiyelini ve hücreler arasındaki su akıĢını azaltır. Bu nedenle hücreler susuz kalır ve zarar görür. Donmaya dayanıklı tür ve çeĢitler dondan sonra hayatta kalabilirler. Ancak, buzlanma devam ederse ve simplastta buz geliĢirse bu tür ve çeĢitler de zarar görecektir (Lösch, 2001).
Bitkilerin soğuğa dayanıklılığı yetiĢtirildiği bölgeye, yetiĢtirme Ģartlarına, yaĢ ve taç Ģekline bağlı olarak aynı bitkide bile tür ve çeĢide göre değiĢmektedir. Örneğin, elma çiçek tomurcukları kıĢın ortasında -40°C'ye kadar dayanabilir (Ma ve ark., 2013), ancak ilkbaharda açıldığında daha yüksek (-2°C) sıcaklıklarda zarar görebilirler. Meyve türlerinde soğuğa dayanıklılığı etkileyen bir diğer faktör ise, ağaçların kıĢa hazırlandığı sonbaharda uygulanan gübre ve sulama rejimleridir. Topraktaki yetersiz besinler bitki örtüsünün kıĢa hazırlanmasını engelleyecek gübreleme ve sulama uygulamaları ya da kıĢ mevsimi için hazırlıkların gecikmesi dona dayanım gücünü azaltır. Ayrıca donmadan önce meydana gelen kuraklık da dona karĢı direnci azaltır (Snyder ve Connell, 1993). Sıcaklık 0°C'nin altına düĢmeden önce ortaya çıkan yüksek sıcaklıklar da bitkilerin donma eğilimini azaltır. 0°C'nin hemen üzerindeki sıcaklıklar donma direncinin artmasına neden olabilir (AslantaĢ ve ark., 2010).
Bitkilerde dona dayanma fotosentez gibi karmaĢık biyokimyasal ve fizyolojik bir olgudur (Klimov ve ark., 1999). EriĢ (1982) de dona dayanım ve düĢük sıcaklıklara uyum konusunun bitkilerde son derece karıĢık fizyolojik ve biyokimyasal olayların sonucu olarak ortaya çıktığını belirtmiĢtir. Aklimasyon sırasında su ve suda çözünür içerikli dokular, hücre zarlarındaki lipid kompozisyonu, antioksidan enzim aktivitesi, glutatyon miktarı gibi pek çok değiĢiklik meydana gelir. Bu değiĢikliklerin bir sonucu olarak bitki dokuları dona karĢı dirençli hale gelebilir (Kocsy ve ark., 2000). Bitki dokularında meydana gelen bu değiĢimler türlere ve çeĢitlere göre değiĢir. Meyve türleri ile çeĢitleri arasındaki duyarlılık farklılıklarının açıkça görülebileceği zaman erken kıĢ dönemidir (Lapins, 1962). ġeftali çiçek tomurcukları kıĢın ortasında zarar görebilir (Szymajda ve Żurawicz, 2016). Ayrıca, farklı fenolojik periyotların tomurcukların maruz kaldığı düĢük sıcaklıklardan da farklı olduğu için bu türlerin ve hatta çeĢitlerin dayanma sıcaklıklarını belirlemek gerekir.
Tüm bitkinin ve özellikle çiçek tomurcuklarının su içeriği meyve türlerinin soğuk dayanıklılığını belirleyen önemli bir faktördür. Birçok araĢtırıcıya göre su içeriği soğuğa dayanıklılıkla ters iliĢkilidir ve bitki dokularındaki su içeriği arttıkça soğuktan kaynaklanan zarar oranı artmaktadır (Andrews ve Proebsting Jr, 1987; Lu, 1992; Sakai ve Larcher, 2012). Buna karĢılık bitki dokularındaki kuru madde içeriğindeki artıĢla birlikte dona karĢı direncin arttığı bildirilmiĢtir (Gusta ve Fowler, 1976; Dalmannsdóttir ve ark., 2001; Setter ve Waters, 2003). Ayrıca soğuğa duyarlı türlerde kuru madde biriktirme kabiliyetinin zayıf olduğu, yani su içeriğinin yüksek olduğu da bilinmektedir.
Bitki besin maddelerinin soğuk toleransını etkilediğini bildiren çalıĢmalar vardır. Özellikle, fosfor ve potasyumun soğuğa mukavemeti artırdığı rapor edilirken, aĢırı azotun antosiyaninlerin sentezini azaltarak dayanma gücünü azalttığı bildirilmiĢtir (Solovieva, 1977).
Çiçek tomurcuklarının soğuğa direnci açılmamıĢ tomurcukların besin içeriğiyle de iliĢkilidir. Bazı araĢtırıcılara göre azot, fosfor, potasyum, sorbitol, glukoz, hidrofilik ve asidik amino asitlerin fazlalığı soğuk direncinin nedeni olabilir (Khanizadeh ve ark., 1992). Benzer Ģekilde, önceki yıllarda meyve ağaçlarında besin maddesi birikimi tomurcukların soğuk toleranslarını etkileyebilir (Rodrigo, 2000).
Proteinlerin soğuğa karĢı dirençte etkili olduğunu bildiren çalıĢmalar da vardır. DüĢük sıcaklığa maruz kaldığında bitkilerde çözünebilir protein miktarının arttığı, bunların soğuğa dayanıklı tür ve çeĢitlerde daha yüksek olduğu bildirilmektedir. Proteinlerin bu etkisi etrafında su tutulmasından ve hücre duvarından su kaybının önlenmesinden kaynaklandığı bildirilmiĢtir (Doğru ve Balkaya, 2015).
Lipitlerin bitkilerde soğuğa toleransı etkilediği uzun süredir bilinmektedir ve dona dayanıklılığı etkileyen en önemli faktörler arasındadır (Szalai ve ark., 2001). Hücre zarlarındaki doymamıĢ yağ asitlerinin miktarı azalan sıcaklık ile artar ve çoğu durumda doymamıĢ hücre zarlarının lipit direnci donun kuvvetiyle iliĢkilidir (Nishida ve Murata, 1996). Lipidler hücre zarının kimyasal ve fiziksel yapısını değiĢtirerek hücre dıĢı donma sırasında hücrelerden su kaybını önleyerek hücre zararını önler (Steponkus ve Webb, 1992). DoymamıĢ yağ asitlerinin miktarındaki bu artıĢ doymuĢ yağ asitlerinin doymamıĢ yağ asitlerine yağ asidi desatürazı ile dönüĢtürülmesine bağlanır, bu da bitkinin aklimasyonu ile artar (Sakamoto ve Murata, 2002).
Bazı Ilıman Ġklim Meyve Tür ve ÇeĢitlerinin Soğuğa Dayanıklılığı Üzerinde ÇalıĢmalar
Ġlkbahar donları viĢnede verim için sınırlayıcı bir faktördür. Litvanya Tarım ve Orman AraĢtırma Merkezi Bahçe Bitkileri Genetik ve Biyoteknolojisi Enstitüsünde yapılan çalıĢmada viĢne çeĢitlerinin sürgünleri 30 dakika süreyle -4°C'de tutulmuĢ ve fenolojik dönemlerdeki dayanıklılık ve ilkbaharda zarar görme ihtimalleri değerlendirilmiĢtir. ÇeĢitler farklı fenolojik dönemlerde ilkbahar donlarına karĢı dayanıklılığa göre iki gruba ayrılmıĢtır. Birinci grup çiçek tomurcuğu fenolojik döneminde dona karĢı en hassas olan bitkilerden oluĢmaktadır. Ġkinci grup ise tam çiçeklenme döneminde çoğunlukla hassas olan çeĢitlerden oluĢmuĢtur. ViĢne çeĢitlerinin ilkbahar donlarına karĢı direncinin çeĢitli biyolojik bileĢenlerin bir kombinasyonu olduğu belirlenmiĢtir. Ġlkbaharda dona karĢı viĢne direnci donma döneminde ortaya çıkan tomurcukların geliĢim düzeyine bağlıdır. AraĢtırmada Orkolija çeĢidinin bitkileri diğer fenolojik fazlara göre tam çiçek fazında −4°C'de en dayanıklı olarak bulunmuĢtur (Rutkowski ve ark., 2015).
Edgerton (1954), Proebsting Jr ve Fogle (1956), Proebsting Jr (1959), Campbell ve Hadle (1960), gibi araĢtırıcılar değiĢik Ģeftali çeĢitlerinin çiçek tomurcuklarını dinlenme döneminde değiĢik sıcaklık derecelerine maruz bırakarak bunların dona dayanım mekanizmalarını incelemiĢlerdir. Dona dayanım bakımından çeĢitler arasında büyük farklılık bulunurken, bir çeĢidin değiĢik sıcaklık derecelerine dayanımlarının da farklı olabileceği ve tomurcuğun tam dinlenme safhasında dona dayanıklığının yüksek olduğu vurgulanmıĢtır.
Edgerton (1960), Ģeftali ağaçlarının dona dayanıklılıklarının sonbahar ve kıĢ aylarında düĢük sıcaklıkla arttığını, dinlenme periyodundan sonra ılık havada ise azaldığını bildirmiĢtir.
Proebsting (1963), çiçek tomurcuklarının düĢük sıcaklık derecelerinden çok, düĢük sıcaklığın devam etme süresinden etkilendiklerini belirtmiĢtir.
Zavarzin (1963), Ģeftali, kayısı ve kirazların dona dayanımlarını saptamak için bunları Ģubat ve mart aylarında 10˚C ve 15˚C’de 68 saat süreyle tutmuĢtur. Sonuçta -10'de Ģeftali tomurcuklarının tamamının, kaysı tomurcuklarının % 88.8'inin ve kiraz tomurcuklarının % 96'sının zararlandığı saptanmıĢtır.
Burak (1989), beĢ Ģeftali çeĢidinde yaptığı don testi çalıĢmalarında tüm çeĢitlerin dona dayanıklılıklarının ocak ve Ģubat aylarında yüksek, kasım ve mart aylarında ise düĢük olduğunu saptamıĢtır.
Szabó (1990), 35 Ģeftali ve nektarin çeĢidinin meyve veren dallarındaki soğuk zararını soğuklardan hemen sonra değil, daha sonraki dinlenme periyodunda 0-5 ıskalası kullanarak ölçmüĢ ve bu ölçümlerde ksilemin en az % 10 oranında kahverengileĢtiğini, bunun % 76 ve daha yukarıya kadar çıktığını saptamıĢtır.
Weaver ve Jackson (1969), Ģeftalilerde yaptıkları çalıĢmalar sonucunda, sıcaklığın düĢüĢ hızının dondan zararlanma üzerinde önemli rol oynadığını vurgulamıĢlardır.
Kiraz tomurcukları ve çiçekleri genellikle ilkbaharda meydana gelen don olayları nedeniyle zarar görür (Dexter, 1933; Smeeton, 1963; Rodrigo, 2000; Cittadini ve ark., 2006). Ġlkbaharda donma sıklığı ve donma sırasında ve sonrasında donma sıcaklığının altındaki sıcaklıkların sıklığı sadece tomurcukları değil, aynı zamanda meyve ve sürgünleri de etkiler. Çiçek tomurcukları dinlenmeden meyve tutumuna kadar düĢük sıcaklıklara karĢı giderek artan bir hassasiyet gösterirler (Longstroth ve Perry, 1996; Rodrigo, 2000). Meyve bahçesi yönetim kararlarının çoğu düĢük sıcaklıklardan zarar görme tehlikesine göre alınır (Proebsting ve Mills, 1971; Longstroth ve Perry, 1996). DüĢük sıcaklık oluĢumundan önce veya hemen sonra meydana gelen çevre
Ģartları donma hasarının derecesine büyük ölçüde etki eder. Tomurcuklanma evresi ile çakıĢan bir ilkbahar donması ciddi zararlanmalara yol açabilirken, tomurcuk uyanmasının daha ileri aĢamalarında hassasiyet giderek artar. Ġlkbahar donları sırasında hücresel hasar genellikle hücre dıĢı donma ile meydana gelir ve çeĢitli morfolojik değiĢikliklerde yeniden ortaya çıkabilen bir dizi anatomik etki ile sonuçlanır (Rodrigo, 2000).
Kirazlarda sonbaharın sonlarında ve tomurcukların aklimatize edilmesinden önce ve özellikle de tomurcukların kabarması ve açılması sırasında donma hasarı meydana gelebilir. Bu süre zarfında tomurcukların dayanıklılık seviyelerinde önemli ölçüde değiĢme meydana gelir. Kirazların çiçek tomurcukları vejetatif tomurcuklara göre donmaya daha yatkındır. Genç kiraz ağaçlarının çiçekleri dona karĢı aĢırı derecede hassastır (Proebsting Jr ve Mills, 1978b).
Ġlkbaharda tomurcuk geliĢimi sırasında sertlik seviyesindeki düĢüĢü yansıtan geliĢme aĢamalarının her biri için kiraz çeĢitlerinin kritik sıcaklık ve don duyarlılıklarının bilinmesi hassas bölgelerde bahçe kurulmasına yardımcı olabilir. Diferansiyel termal analiz (DTA), bazı türlerin odunsu dokularında soğuğa dayanımı belirlemek için kullanılan yaygın yöntemlerden biridir. Elma (Quamme ve ark., 1972a; Quamme ve ark., 1972b), armut (Montano ve ark., 1987) ve açelya (Graham ve Mullin,
1976), uykuda odunsu bir düĢük sıcaklıklı ekzoterm (LTE) DTA ile saptanan gövde kesitleri soğutma sırasında meydana gelen hem ksilem hem de floem zararlanması ile iliĢkili bulunmuĢtur (Volk ve ark., 2009).
Ġlkbahar süresince soğuğa dayanım hem çevrenin hem de tomurcuk döneminin bir fonksiyonudur (Mexal ve ark., 1991; Longstroth, 2013) ve donma sıcaklıklarında meyve ürünlerinin direnci farklıdır (Rodrigo, 2000; Caprio ve Quamme, 2006; Longstroth, 2013). ÇeĢitler arasında yıldan yıla değiĢiklik ve farklılıklar kirazın meyve tutumunda genotip ve hava Ģartlarının güçlü bir etkisi olduğunu göstermiĢtir (Andersen ve L, 2001; García-Montiel ve ark., 2010).DüĢük bir sıcaklık sonrası veya öncesindeki çevre Ģartları donma zararının derecesini büyük ölçüde etkiler. Ġlkbahar donu bitkiyi tomurcukların patlamasının daha ileri evrelerinde ve küçük çiçek halinde iken gittikçe hassaslaĢtırırken, tomurcuğun patlama evresi ile aynı zamana denk gelen bir ilkbahar donu ciddi zarara neden olabilir.
Donma sıcaklıklarının etkilediği dinlenme halindeki çiçek tomurcuklarında zarar aynı değildir,çünkü buz kristalleri sadece bazı çiçek dokularında oluĢmuĢtur. Sıcaklık kademeli olarak azalırsa bir ağacın zarara direnci daha iyidir ve zararın çoğuna neden olan sıcaktan soğuğa doğru dik bir düĢüĢle karĢılaĢtırıldığında ĢaĢırtıcı derecede düĢük sıcaklıklara dayanabilir. Farklı ağaç türlerinin donma sıcaklığına nasıl tepki vereceği büyümenin gerçekleĢtiği yere bağlıdır ve hücreler sanki buz kristali oluĢumunu önlemek için antifriz sistemler içine kurulmuĢtur. Bu tolerans ayrıca dondurma iĢleminden etkilenmiĢ olabilir (Mazur, 1969; Ashworth ve Wisniewski, 1991; Palonen ve Buszard, 1997).
Bitkilerin zarara sebep olan kritik sıcaklıklardan etkilenmesi ve kısa süreli çevre sıcaklığına adapte olabilmesi sebebiyle çevre ve bir ürün dikkate alındığında bu kritik sıcaklıkların tahmin edilmesi yanıltıcıdır (Snyder ve ark., 2012).
Donma toleransı kritik dönemlerde bitki canlılığı için önemli bir özelliktir. Donma hasarının meydana geldiği ölümcül sıcaklığı tanımlar (Gray ve ark., 1997). Sıcaklık 0◦C'nin altındaysa soğuk zararlanması meydana gelebilir. Donma zararı bitki dokusunun içindeki buz oluĢumundan ve buz kristallerinin hücre zarı sistemlerine zarar vermesinden kaynaklanır (Westwood, 1978).
Donmalar meyve tomurcuklarına ve elmanın genç meyvelerine zarar verebilir ve hasar seviyesi donma yoğunluğu ve süresi ile birlikte donma meydana geldiği zamanın geliĢim aĢaması ile doğrudan iliĢkilidir (Proebsting Jr ve Mills, 1978a; 1978b; Iezzoni ve Hamilton, 1985). Sıcaklık düĢüĢü kademeli ise ağaçlar zararlanmaya direnmek için
daha iyi durumdadır. Bununla birlikte aniden sıcaklık düĢüĢü ile hızlı bir donma meydana gelirse uzamıĢ bir donma ağacın ölümüne veya zarar görmesine neden olabilir (Rodrigo, 2000).
Elma ve armut tomurcukları dinlenme döneminde nadiren zarar görür, sıcaklık aniden düĢerse ve aĢırı düĢük sıcaklıklar meydana gelirse tam çiçeklenmeye yaklaĢan tomurcuklarda zarar meydana gelebilir (Proebsting, 1963; Proebsting, 1982). Ancak, tomurcuklanma sırasında ve sonrasında düĢük sıcaklıklar sadece tomurcukları değil, aynı zamanda meyvelere ve hatta sürgünlere de zarar verebilir (Smeeton, 1963; Rodrigo, 2000; Atkinson ve ark., 2013). Tomurcuk patlamasının daha ileri aĢamalarında tomurcuklar ve küçük meyveler dona karĢı giderek daha hassas hale gelirler (Rodrigo, 2000).
Soğuğa dayanıklılık bitkinin donma sıcaklığına uyum sağlaması ve buna dayanmasıdır (Gusta ve Wisniewski, 2013; Wisniewski ve Gusta, 2014). Tomurcuklar ve dallar gibi farklı bitki dokuları kıĢın dinlenme süreleri boyunca aĢırı sıcaklıklara farklı cevaplar verirler (Ketchie, 1984). Dinlenme sırasında donma sıcaklıklarının tomurcuklar üzerindeki etkisi farklıdır ve bu tomurcuklar çok düĢük sıcaklıklara dayanabilir, ancak dinlenme süreleri tamamlandıktan sonra tomurcuklar kabarmaya ve geliĢmeye baĢlar, hava Ģartlarına daha duyarlı hale gelir ve bu süre zarfında soğuk veya donma sıcaklıklarından kolayca zarar görebilir. Dinlenme süresinden sonra canlı kalan tomurcuklar çiçeklenmeye devam eder (Proebsting Jr ve Mills, 1978b).
Soğuğa dayanıklılıktaki yıldan yıla değiĢkenlik iklim Ģartlarındaki geçici değiĢkenlik ile ilgilidir. Bahçedeki farklı ağaçlar veya ağacın içindeki farklı bölümler mikro iklim farklılıklarından dolayı etkilenebilir veya meydana gelen donma türü değiĢebilir (Glozer, 2010).
3. MATERYAL VE YÖNTEM 3.1. Materyal
AraĢtırma 2018 yılında Selçuk Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümünde yürütülmüĢtür. Bitki materyali olarak Selçuk Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü AraĢtırma ve Uygulama Bahçesinde bulunan mahlep anacı üzerine aĢılı 13 yaĢında, verim çağında Kütahya viĢne çeĢidi ağaçları kullanılmıĢtır.
Kütahya viĢne çeĢidi ülkemizin en önemli viĢne çeĢididir. Nisan-Mayıs aylarında çiçeklenir. Yaprakları parlak, tüysüz ve kısa saplıdır. Meyve parlak koyu kırmızı, yuvarlakça kalp Ģeklinde ve uç tarafı küttür. MayhoĢ, sulu ve aromalıdır. Çok iyi kalitededir. Ağaçları çok verimlidir (Özçağıran ve ark., 2003).
ġekil 3.1. ViĢne tomurck örneklerinin alındığı bahçenin görünümü (Orijinal)
3.2. Metot
3.2.1.Arazi ÇalıĢmaları
ÇalıĢma ve uygulamalar meyve verme çağında olan ve aynı iklim ve bakım Ģartlarında bulunan bitkilerden alınan sürgünler üzerinde yapılmıĢtır. Örnek alınan bitkiler aynı anaç üzerine aĢılanmıĢ, aynı yaĢ ve büyüklüktedir. Kütahya viĢne çeĢidinden 25 adet bitki belirlenmiĢ ve örnekler bu bitkilerden alınmıĢtır. ÇalıĢma, 1 kıĢ ve ilkbahar dönemini kapsayacak Ģekilde yürütülmüĢ ve don testleri yapılacak örnekler 15’er gün arayla alınmıĢtır (Felipe, 1977).
Örnekler ağaçların orta kısımlarından yerden en az 1 m yükseklikten ve sabah saat 10-11 arasında alınmıĢtır (Pramsohler ve Neuner, 2013). Örnekler mümkün olduğunca tüm ağaçların aynı bölgelerinden ve aynı uzunlukta olacak Ģekilde alınmıĢtır. Alınan örnekler soğuk uygulaması yapılacak zamana kadar exoterm sıcaklıklarının değiĢmemesi için (en fazla 12 saat) +4oC sıcaklıkta tutulmuĢtur (Quamme, 1983). Her dönemde don ve canlılık testleri, bünyesel organik ve inorganik maddelerin belirlenmesi için 300 adet çiçek tomurcuğu alınmıĢtır.
ġekil 3.2. ViĢne örneklerinin soğuğa dayanıklılık çalıĢması (Orijinal)
3.2.2.1.Tomurcukların Öldüğü Sıcaklıkların Belirlenmesi
Çiçek tomurcuklarının zarar gördüğü sıcaklıkların belirlenmesi için farklı dönemlerde çiçek tomurcuğu ve çiçek örnekleri alınmıĢtır. Örnek alım tarihleri aĢağıda verilmiĢtir;
29.01.2018 1. Tomurcuk dönemi 19.02.2018 2. Tomurcuk dönemi 05.03.2018 3. Tomurcuk dönemi 19.03.2018 4. Tomurcuk dönemi
04.04.2018 1. Çiçek dönemi (Çiçeklenme baĢlangıcı) 20.04.2018 2. Çiçek dönemi (tam Çiçeklenme)
Alınan örnekler kademeli olarak sıcaklığı düĢürülen kabine yerleĢtirilip test edilmiĢtir. Bu amaçla alınan örnekler en geç 1 gün içerisinde teste tabi tutulmuĢtır. Test yapılırken örneklerin sıcaklık dalgalanmalarından etkilenmemeleri için tomurcuklar alüminyum folyo içine yerleĢtirilmiĢtir (Wheeler ve ark., 2014). Testler örnekler 0 ile -22oC arasındaki sıcaklıklarda 1 saat bekletilerek yapılmıĢtır (Hacker ve ark., 2011; Reig ve ark., 2013). Uygulama sonrasında % 70 zarar gören örnekler ölü olarak kabul edilmiĢtir. Zarar tespiti örneklerin kesilmesiyle yapılmıĢtır (Martin ve ark., 2010). Kesit alınan örneklerin renkleri kontrol edilmiĢ, açık sarı ve yeĢil olanlar canlı, rengi koyulaĢıp kahverengiye dönenler ölü olarak kabul edilmiĢtir (Felipe, 1977; Hacker ve ark., 2011; Reig ve ark., 2013).
ġekil 3.3. Tomurcuk, çiçek ve küçük meyvelerden alınan kesitler (Orijinal)
ġekil 3.4. ViĢne tomurcuğu kesitleri: A ve B canlı, C ve D donma sunucu esmerleĢmiĢ doku Ģeklinde temsil edilir (Orijinal)
ġekil 3.5. ViĢne çiçeklerinden alınan kesitler (Orijinal)
3.2.2.2. Toplam, Bağlı ve Serbest Su Miktarlarının Belirlenmesi
Her bir aĢamada alınan 50 adet örneğin taze, hava kurusu ve fırın kurusu ağırlıkları alınmıĢtır. Elde edilen bu değerler aĢağıdaki formüllere iĢlenmiĢ ve böylece çiçek tomurcukları ve çiçeklerin serbest, bağlı ve toplam su içerikleri belirlenmiĢtir. Su içerikleri ile dona dayanım dereceleri kıyaslanarak örneklerin bulundurdukları serbest, bağlı ve toplam su içeriklerinin dona dayanım üzerine etkisi tespit edilmeye çalıĢılmıĢtır (Buchner ve Neuner, 2011).
Toplam Su Miktarı (%) = (Taze Ağırlık - Fırın Kurusu) / Fırın Kurusu Bağlı Su Miktarı (%) = (Hava Kurusu - Fırın Kurusu) / Fırın Kurusu Serbest Su (%) = Toplam Su Miktarı - Bağlı Su Miktarı
3.2.2.3.Toplam Kuru Madde Tayini
Örneklerin kuru madde oranlarının tespit edilmesi amacıyla her çeĢitten 50 adet tomurcuk ve çiçek sürgünün farklı noktalarından alınmıĢ, 70oC sıcaklığa ayarlanmıĢ etüvde tam kuruma sağlanana kadar kurutulmuĢtur (Reig ve ark., 2013). Kurutulan bu
örneklerin kuru madde miktarlarıyla soğuğa dayanıklılık arasında iliĢki olup olmadığı araĢtırılmıĢtır.
3.2.2.4. Tomurcuk ve Çiçeklerde Bitki Besin Maddesi Tayini
Bitki besin maddesi analizi için bitkinin orta kısımlarından alınan sürgünlerden 100 adet örnek analiz için oda sıcaklığında kurutulmuĢtur. Daha sonra havan kullanılarak toz haline getirilen örneklerde azot, fosfor, potasyum, kalsiyum, kükürt, bor, demir, bakır, çinko, mangan, magnezyum ve molibden miktarlarının tespiti için analiz yapılmıĢtır.
Bitki örneklerinin P, K, Ca, Mg, Fe, Mn, Zn, Cu, B ve S içerikleri nitrik asit-hidrojen peroksit (2:3) ile 3 farklı adımda (1. adım; 145ºC’de % 75 mikrodalga gücünde 5 dakika, 2. adım; 180ºC’de % 90 mikrodalga gücünde 10 dakika ve 3. adım 100ºC’de % 40 mikrodalga gücünde 10 dakika) 40 bar basınca dayanıklı mikrodalga yaĢ yakma ünitesinde tutulduktan sonra, P, K, Ca, Mg, Fe, Mn, Zn, Cu, B ve S tayini spektofotometrede okunmak suretiyle, N tayini de mikro Kjeldahl metodu ile belirlenmiĢtir (Mertens, 2005).
3.2.2.5. Toplam Karbonhidrat Tayini
Toplam karbonhidrat analizi (Daniels ve ark., 2007)’a göre yapılmıĢtır. Bunun için kuru tomurcuk örnekleri (0.1 g) bir test tüpünde 10-5 ml NaOH içinde su banyosuna yerleĢtirilmiĢtir. 10 dakika sonra, 5 ml damıtılmıĢ su eklenmiĢ ve protein içeriği elde edilmiĢtir. Kalıntılar su ile yıkanmıĢ ve oda sıcaklığında iki kez 30 dakika süreyle 10 ml % 67 H2S04 ile ekstrakte edilmiĢtir. Örneklerin uygun seyreltmesi gerçekleĢtirilmiĢ ve seyreltilmiĢ örnekler filtrelenmiĢtir. 1 ml numuneye, 1 ml fenol % 5 reaktifi eklenmiĢ ve daha sonra 5 ml konsantre edilmiĢtir. H2S04 örnek ile karıĢtırılmıĢ ve oda sıcaklığında 30 dakika beklemeye bırakılmıĢtır. Elde edilen renk bir reaktif kağıdına karĢı spektrofotometre kullanılarak 490 nm'de okunmuĢ ve toplam karbonhidratlar standart glikoz eğrisi kullanılarak hesaplanmıĢtır (Daniels ve ark., 2007).
ġekil 3.6. Örneklerde karbonhidrat analizi (Orijinal) 3.2.2.6. Protein Tayini
Protein tayini bitkilerin tomurcuk ve çiçeklerinden alınan 0,5 g’lık örnekler kullanılarak ―Bradford‖ metoduna göre yapılmıĢtır. Sonuçlar ―mg protein/g’’ taze doku cinsinden hesap edilmiĢtir. Küçük parçalara ayrılmıĢ örneklerinden 0.5 g alınarak 10 misli hacimdeki 0.05 M fosfat tamponunda (pH:6.5) havanda ezilerek homojenizasyon yapılmıĢtır. Homojenat dört katlı tülbentten süzülmüĢ ve süzüntü santrifüj tüplerine alınıp, 15.000 rpm’de 20 dk boyunca santrifüj edilmiĢtir. Protein tayini için tüplerin üst kısmındaki sıvı faz (süpernatant) kullanılmıĢ ve protein miktarı spektrofotometrik yolla tayin edilmiĢtir (Brasford, 1976).
3.2.2.7. Prolin tayini
Prolin tayini spektrofotometrik olarak asit-ninhidrin metoduyla belirlenmiĢtir (Bates ve ark., 1973). Bu iĢlem için saf prolin kullanılarak standart bir grafik çizilmiĢtir. 1 ml’sinde 200 μg prolin içeren stok çözeltiden tüplere 0.2, 0.4, 0.6, 0.8, 1.0, 1.2, 1.4, 1.6, 1.8 ve 2.0 ml alınarak her tüpün üzeri saf su ile 2 ml’ye tamamlanmıĢtır. Daha sonra tüplere 2 ml glasiyal asetik asit ve 2 mlasit-ninhidrin çözeltisi ilave edilmiĢtir. Bu örnekler hemen 100oC’ye ayarlı bir etüve alınmıĢtır. 1 saat sonra, buz banyosunda 10 dk tutularak reaksiyon durdurulmuĢtur. Her tüpe 4 ml toluen ilave edilip, bir vorteksle 20-30 sn karıĢtırılmıĢtır. Sonra her tüpte üstte kalan faz bir otomatik pipetle alınarak 520 nm’de absorbansları ölçülmüĢtür. Kör numune için toluen kullanılmıĢtır. Bu değerler kullanılarak standart grafik çizilmiĢtir. Örneklerdeki prolin tayini için ise 0,1g doku 10 ml % 3’lük sülfosalisilik asit içinde bir porselen havanda homojenize edilip, homojenat Whatman–42 filtre kâğıdıyla analitik bir huniden süzülmüĢtür. Süzüntüden 2 ml alınarak yukarıda standart grafik için anlatılan iĢlemlerden geçirilmiĢtir. Daha sonra elde edilen A520 değerleri standart grafik üzerinden μg prolin olarak belirlenmiĢtir. Bu değerler aĢağıdaki formülde yerine konulup μ molar / g taze yaprak prolin cinsinden hesaplanmıĢtır.
Prolin (µM) / Taze Yaprak (g) = (Prolin (µM) / ml x ml toluen) / (115,5 µg / mM) / (g doku/5)
4. ARAġTIRMA SONUÇLARI VE TARTIġMA
4.1. Tomurcuk ve Çiçeklerin Ölümine Sıcaklıkların Etkisi
Farklı tarihlerde alınan viĢne tomurcuk ve çiçek örneklerinin 0 - -22°C sıcaklıklar arasında tutulduklarında meydana gelen ölüm oranlarına ait sonuçlar Çizelge 4.1. ve ġekil 4.1.’de verilmiĢtir. Bütün dönemlerde farklı sıcaklıklarda örneklerin ölüm oranları arasındaki farklar istatistiki olarak önemli bulunmuĢtur. Tomurcuklarının soğuklara dayanıklılığı içinde bulundukları geliĢme periyoduna göre değiĢmektedir. Bu tamamen bitkinin içindeki metabolik değiĢimlerden kaynaklanmaktadır. Örneğin, dinlenme halindeki hücrelerde Ģeker oranının ve proteinlerin artıĢı hücre içindeki buz oluĢumunu azaltarak dona dayanıklılığı artırmaktadır (Küden ve ark., 1998).
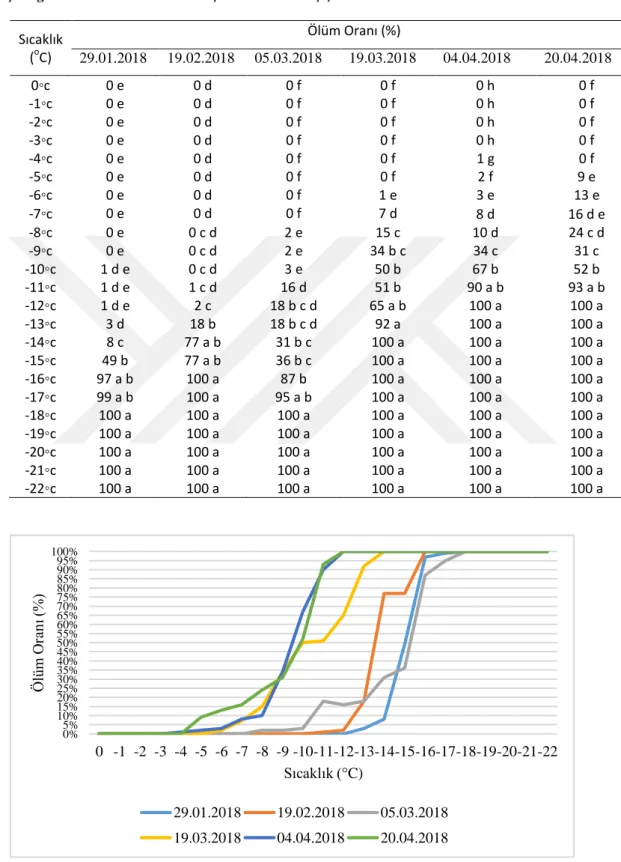
Buna göre 29.01.2018 tarihinde alınan örneklerde 0 ile -9°C arasında ölüm meydana gelmemiĢtir. -10°C’den itibaren örneklerde ölüm baĢlamıĢ; ölüm oranları -10 ile 12°C arasında % 1; 13°C’de % 3; 14°C’de % 8; 15°C’de % 49; 16°C’de % 97; -17°C’de % 99 ve -18°C’den itibaren % 100 olarak tespit edilmiĢtir.
19.02.2018 tarihinde alınan örneklerde -11°C’ye kadar ölüm meydana gelmemiĢtir. 11°C’den itibaren örneklerde ölüm meydana gelmiĢ; ölüm oranları -11°C’de % 1; -12°C’de % 2; -13°C’de % 18; -14 ve -15°C’de % 77; -16°C’den itibaren % 100 olarak bulunmuĢtur.
05.03.2018 tarihinde alınan örneklerde -7°C’ye kadar ölüm meydana gelmemiĢ, 8°C’den itibaren örneklerde ölüm meydana gelmeye baĢlamıĢtır. Ölüm oranları 8 ve 9°C’lerde % 2; 10°C’de % 3; 11°C’de % 16; 12 ve 13°C’de % 18; 14°C’de % 31; -16°C’de % 87; -18°C’den itibaren % 100 olarak bulunmuĢtur.
19.03.2018 tarihinde alınan tomurcuklarda -6°C’ye kadar ölüm meydana gelmemiĢtir. -6°C’de tomurcukların % 1’i ölmüĢ, bu sıcaklıktan sonra ölüm oranları düzenli olarak artıĢ göstermiĢtir. Ölüm oranları -7°C’de % 7; -8°C’de % 15; -9°C’de % 34; -10°C’de % 50; -11°C’de % 51; -12°C’de % 65; -13°C’de % 92 ve bu sıcaklıktan sonra % 100 olarak belirlenmiĢtir.
04.04.2018 tarihinde örneklerin dona hassasiyetleri artmaya devam etmiĢtir. Nitekim bu dönemde alınan çiçeklerde ilk zararlanma sıcaklığı -4°C (% 1 ölüm) olarak belirlenmiĢ, sıcaklık düĢtükçe zarar oranı artıĢ göstermiĢtir. Ölüm oranları -5°C’de % 2; -6°C’de % 3; -7°C’de % 8; -8°C’de % 10; -9°C’de % 34; -10°C’de % 67; -11°C’de % 90 ve bu sıcaklıktan sonra % 100 olarak belirlenmiĢtir.
20.04.2018 tarihinde alınan örneklerde ilk zararlanma -4°C’de meydana gelmiĢ (% 9 ölüm), yine diğer dönemlere benzer Ģekilde sıcaklık düĢtükçe zarar oranı artıĢ
göstermiĢtir. Ölüm oranları -6°C’de % 13; -7°C’de % 16; -8°C’de % 24; -9°C’de % 31; -10°C’de % 52; -11°C’de % 93 ve bu sıcaklıktan sonra % 100 olarak tespit edilmiĢtir.
Çizelge 4.1. Farklı sıcaklıklarda viĢne tomurcuk ve çiçeklerinin ölüm oranları Sıcaklık (oC) Ölüm Oranı (%) 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 0◦c 0 e 0 d 0 f 0 f 0 h 0 f -1◦c 0 e 0 d 0 f 0 f 0 h 0 f -2◦c 0 e 0 d 0 f 0 f 0 h 0 f -3◦c 0 e 0 d 0 f 0 f 0 h 0 f -4◦c 0 e 0 d 0 f 0 f 1 g 0 f -5◦c 0 e 0 d 0 f 0 f 2 f 9 e -6◦c 0 e 0 d 0 f 1 e 3 e 13 e -7◦c 0 e 0 d 0 f 7 d 8 d 16 d e -8◦c 0 e 0 c d 2 e 15 c 10 d 24 c d -9◦c 0 e 0 c d 2 e 34 b c 34 c 31 c -10◦c 1 d e 0 c d 3 e 50 b 67 b 52 b -11◦c 1 d e 1 c d 16 d 51 b 90 a b 93 a b -12◦c 1 d e 2 c 18 b c d 65 a b 100 a 100 a -13◦c 3 d 18 b 18 b c d 92 a 100 a 100 a -14◦c 8 c 77 a b 31 b c 100 a 100 a 100 a -15◦c 49 b 77 a b 36 b c 100 a 100 a 100 a -16◦c 97 a b 100 a 87 b 100 a 100 a 100 a -17◦c 99 a b 100 a 95 a b 100 a 100 a 100 a -18◦c 100 a 100 a 100 a 100 a 100 a 100 a -19◦c 100 a 100 a 100 a 100 a 100 a 100 a -20◦c 100 a 100 a 100 a 100 a 100 a 100 a -21◦c 100 a 100 a 100 a 100 a 100 a 100 a -22◦c 100 a 100 a 100 a 100 a 100 a 100 a 0% 5% 10% 15% 20% 25% 30% 35% 40% 45% 50% 55% 60% 65% 70% 75% 80% 85% 90% 95% 100% 0 -1 -2 -3 -4 -5 -6 -7 -8 -9 -10-11-12-13-14-15-16-17-18-19-20-21-22 Ö lüm O ra nı (%) Sıcaklık ( C) 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018
Bütün örnek alma dönemlerinde sıcaklık derecesinin düĢmesiyle birlikte örneklerde ölüm oranları artmıĢ olup, bu beklenen bir sonuçtur. Konu hakkında yapılan birçok çalıĢmada da benzer sonuçlar alınmıĢtır (Andrews ve Proebsting Jr, 1987; Burak, 1989; Demirel, 1997; Bartolozzi ve Fontanazza, 1999; Bartolini ve ark., 2001; Ertürk ve Güleryüz, 2007; Erdem ve ark., 2015).
ġeftalilerde yapılan dona dayanıklılık çalıĢmasında, tüm çeĢitlerin en yüksek dayanıklılığı ocak ve Ģubat aylarında, en düĢük dayanıklılığı mart ayında gösterdiği belirlenmiĢtir. Triogem ve Golden Jubilee çeĢitlerinin tomurcukları -21.1°C’de suni don testine tabi tutulmuĢtur. Ocak ayında sırasıyla % 58.3 ve % 55.1 oranlarında canlılık gösterirken, Mart ayında bu oran her iki çeĢitte de % 0.6’ya düĢmüĢtür. Ayrıca dayanıklılığın çeĢitlere göre değiĢtiği ifade edilmiĢtir (Küden ve ark., 1998).
KıĢ mevsiminde dona dayanıklılığı yüksek olan tomurcuklar ilkbahar mevsimine doğru bu dayanıklılığını kaybetmeye baĢlamıĢtır. Bu da havaların ısınmasıyla birlikte tomurcuklara su yürümesi ve uyanmaya baĢlamasının tabii bir sonucudur.
Güvenli bir ürün için çiçek tomurcuklarının en az % 50’sinin sağlıklı kalması gerektiğini kabul edildiğinden % 50’den fazla zarar meydana gelen sıcaklıklar 29.01 tarihinde -16°C, 19.02 tarihinde -14°C, 05.03 tarihinde -16°C, 19.03, 04.04 ve 20.04 tarihlerinde -10°C olarak tespit edilmiĢtir. Konu hakkında ılıman iklim meyve türlerinde daha önce yapılan çalıĢmalarda da dona mukavemetin kıĢ ortasına kadar arttığı ve en yüksek seviyeye ulaĢtığı, daha sonra azalmaya baĢladığı tespit edilmiĢtir (Zavarzin 1963; Burak 1989; Demirel 1997; AslantaĢ 1999).
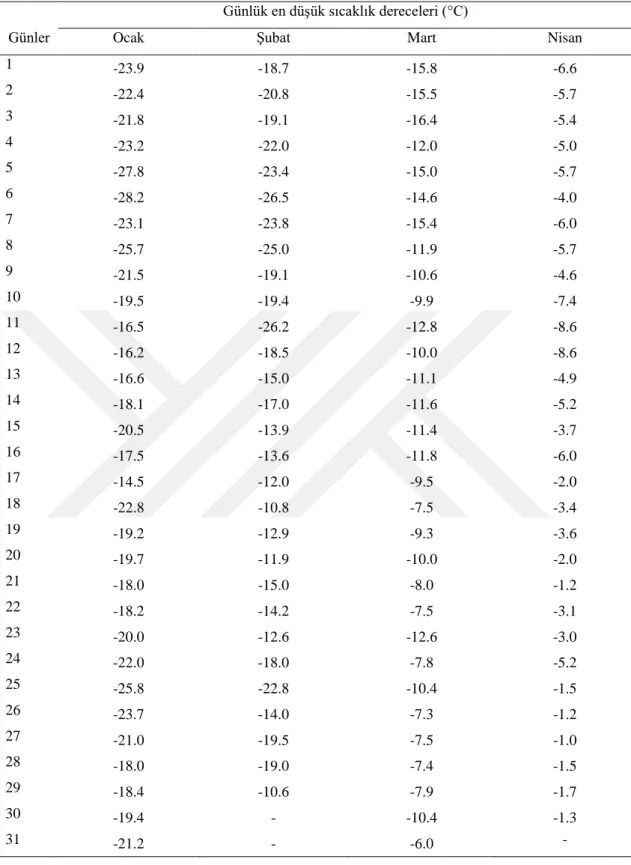
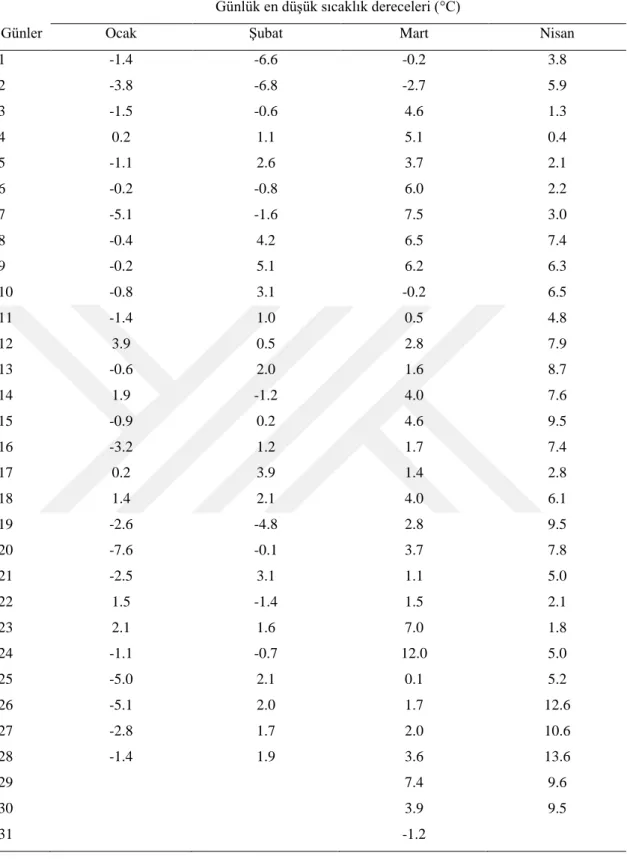
Konya il merkezinde uzun yıllar verilerine göre 1 Ocak -30 Nisan tarihleri arasında en düĢük sıcaklık değerleri Çizelge 4.1.’de verilmiĢtir. Buna göre Konya il merkezinde viĢne çiçek tomurcuklarının Ocak ayında dondan zarar görme ihtimali bulunmaktadır. Nitekim yaptığımız çalıĢmada 29.01 tarihinde alınan örneklerde -15°C’den itibaren % 50’den fazla zararlanma meydana geldiği tespit edilmiĢtir.
Uzun yıllar verilerine göre de Konya il merkezinde Ocak ayının her günü sıcaklığın -15°C’nin altına düĢme ihtimali vardır (Çizelge 4.2. ve Çizelge 4.3.). Aynı Ģekilde ġubat ayında da tomurcukların zarar görme ihtimali vardır. 19.02 tarihinde alınan örneklerde -14°C’den sonra % 77’den fazla ölüm tespit edilmiĢtir. Uzun yıllık verilere göre ġubat ayının 20’sine kadar sürede 14 gün sıcaklık -14°C’den düĢük gerçekleĢmiĢtir. 05.03 tarihinde alınan tomurcuklarda -8°C’den itibaren ölümler baĢlamıĢtır. Ancak bu ölüm oranları -17°C’ye kadar % 50’nin altında kalmıĢ, bu derecede ise % 87 olmuĢtur. Konya il merkezinde 20 ġubat – 6 Mart tarihleri arasında
sıcaklık 4 gün -17°C’den aĢağı düĢmüĢtür. Bu itibarla belirtilen dönemde viĢne tomurcuklarında zarar görme ihtimali azalmıĢ olmakla birlikte devam etmektedir. 20 Mart tarihinde alınan örneklerde -10°C’nin altında % 50’den fazla zarar meydana gelmiĢtir. Meteorolojik verilere göre bu dönemde de viĢne çiçek tomurcuklarının zarar görme ihtimali mevcuttur. Zira belirtilen dönemde 8 gün -10°C’nin altında sıcaklıklar ölçülmüĢtür. 5 Nisan tarihinde alınan örneklerde de -10°C’nin altında zarar oranı % 50’nin üzerine çıkmıĢtır. Bu dönemde havalar ısınmaya devam ettiğinden -10°C’nin altında gün sayısı 3’e düĢmüĢ, dolayısıyla tomurcukların zarar görme ihtimali azalmıĢtır. 20 nisan tarihinde alınan örneklerde de -10°C’den itibaren önemli zararlar meydana gelmiĢtir. Uzun yıllar verilerine göre Konya il merkezinde 5-20 nisan tarihleri arasında sıcaklık -10°C’nin altına hiç düĢmemiĢtir. Buna göre bu tarihler arasında viĢne tomurcukları ve çiçeklerinin zarar görme tehlikesi bulunmamaktadır.
Çizelge 4.2. Konya il merkezinde Ocak-Mayıs ayları arasında uzun yıllar günlük en düĢük sıcaklıklar Günlük en düĢük sıcaklık dereceleri (°C)
Günler Ocak ġubat Mart Nisan
1 -23.9 -18.7 -15.8 -6.6 2 -22.4 -20.8 -15.5 -5.7 3 -21.8 -19.1 -16.4 -5.4 4 -23.2 -22.0 -12.0 -5.0 5 -27.8 -23.4 -15.0 -5.7 6 -28.2 -26.5 -14.6 -4.0 7 -23.1 -23.8 -15.4 -6.0 8 -25.7 -25.0 -11.9 -5.7 9 -21.5 -19.1 -10.6 -4.6 10 -19.5 -19.4 -9.9 -7.4 11 -16.5 -26.2 -12.8 -8.6 12 -16.2 -18.5 -10.0 -8.6 13 -16.6 -15.0 -11.1 -4.9 14 -18.1 -17.0 -11.6 -5.2 15 -20.5 -13.9 -11.4 -3.7 16 -17.5 -13.6 -11.8 -6.0 17 -14.5 -12.0 -9.5 -2.0 18 -22.8 -10.8 -7.5 -3.4 19 -19.2 -12.9 -9.3 -3.6 20 -19.7 -11.9 -10.0 -2.0 21 -18.0 -15.0 -8.0 -1.2 22 -18.2 -14.2 -7.5 -3.1 23 -20.0 -12.6 -12.6 -3.0 24 -22.0 -18.0 -7.8 -5.2 25 -25.8 -22.8 -10.4 -1.5 26 -23.7 -14.0 -7.3 -1.2 27 -21.0 -19.5 -7.5 -1.0 28 -18.0 -19.0 -7.4 -1.5 29 -18.4 -10.6 -7.9 -1.7 30 -19.4 - -10.4 -1.3 31 -21.2 - -6.0 -
Çizelge 4.3. Konya il merkezinde 2018’in Ocak-Mayıs ayları arasında günlük en düĢük sıcaklıklar Günlük en düĢük sıcaklık dereceleri (°C)
Günler Ocak ġubat Mart Nisan
1 -1.4 -6.6 -0.2 3.8 2 -3.8 -6.8 -2.7 5.9 3 -1.5 -0.6 4.6 1.3 4 0.2 1.1 5.1 0.4 5 -1.1 2.6 3.7 2.1 6 -0.2 -0.8 6.0 2.2 7 -5.1 -1.6 7.5 3.0 8 -0.4 4.2 6.5 7.4 9 -0.2 5.1 6.2 6.3 10 -0.8 3.1 -0.2 6.5 11 -1.4 1.0 0.5 4.8 12 3.9 0.5 2.8 7.9 13 -0.6 2.0 1.6 8.7 14 1.9 -1.2 4.0 7.6 15 -0.9 0.2 4.6 9.5 16 -3.2 1.2 1.7 7.4 17 0.2 3.9 1.4 2.8 18 1.4 2.1 4.0 6.1 19 -2.6 -4.8 2.8 9.5 20 -7.6 -0.1 3.7 7.8 21 -2.5 3.1 1.1 5.0 22 1.5 -1.4 1.5 2.1 23 2.1 1.6 7.0 1.8 24 -1.1 -0.7 12.0 5.0 25 -5.0 2.1 0.1 5.2 26 -5.1 2.0 1.7 12.6 27 -2.8 1.7 2.0 10.6 28 -1.4 1.9 3.6 13.6 29 7.4 9.6 30 3.9 9.5 31 -1.2
4.2. Örneklerin Toplam, Bağlı, Serbest Su ve Kuru Madde Miktarı
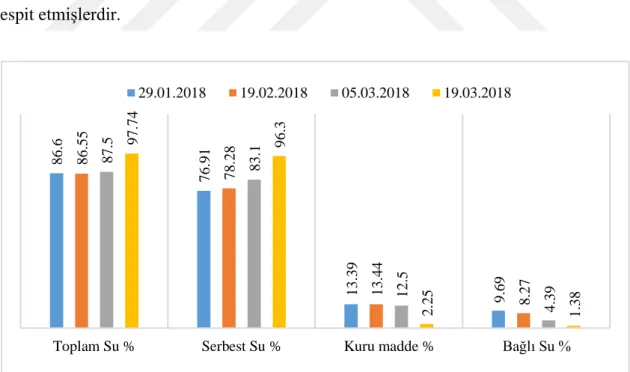
Farklı dönemlerde viĢne tomurcukları ve çiçeklerinin su ve kuru madde içeriklerine ait sonuçlar Çizelge 4.4.’de verilmiĢtir. Farklı dönemlerde örneklerin su ve kuru madde içerikleri arasındaki farklar istatistiki olarak önemli bulunmuĢtur. 29.01 tarihinde tomurcuklarda toplam su içeriği % 86.60 iken, 19.02 tarihinde % 86.55, 05.03’de % 87.50 ve 19.03’de % 97.74’a yükselmiĢtir. Buna göre kıĢ mevsiminden çıkıĢla birlikte tomurcuklarda su içeriğinin arttığı bariz bir Ģekilde görülmektedir. Bu da havaların ısınması, buna bağlı olarak tomurcukların uyanması ve büyümesinin sonucu olup beklenen bir durumdur.
Bitki dokularındaki su durumu, genel olarak don zararı potansiyelini gösterir. Don zararı, donmaya iliĢkin bitki hücrelerinin dehidrasyonun sonucunda gerçekleĢmektedir (Xin ve Browse, 2000) ve su içeriği yüksek olan bitki dokuları donma sonucu daha fazla zarar görmektedir (Hao ve ark., 2009).
Tomurcuklarda kıĢtan çıkıĢla birlikte bağlı su miktarında hızlı bir azalma, buna karĢılık serbest su miktarında ise hızlı bir yükselme tespit edilmiĢtir. 29.01 tarihinde 9.69 % olan bağlı su 19.02 tarihinde 8.27, 05.03 tarihinde 4.39 ve 19.03 tarihinde 1.38 % düĢmüĢtür. Buna karĢılık 29.01 tarihinde 76.91 % olan serbest su 19.02 tarihinde 78.28, 05.03 tarihinde 83.1 ve 19.03 tarihinde 96.3 % yükselmiĢtir (Çizelge 4.4. ve ġekil 4.2.). Benzer Ģekilde Ertürk ve Güleryüz (2007) tarafından kayısıda yapılan çalıĢmada da kıĢ mevsiminden ilkbahara doğru tomurcukların su içeriklerinin arttığı ve su içeriği ile dona dayanım arasında negatif korelasyon olduğu tespit edilmiĢtir. Dokularda su içeriğinin artması dokuların dona dayanıklılığını azaltmakta ve zararlanma tehkilesini artırmaktadır. Dokulardaki su miktarının yüksekliği piĢkinleĢmeyi engellemekte, bu durum dayanıklılığı azaltmaktadır. Soğuğa alıĢma dönemi hem dokularda suyun azalmasını, hem de osmotik olarak aktif olmayan protein ve niĢastanın birikimini teĢvik etmektedir. Bu itibarla bitkilerin dokularında daha az miktardaki su, adaptasyon için oldukça önemli bir avantaj sağlayabilir. Bu sayede fiziksel olarak hücreler arası boĢluklarda daha az geniĢleme ve bozulma meydana gelebilir (Levitt, 1980; Stancu, 1991).
Çizelge 4.4. ViĢne tomurcukların toplam, bağlı, serbest su ve kuru madde miktarları (%) Dönem Tarihi 29.01.2018 19.02.2018 05.03.2018 19.03.2018 Serbest Su % 76.91 c 78.28 c 83.10 b 96.30 a Bağlı Su % 9.69 a 8.27 a 4.39 b 1.38 c Toplam Su % 86.60 b 86.55 b 87.50 b 97.74 a Kuru madde % 13.39 a 13.44 a 12.50 a 2.25 b
Farklı tarihlerde alınan tomurcuklarda kuru madde içerikleri ise 29.01.2018 ile 05.03.2018 arasında hemen hemen aynı bulunurken, 19.03.2018 tarihinde hızlı bir azalma tespit edilmiĢtir. KıĢı dinlenme döneminde geçiren çiçek tomurcukları ilkbaharda havaların ısınmasıyla birlikte uyanmaya baĢlar. Bu uyanmada havaların ısınmasıyla birlikte toprakta ve bitki bünyesinde suyun hareketlenmesinin etkisi vardır. Bitkide suyun hareketiyle birlikte depolanmıĢ besin maddeleri tomurcuklara doğru hızla hareket eder. Tomurcuklarda su ve kuru madde miktarları birbirine zıt yönde değiĢim göstermiĢtir. Suyun artıĢıyla birlikte dona hassasiyeti artan tomurcuklar kuru madde artıĢıyla birlikte dayanıklılık kazanır. Benzer Ģekilde AslantaĢ (1999) da badem tomurcuklarında kuru madde miktarı ile dana dayanıklılık arasında pozitif korelasyon tespit etmiĢlerdir. 86. 6 76. 91 13 .39 9. 69 86 .55 78. 28 13. 44 8. 27 87. 5 83. 1 12. 5 4. 39 97. 74 96. 3 2. 25 1. 38
Toplam Su % Serbest Su % Kuru madde % Bağlı Su %
29.01.2018 19.02.2018 05.03.2018 19.03.2018
4.3. Bitki Besin Maddesi Ġçerikleri
Farklı tarihlerde alınan viĢne tomurcuk ve çiçeklerinde besin elementi analiz sonuçları (Çizelge 4.5., ġekil 4.3., 4.4., 4.5., 4.6. ve 4.7.)’de verilmiĢtir. Örnek alma dönemleri arasında besin maddesi miktarları arasındaki farklar istatistiki olarak önemli bulunmuĢtur. Buna göre örneklerde N içeriği 29 Ocak ile 20 Nisan tarihleri arasında sürekli azalma göstermiĢtir. Leece ve Gilmour (1974) azotun ilkbaharda hızla azalmasını geliĢmenin sonucu olarak su içeriğinin artmasıyla açıklanabileceğini bildirmiĢtir. Benzer Ģekilde Uçgun (2012) da elmada ilkbaharda bitki bünyesinde azot konsantrasyonunun hızla düĢtüğünü belirlemiĢtir. P, K, Mn, Zn ve S miktarları 29.01 tarihinden 19.03 tarihine kadar artmıĢ, bu tarihten sonra azalmaya baĢlamıĢtır. Azotta olduğu gibi baĢlangıçta yüksek olan fosforun hızla azalması kuru madde artıĢının topraktan alınan fosfordan daha fazla olması ve fosforun diğer organlara dağılmasından kaynaklanmıĢ olabilir (Leece ve Gilmour, 1974; Kovancı ve Köseoğlu, 1978; Uçgun, 2012). Mg ve Fe miktarları 19.03 tarihine kadar düzenli bir Ģekilde artmıĢ, bu tarihten sonra sabit kalmıĢtır. Mg ve K ksilem ve floemde hareketli elementler olup katyonlar arasında rekabet vardır (Neilsen ve Neilsen, 2003). Özellikle bu etki Mg, K ve Ca arasında belirgindir (Stiles, 1994). Ca ve Cu miktarları ise 05.03 tarihine kadar yükselmiĢ, daha sonra düĢmeye baĢlamıĢtır. B içeriği ise sürekli artıĢ göstermiĢtir. 20.04 tarihinde de 178 mg/kg’a yükselmiĢtir. Zn miktarı 04.04 tarihine kadar düzenli olarak artıĢ göstermiĢ, daha sonra düĢmüĢtür. S içeriği ise farklı tarihlerde dalgalanma göstermiĢtir. 20.04 tarihinde artmıĢ, bu tarihten sonra azalmaya baĢlamıĢtır.
Çizelge 4.5. Farklı tarihlerde viĢne çiçek tomurcuğu ve çiçeklerinde besin maddesi miktarları Besin Elementleri 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 N (%) 2.75 a 2.76 a 2.66 ab 2.45 ab 2.50 ab 2.31 b Ca (mg/kg) 11230 b 11320 b 13450 a 12340 ab 12300 ab 11800 b K (mg/kg) 20134 c 22243 b 22350 b 24350 a 21220 bc 21560 bc P (mg/kg) 2540 c 3012 b 2979 b 3355 a 2544 c 2715.5 c S (mg/kg) 2012 a 2033 a 2045 a 2320 b 2243 b 2210 b Mg (mg/kg) 1890 b 2034 b 2120 ab 2340 a 2310 a 2300 a Fe (mg/kg) 134 c 145 bc 167 a b 176 a 175 a 178 a Mn (mg/kg) 34.45 c 35.76 bc 36.74 bc 45.65 a 40.21 b 38.77 bc Zn (mg/kg) 28.76 b 28.44 b 33.23 ab 34.35 a 36.54 a 32.24 ab Cu (mg/kg) 44.56 d 47.68 cd 55.34 a 53.44 ab 52.23 bc 48.76 cd B (mg/kg) 12.33 b 13.05 ab 12.24 b 13.43 ab 14.32 ab 15.46 a
2.75 2.76 2.66 2.45 2.5 2.31 2.2 2.3 2.4 2.5 2.6 2.7 2.8 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 A zo t M ikt ar i (% )
Örnek alma tarihleri N,%
ġekil 4.3. Örneklerde N içeriğinin dönemlere göre değiĢimi
11230 11320 13450 12340 12300 11800 20134 23243 22340 24350 21220 21560 10000 12000 14000 16000 18000 20000 22000 24000 26000 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g /k g )
Örnek alma tarihleri
Ca, mg/kg K, mg/kg
ġekil 4.4. Örneklerde Ca ve K içeriğinin döneme göre değiĢimi
1800 2000 2200 2400 2600 2800 3000 3200 3400 3600 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g /k g )
Örnek alma tarihleri
P, mg/kg Mg, mg/kg S, mg/kg
134 145 167 176 145 178 120 130 140 150 160 170 180 190 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g /k g )
Örnek alma tarihleri Fe, mg/kg
ġekil 4.6. Örneklerde demir içeriğinin dönemlere göre değiĢimi
34.45 38.76 33.24 45.65 40.21 38.77 28.76 25.44 33.23 34.35 36.54 32.24 44.56 47.68 55.43 53.44 50.23 48.76 12.33 13.25 11.24 13.43 14.32 15.46 10 15 20 25 30 35 40 45 50 55 60 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g /k g )
Örnek alma tarihleri
Mn, mg/kg Zn, mg/kg Cu, mg/kg B, mg/kg
ġekil 4.7. Örneklerde Mn, Zn, Cu ve B içeriğinin dönemlere göre değiĢimi
Donma/çözünme stresi sonucu hücrelerde iyon kaybı belirlenmiĢ ve bu durum membran geçirgenliğinin kaybolmasına neden olmuĢtur. Don zararı olan hücrelerden kaybolan esas katyon potasyumdur. Hücre plasma membranı vasıtasıyla potasyum taĢınımı H+-ATPaz aktivitesi ile olduğundan, donma/erime stresi süresince H+-ATPaz fonksiyonunda değiĢim olduğu belirtilmektedir (AslantaĢ ve ark., 2010). DüĢük sıcaklığa maruz kalma ile dokularda kalsiyum miktarı geçici olarak artar (Puhakainen, 2004). Bu süreçte apoplasttan sitoplâzma sıvısına kalsiyum geçiĢi meydana gelmektedir. Plasma membranlarında bulunan kalsiyum kanalları sıcaklık düĢüĢüne karĢı sensör görevi yaparlar (Smallwood ve Bowles, 2002). DüĢük sıcaklığa maruz kalan bitkilerde hücre membran akıĢkanlığında azalma meydana gelir. Bu durum soğuk uyartımlı
genlerin çalıĢması için bir sinyal özelliği taĢıdığı yapılan çalıĢmalarla gösterilmiĢtir. Yigit ve Güleryüz (1995) Kütahya viĢne çeĢidinde potasyum gübrelemesinin soğuğa dayanım üzerindeki etkilerini incelediği çalıĢmada, potasyumun bitkilerdeki dayanıklılığı karbonhidrat metabolizmasını etkileyerek artırdığı, uygun dozlarda uygulanan potasyumlu gübrelerin bitki dokusunda Ģeker birikimini ve osmotik basıncı artırmak suretiyle donma noktasını daha düĢük sıcaklıklara çektiğini belirlemiĢlerdir.
4.4. Toplam Karbonhidrat Miktarı
Farklı tarihlerde alınan örneklerin toplam karbonhidrat içerikleri Çizelge 4.6. ve ġekil 4.8.’de verilmiĢtir. Karbonhidrat içeriği bakımından tarihler arasında önemli farklar tespit edilmiĢtir. Buna göre karbonhidrat içeriği 29.01 tarihinden 04.04 tarihine kadar düzenli olarak azalmıĢ, 20.04 tarihinde ise tekrar yükselmeye baĢlamıĢtır. Bu durum tomurcukların bünyesindeki depo karbonhidratlarının tomurcukların açılması aĢamasında harcandığını göstermektedir. 20.04 tarihinde toplam karbonhidrat içeriğinin artmaya baĢlaması ise yeni meydana gelen yapraklardan fotosentez sonucu karbonhidrat üretimi ile açıklanabilir.
Çizelge 4.6. Farklı tarihlerde viĢne örneklerin karbonhidrat, protein ve prolin miktarları
Dönem Tarihi 29/01 19/02 05/03 19/03 04/04 20/04 Karbonhidrat (mg/g) 4.232 a 4.063 b 3.933 c 3.542 e 3.516 e 3.768 d Protein (mg/g) 1.423 a 1.409 b 1.346 c 1.315 d 1.236 e 1.215 e Prolin (mg/g) 2.573 2.526 2.550 2.566 2.589 2.590 4.2329 4.06324 3.93359 3.5421 3.51679 3.76831 3.4 3.5 3.6 3.7 3.8 3.9 4 4.1 4.2 4.3 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g /g )
Örnek alma tarihleri karbonhidrat miktari
Karbonhidratlar bitkilerde soğuğa dayanımda temel kriyoprotektanlardır. Karbonhidrat seviyesinin düĢük sıcaklık süresince 10 kata kadar bir artıĢ gösterdiği ve bu birikimin dona dayanımda etkili olduğu bildirilmiĢtir (Guy, 1990). Bu etkinin muhtemelen karbonhidratların ölümcül hücre içi donma noktasını daha düĢük derecelere çekmesiyle meydana geldiği düĢünülmektedir. Bu maddeler ayrıca sitoplâzmadan dehidrasyonu önlemek için osmoprotektan olarak görev alarak donmanın tolere edilmesi için hücreleri hazırlar. Ayrıca poteinlerin etrafında bulunan suyun muhafaza edilmesine de yardımcı olarak dayanıma katkı sağlarlar (Smirnoff, 1995).
Farklı meyve türlerinde dinlenme döneminde karbonhidratların dokulardaki birikimi ve soğuğa dayanım arasındaki iliĢkiler incelenmiĢtir. Bu kapsamda, Burak (1989) Ģeftali, Bolat (1993) ve Demirel (1997) kayısı, Palonen (1999) ahududu, Ameglio (2001) cevizde karbonhidratların dönemsel değiĢimi ile dona dayanım arasında önemli iliĢkiler olduğunu belirlemiĢlerdir.
4.5. Protein Miktarı
Farklı tarihlerde alınan örneklerin toplam protein içerikleri Çizelge 4.6. ve ġekil 4.9.’da verilmiĢtir. Protein içerikleri birbirini takip eden örnek alma dönemlerinde sürekli azalma göstermiĢtir. 29.01 tarihinde 1.423 olan miktar 05.03 tarihinde 1.346, 20.04 tarihinde ise 1.215’e düĢmüĢtür. Bu düĢüĢ de toplam karbonhidratta olduğu gibi tomurcukların açılma döneminde proteinlerin besin kaynağı olarak kullanılmasının sonucudur. Bitkilerin soğuğa dayanıklılık fizyolojisinde proteinler önemli bir rol oynamaktadır (Ertürk ve Güleryüz, 2007). Birçok çalıĢma düĢük sıcaklığa tepki olarak yeni polipeptidlerin sentezlendiğini göstermiĢtir. Son yıllarda dona dayanım ile proteinler arasında iliĢki olduğuna dair çalıĢmalar yoğunluk kazanmaktadır (AslantaĢ ve ark., 2010). Burak (1989) çiçek tomurcuklarında protein oranlarının yüksek olmasının dona dayanıklılığı olumlu etkilediğini belirtmiĢtir.
Elde ettiğimiz sonuçlar Litvinova (1974), Tamássy ve Zayan (1981), Burak (1989)’ın sonuçlarıyla benzerlik göstermektedir. Proteinlerin membran yapısına katılması, suyun tutulmasındaki hidrofilik özellikleri, protoplazma akıĢkanlığı ve elastikiyetini yönlendirmesi gibi önemli görevleri vardır. Bu nedenle proteinlerin soğuğa mukavemete katkısı olduğu bilinmektedir. Çözülebilir proteinlerin değiĢimi sonucu buz kristallerinin hücre içinde oluĢumu teĢvik edilmektedir. Çözünebilir protein içeriği ile dona dayanıklılık arasındaki yakın iliĢki, protein sentezi için gerekli olan
tRNA, mRNA ve polizomların miktarındaki artıĢla alakalıdır (Levitt, 1980). Proteinlerin dokuların piĢkinleĢmesinde olumlu rol oynadığı kayısı, Ģeftali ve zeytinde yapılan çalıĢmalarla tespit edilmiĢtir (Ashworth, 1983; Bartolozzi ve Fontanazza, 1999; Marquat ve ark., 1999; Bartolini ve ark., 2001).
Bitkilerin dona dayanıklılık fizyolojisinde aminoasitler ve proteinlerin rolü bulunmaktadır. Genellikle bitkilerin düĢük sıcaklığa maruz kalması sırasında bünyelerindeki çözünebilir protein miktarının nispi olarak arttığı, dona dayanıklı tür ve çeĢitlerde bu oranın yüksek, dayanıksız çeĢitlerde ise düĢük olduğu ifade edilmektedir (Ertürk ve Güleryüz, 2007). Soğuk aklimasyonu süresince biriken özel proteinlerden bazıları antifriz protein özelliği göstermektedir (Pearce, 2001). Antifiriz proteinler (AFPs) buz kristalinin Ģekli ve oluĢumunu değiĢtirme özelliğine sahiptir ve buzun yeniden kristalizasyonunu engeller. Bunun sonucunda hücrelerin don zararından korunmasına katkı sağlarlar. Arpa, buğday, çavdar ve kanolada soğuk aklimasyonu süresince AFP birikimi olduğu gözlemlenmiĢtir (Scebba ve ark., 1998; Baek ve ark., 2000). Deterjan özelliğe sahip olan bu proteinler suyu çekmeyen ve makromoleküllerin bir araya gelmesini önleyici özelliğe sahiptir (Pearce, 1999; Smallwood ve Bowles, 2002). Soğuk uyartımlı proteinlerin çoğunun sentezi absisik aside karĢı tepki olarak değerlendirilmektedir (Tseng ve Li, 1990).
1.42308 1.40959 1.34611 1.31596 1.23646 1.21511 1.2 1.25 1.3 1.35 1.4 1.45 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g /g )
Örnek alma tarihleri Protein miktari (mg/g yaş made)
4.6. Prolin Miktarı
Farklı tarihlerde alınan örneklerde toplam prolin miktarı Çizelge 4.6. ve ġekil 4.10’da verilmiĢtir. Prolin içeriği arasındaki farklar istatistiki olarak önemli bulunmamıĢtır. Bir amino asit olan prolin birçok çevresel strese tepki olarak geliĢmiĢ bitkilerde sentezlenip biriktirilmektedir (Yeo ve Flowers, 1989; Ashraf ve Wu, 1994; Ali ve ark., 1999; Öztürk ve Demir, 2002; Hsu ve ark., 2003; Kishor ve ark., 2005). Soğuğa alıĢma süresi boyunca stres gören bitkilerde prolin miktarı büyük bir ölçüde artıĢ gösterir (Hare ve Cress, 1997). Soğuğa uyum sağlamıĢ turunçgil bitkilerinde aklimize olmamıĢ bitkilere göre 3 ile 6 kat daha fazla prolin biriktirildiği belirlenmiĢtir (Yelenosky, 1979).Xin (1998)’ un yaptığı bir çalıĢmada soğuk aklimizasyonu boyunca
Arabidopsis bitkilerinde 30 kat prolin artıĢı görülmüĢtür. Diğer stres faktörleri de prolin
miktarını oldukça değiĢtirmektedir. Pırlak ve EĢitken (2004),çilek çeĢitleri ile yaptıkları bir çalıĢmada tuz stresi altındaki bitkilerde prolin miktarının büyük ölçüde arttığını bildirmiĢlerdir. Burada farklı sonuçların elde edilmesi türlerin genetik yapılarının farklı olmasından kaynaklanmıĢ olabilir.
2.57358 2.52611 2.55099 2.56687 2.58906 2.59097 2.52 2.53 2.54 2.55 2.56 2.57 2.58 2.59 2.6 29.01.2018 19.02.2018 05.03.2018 19.03.2018 04.04.2018 20.04.2018 (m g/ g )
Örnek alma tarihleri Prolin miktari (mg/g yaş made)
5. SONUÇLAR VE ÖNERĠLER
Ölümcül sıcaklıklar ve soğuğa dayanımdaki farklılıklar farklı geliĢim evrelerinde gözlenmiĢtir. Gözlenen hasar çiçek tomurcuğu geliĢim evrelerine yüksek oranda bağlıdır. Tomurcukların düĢük sıcaklıklara karĢı artan bir hassasiyeti olduğu gözlenmiĢtir. Çiçek toleransı erken çiçek tomurcuğu evresi ve çiçeklenme boyunca sıcaklık dalgalanmasından etkilenir, sıcak Ģartlar çiçek hassasiyetini artırırken, düĢük sıcaklık bu Ģartı azaltır. Maksimum dayanıklılığın belirlenmesi genetik olarak elde edilebilmesine rağmen, iklimsel etkinin bir sonucu olarak daha fazla yıllar içerisinde süren gözlemlere ihtiyaç vardır. Dinlenme halindeki viĢne çiçek tomurcukları için öldürücü sıcaklık -16°C iken, açılmamıĢ çiçek tomurcuğu evresinde -11°C’ye doğru hızlı bir Ģekilde yükselmiĢtir.
Konya il merkezinde ekonomik viĢne yetiĢtiriciliğinde özellikle dinlenme döneminde meydana gelebilecek extrem düĢük sıcaklıklar bazı yıllarda viĢne çiçek tomurcuklarına zarar verebilir. Bu itibarla kapama bahçeler kurulurken mümkün olduğu kadar soğuk havanın biriktiği çukur alanlardan kaçınmak, mümkün olduğu kadar korunaklı alanların tercih edilmesi önerilebilir.