T.C.
EGE ÜNİVERSİTESİ TIP FAKÜLTESİ
İNSAN PROSTAT KANSER HÜCRE HATTINDA (Du-145 Hücre Hattı) GM (SINGLE NANA GANGLIOSIDE) ÜZERİNDE KDN
(2-KETO-3-DEOXY-D-GLYCERO-D-GALACTONONONIC ACID)’NİN GÖSTERİLMESİ
Dr. Kenan DEMİR
HİSTOLOJİ VE EMBRİYOLOJİ ANABİLİM DALI UZMANLIK TEZİ
Tez Yöneticisi: Prof. Dr. Hüseyin AKTUĞ
i ÖNSÖZ
Uzmanlık eğitimim süresince engin bilgi ve tecrübelerinden yararlandığım, yetişmemde büyük katkıları olan, tez çalışmalarım boyunca vermiş olduğu sonsuz katkı nedeniyle Anabilim Dalı başkanımız Sayın Prof. Dr. Ayşegül UYSAL’a teşekkür ederim.
Eğitim sürem boyunca haftanın 7 günü, günün 24 saati bana her konuda yardımcı olan ve destek veren, tez çalışmalarım esnasında bana her türlü yardımı fazlasıyla sağlayan çok değerli hocam Sayın Prof. Dr. Hüseyin AKTUĞ’a özellikle teşekkür ederim. Dünya sizin ve Sayın Prof. Dr. Altuğ YAVAŞOĞLU gibi kalbi olan insanlar sayesinde hala dönüyor.
Tezimde projesini, fikirlerini ve malzemelerini benimle paylaşan, her zaman beni her konuda destekleyen Sayın Prof. Dr. Gülperi ÖKTEM’e teşekkür ederim.
Benimle sadece hoca asistan ilişkisi içerisinde kalmayıp, aynı zamanda arkadaş olan, dost olan, dertlerimi dinleyen, hayatımı paylaşan değerli hocalarım Sayın Prof. Dr. Hüseyin AKTUĞ, Sayın Prof. Dr. Altuğ YAVAŞOĞLU ve uzman abilerim Sayın Uz. Dr. Fatih OLTULU ve Sayın Uz. Dr. Türker ÇAVUŞOĞLU’na teşekkür ederim.
Yine her zaman yanımda olan değerli hocalarım Sayın Prof. Dr. Meral BAKA’ya, Sayın Prof. Dr. Yiğit UYANIKGİL’e, Sayın Doç. Dr. Özlem YILMAZ’a teşekkür ederim.
Birlikte olmaktan büyük mutluluk duyduğum çok sevdiğim çalışma arkadaşlarım Sayın Dr. Aylin GÖKHAN’a, Sayın Araş. Gör. Gürkan YİĞİTTÜRK’e, Sayın Araş. Gör. Eda AÇIKGÖZ’e, Sayın Araş. Gör. Fahriye DÜZAĞAÇ’a, Sayın Araş. Gör. Ümmü GÜVEN’e, Sayın Araş. Gör. Hadi ROUHRAZİ’ye ve bölümümüzdeki tüm çalışma arkadaşlarıma teşekkürü borç bilirim.
Fourier Transform Infrared Spectroscopy Assay (FTIR) çalışmalarında değerli yardımlarını sunan (Ege Üniversitesi ARGEFAR) Sayın Dr. Günnur GÜLER’e ve Prof. Dr. Ercüment KARASULU’ya ayrıca teşekkür ederim.
Bu tez çalışmamı, yetişmemde büyük emeği olan başta rahmetli babama olmak üzere anneme, kardeşime ve eşime armağan ediyorum.
ii İÇİNDEKİLER ÖNSÖZ ... i TABLOLAR/ŞEKİLLER/GRAFİKLER LİSTESİ ... iv KISALTMA ve SEMBOLLER DİZİNİ ... vi ÖZET ... viii SUMMARY ... ix 1. BÖLÜM ... 10 1.1. GİRİŞ VE AMAÇ: ... 10 1.1.1. GCS Enzimi: ... 17 1.1.2. GD3 Sentaz Enzimi: ... 17 1.1.3. GCS İnhibitörleri: ... 18 1.1.4. GD3 Sentaz İnhibitörleri: ... 18 2. BÖLÜM ... 20 2.1. GEREÇ VE YÖNTEMLER: ... 20
2.1.1. Çalışmada Kullanılan Kimyasal Maddeler ve Cihazlar: ... 20
2.1.2. Çalışmada Kullanılan Hücre Hatlarının Temini ve Üretilmesi:... 21
2.2. ÇALIŞMADA KULLANILAN İLAÇLARIN VE KIMYASALLARIN HAZIRLANMASI: ... 22
2.2.1. Genz-123346‘nın stok solüsyonlarının hazırlanması: ... 22
2.2.2. Triptolide’in stok solüsyonun hazırlanması: ... 22
2.2.3. Tampon Çözeltisinin Hazırlanması: ... 22
2.2.4. Paraformaldehid’in Hazırlanması: ... 23
2.3. DENEYİN KURGULANMASI VE DENEY GRUPLARI: ... 23
2.4. ÇALIŞMADA KULLANILAN YÖNTEMLER: ... 24
2.4.1. Hücre Sayısı ve Canlılığının Belirlenmesi: ... 24
2.4.2. İmmunohistokimyasal Yöntem: ... 25
2.4.3. (Fourier Transform Infrared Spectroscopy Assay) FTIR Analizi: . 26 3. BÖLÜM ... 30
3.1. BULGULAR: ... 30
iii
3.1.2. İmmunohistokimya Boyama Sonuçları: ... 38
3.1.3. Fourier Transform Infrared Spectroscopy Assay (FTIR) Sonuçları: . ... 45 4. BÖLÜM ... 49 4.1. TARTIŞMA: ... 49 5. BÖLÜM ... 53 5.1. SONUÇLAR: ... 53 6. KAYNAKLAR ... 54 6.1. KAYNAKLAR ... 54
iv
TABLOLAR/ŞEKİLLER/GRAFİKLER LİSTESİ
Şekil 1. Prostat [2]. ... 10
Şekil 2. Prostat bezi [3]. ... 11
Şekil 3. Du-145 Prostat Kanser Hücre Hattı hücrelerinin kontrast mikroskop altındaki görüntüsü [6]. ... 13
Şekil 4. GM3’ün biyokimyasal yapısı [14]. ... 15
Şekil 5. GD3’ün biyokimyasal yapısı [15]. ... 15
Şekil 6. Seramidden Gangliozid sentezinin basamakları [20]. ... 18
Tablo 1. Çalışmada kullanılan kimyasal maddeler, üretici firma bilgileri ve kullanım amaçları. ... 20
Tablo 2. Çalışmada kullanılan cihazlar ve markaları. ... 21
Şekil 7. Muse™ Cell Analyzer cihazında % canlılık değerlerinin belirlenmesi [22].25 Şekil 8. Fourier Transform Infrared Spectroscopy Assay (FTIR) [23] ... 26
Şekil 9. Genz-123346’nın çeşitli dozlardaki 24. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı). ... 31
Şekil 10. Genz-123346’nın çeşitli dozlardaki 48. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı). ... 32
Şekil 11. Genz-123346’nın çeşitli dozlardaki 72. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı). ... 33
Şekil 12. GCS inhibitörü Genz-123346’nın Du-145 insan prostat kanseri hücre hattında zamana bağlı olarak hücre canlılığı (%) üzerindeki etkileri. ... 34
Şekil 13. GD3S inhibitörü Triptolide’in çeşitli dozlardaki 24. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı)... 35
Şekil 14. GD3S inhibitörü Triptolide’in çeşitli dozlardaki 48. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı)... 36
Şekil 15. GD3S inhibitörü Triptolide’in çeşitli dozlardaki 72. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı)... 37
Şekil 16. GD3S inhibitörü Triptolide’in Du-145 insan prostat kanseri hücre hattında zamana bağlı olarak hücre canlılığı (%) üzerindeki etkileri. ... 38
v
Şekil 17. GCS inhibitörü Genz-123346’nın 1000 µM dozu ile 24 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GM3 gangliozidinde meydana gelen değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X). ... 39 Şekil 18. GCS inhibitörü Genz-123346’nın 1000 µM dozu ile 24 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GD3 gangliozidinde meydana gelen değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X). ... 40 Şekil 19. GCS inhibitörü Genz-123346’nın 1000 µM dozu ile 24 saat muamele edilen Du-145 insan prostat kanseri hücre hattında KDN’de meydana gelen
değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X). ... 41 Şekil 20. GD3 inhibitörü Triptolide’in 60 µM dozu ile 72 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GM3 gangliozidinde meydana gelen
değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X). ... 42 Şekil 21. GD3 inhibitörü Triptolide’in 60 µM dozu ile 72 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GD3 gangliozidinde meydana gelen
değişimin ışık mikroskobu görüntüsü(Ölçek: 20X, 40X). ... 43 Şekil 22. GD3 inhibitörü Triptolide’in 60 µM dozu ile 72 saat muamele edilen Du-145 insan prostat kanseri hücre hattında KDN’de meydana gelen değişimin ışık mikroskobu görüntüsü(Ölçek: 20X, 40X)... 44 Şekil 23. Kontrol, Genz-123346 ve Triptolide ile muamele edilmiş Du-145 insan prostat kanseri hücre hatlarının FTIR spektrumu, 3100-2800 ve 1800-800 cm-1 spektrum bölgeleri. ... 46 Şekil 24. 3000-2800 ve 1800-800 cm-1 spektrum bölgeleri PCA skoruyla temel bileşenler analizi (Şekil 3.1.4.1’den elde edilmiştir). Çember içindeki değerler her temel bileşenin toplam varyasyon yüzdesini göstermektedir... 47 Şekil 25. FTIR spektrum farkı (treated-untreated) (Şekil 3.1.4.1’den elde edilmiştir). Du-145 hücre hatlarının Triptolide ve Genz-123346 uygulanmış FTIR sonuçları 3100-2800 cm-1 ve 1800-800 cm-1 aralığında gösterilmiştir. Spektrumlar baseline düzeltilmiş olup amid II bandına göre normalize edilmiştir. ... 48
vi
KISALTMA ve SEMBOLLER DİZİNİ
KISALTMALAR:
ATCC American Type Culture Collection, Amerikan Tip Kültür Koleksiyonu
ATR Attenuated Total Reflectance
Cer Ceramide, Seramid
Cisplatin Cis-diamminedichloroplatinum
DAPI 4', 6 Diamidino 2 Phenylindole
DMSO Dimetil Sülfoksid
EMT Epithelial Mesenchymal Transition, Epitelyal Mezenşimal Geçiş
FTIR Fourier Transform Infrared Spectroscopy Assay, Fourier Dönüşümlü Kızılötesi Spektrofotometre
Gal Galaktoz
GCS, GlcCerS Glucosylceramide Synthase, Glukozilseramid Sentaz
GD3 Disialosyllactosylceramide
Glc Glukoz
Glc-Cer Glucoceramide, Glukoseramid
GM3 Monosialodihexosyl Ganglioside
GM3 Senthase Lactosylceramide Alpha-2,3-Sialyltransferase HPTLC Yüksek Performans İnce Tabaka Kromatografisi
IC50 Hücrelerin %50’sini öldüren doz
IR Infrared, Kızılötesi
KDN Deaminoneuraminic Acid,
2-Keto-3-Deoxy-D-Glycero-D-Galacto-Nonanic Acid
Lac-Cer Lactosylceramide, Laktozilseramid
NB-DNJ N-butyl-deoxynojirimycin
Neu5Ac, NANA N-Acetylneuraminic Acid
vii
PCA Principal Component Analysis, Temel Bileşenler Analizi
PFA Paraformaldehit
ShRNA Short Hairpin RNA ya da Small Hairpin RNA
Sia Sialik Asit
TLC İnce Tabaka Kromatografisi
SEMBOLLER: cc Kübik Santimetre cm Santimetre dk Dakika g Gram L Litre M Molar MA Molekül Ağırlığı mm Milimetre mg Miligram
mIU Mili Uluslararası Ünite
ml Mililitre µg Mikrogram µl Mikrolitre µm Mikrometre µM Mikromolar ng Nanogram nM Nanomolar nl Nanolitre ºC Santigrat Derece
viii ÖZET
Amaç: Bu çalışmanın amacı KDN’nin (2-keto-3-deoxy-D-glycero-D-galacto-nononic acid) kanser hücresinde bağlandığı yeri göstermektir. KDN molekülü sialik asit ailesinin farklı bir türüdür. Bu molekül kanser hücrelerinde normal hücrelere oranla artmış düzeylerde bulunmaktadır. KDN’nin alabalık spermi üzerinde GM3 (monosialodihexosyl ganglioside)’e bağlı olduğu bilinmekle beraber insanda normal hücrede ve kanser hücresinde hangi yapıya bağlı olduğu henüz netlik kazanmamıştır. Yöntem: Çalışma in vitro modelde DU-145 prostat kanser hücre hattı kullanılarak gerçekleştirildi. Hücre hattı kontrol, Genz-123346 uygulanan ve Triptolide uygulanan olmak üzere 3 gruba ayrıldı. 1 numaralı hücre hattı grubu ilaç uygulanmayan kontrol grubu olarak belirlendi. Genz-123346 ile 2 numaralı hücre hattı grubunda GCS (Glukozilseramid Sentaz) enzim blokajı yapılarak GM3 sentezi engellendi. Triptolide ile 3 numaralı hücre hattı grubunda GD3S (GD3 Sentaz) enzimi bloklanarak GD3 sentezi engellendi. Her hücre grubunda GM3, GD3 ve KDN antikorları kullanılarak immünohistokimyasal boyama yapıldı. FTIR analiziyle hücrelerdeki karbonhidrat, lipit ve proteinlerdeki azalma ve artma gösterildi.
Bulgular: Herhangi bir ilaç uygulanmayan 1 numaralı kontrol hücre hattı grubunda GM3, GD3 ve KDN pozitif boyanırken, Genz-123346 ile GCS enzim blokajı yapılan 2 numaralı hücre hattı grubunda GM3 ve GD3 boyanmazken KDN boyanmıştır. GD3S inhibitörü Triptolide uygulanan 3 numaralı hücre hattı grubunda ise yalnızca GM3 ve KDN pozitif boyandı. FTIR ile yapılan ölçümlerde Triptolide ile apoptozise gidiş gösterilmiş, Genz-123346’nın ise hücre canlılığı üzerine negatif etkisi olmadığı belirlenmiştir. Şeker yapılarındaki azalma ortaya konmuş, boyamalarla elde ettiğimiz sonuçlar pekiştirilmiştir.
Sonuç: KDN’nin bağlı olduğu yeri tespit etmek kanser tedavisi araştırmalarında yeni hedefler seçilmesinde önem taşımaktadır. Hem GM3 inhibisyonunda hem de GD3 inhibisyonunda KDN’nin inhibe olmadığı gösterilmiş ve bu şekilde KDN’nin GM veya GD serisi gangliozidlerden herhangi biri üzerine spesifik ve yalnızca bağlı bulunmadığı, farklı yerlerde de olabileceği ortaya konmuştur.
Anahtar Kelimeler: KDN; GM; Gangliozid; Prostat Kanseri; Triptolide; Genz-123346
ix SUMMARY
Purpose: The aim of this study is to investigate where KDN (2-keto-3-deoxy-D-glycero-D-galacto-nononic acid) binds on cells. KDN molecule is a different kind of sialic acid family. The level of this molecule is increased in cancer cells comparison to normal cells. Although KDN is known to be bound to GM3 on trout sperm, to which structure it is bound on normal and cancer cells remain still unclear.
Method: In this study Du-145 prostat cancer cell line is used. Cell line is divided into 3 groups: control, Genz-123346 treated and Triptolide treated. Number 1 cell line group is defined as control group. GM3 synthesis is inhibited via GCS enzyme blokage in Genz-123346 treated number 2 cell line group. GD3 syntesis is inhibited in Triptolide treated number 3 cell line group. GM3, GD3 and KDN immunohistochemical staining is performed on each cell group. FTIR anlysis showed reduction and increase in carbohydrates, lipids and proteins in the cells.
Results: While non-treated number 1 cell line group stained positive with all of GM3, GD3 and KDN, GCS (Glucosylceramide Synthase) enzyme blocked with Genz-123346 number 2 cell line group stained with only KDN. Furthermore GD3S (GD3 Synthase) inhibitör Triptolide treated number 3 cell line group stained with GM3 and KDN. Measurements with FTIR showed apoptosis with triptolide and it had been determined that Genz-123346 had no negative effect on cell viability. The decrease in sugar constructions has been revealed and the results we have obtained with staining have been reinforced.
Conclusion: Determining where KDN is connected is important in selecting new targets for cancer treatment research. It has been shown that KDN doesn’t inhibit both GM3 inhibition and GD3 inhibition and thus it has been demonstrated that the KDN may also be in different places, specific and not only attached to any of the gangliosides of the GM or GD series.
10 1. BÖLÜM 1.1. GİRİŞ VE AMAÇ:
Prostat erkek genital sisteminde yer alan, mesanenin çıkımını çevreleyen ve meninin akışkanlığında rol oynayan bir salgı bezidir (Şekil 1). Salgılamayı yapan hücrelerden ve bunlara ait destek çevre dokusundan meydana gelmiştir. Bezler tek katlı ya da yalancı çok katlı (psödostratifiye) prizmatik epitel ile döşelidir (Şekil 2). Lümen, glikoproteinden zengin ve bazen kalsiyum birikme alanı olan prostatik taşlar (corpora amylacea) içerir. Hücreler bol miktarda granüllü endoplazmik retikulum ve golgi aygıtına sahiptir. Salgı ürünleri prostat-özgün asit fosfataz, prostat-özgün antijen, amilaz ve fibrinolizin içerir [1].
Prostat kanseri bu salgı yapan hücrelerin anormal gelişimi ile ortaya çıkmaktadır. Erken dönemde tanı konamadığı taktirde prostat kanseri lenf ve kan damarları yoluyla çevre dokulara yayılır. En sık yayılım yeri çevresindeki lenf düğümleri ve kemiklerdir. Ek olarak akciğer, karaciğer ve diğer organlara da yayılabilir.
11 Şekil 2. Prostat bezi [3].
Prostat kanseri erkeklerde deri kanserinden sonra en sık görülen kanserdir. Kansere bağlı ölümlerde ise ikinci sırada yer alır. Prostat kanserinin 70 yaş üzeri erkeklerin %50’sinde, 90 yaş üzerindekilerin de hemen hemen hepsinde mikroskobik düzeyde bulunduğu hesaplanmaktadır. Sağlıklı bir erkeğin hayat boyu prostat kanserine yakalanma riski yaklaşık olarak %17’dir. Yani yaklaşık olarak her 6 erkekten biri prostat kanserine yakalanmaktadır. 1980’lerde PSA testinin bulunup klinik kullanıma girmesi ile hastalığa bağlı ölüm oranları azalmıştır.
Ülkemiz için kesin veriler mevcut olmamakla birlikte ABD’de yılda 250 bin erkekte prostat kanseri saptanmaktadır ve bu kişilerin %95’i 45 yaş üzerindedir.
Prostat kanserine neyin sebep olduğu tam olarak bilinmemektedir. Diğer kanser türlerinde olduğu gibi genetik bir yatkınlıkla birlikte olumsuz çevre koşullarının, bu hastalığın meydana gelmesinde rol aldığı düşünülmektedir. Prostat kanseri oluşumuna katkısı olabileceği düşünülen bazı risk faktörleri şunlardır:
12 Yaş
Ailede prostat kanseri varlığı Yağlı beslenme
Hormonlar Kadmium
A ve D Vitaminleri
En önemli risk faktörleri yaş ve aile hikayesidir. Normal kontroller sırasında prostat kanseri araştırılması 50 yaş sonrası tavsiye edilirken, ailede prostat kanseri mevcudiyeti varsa bu araştırmalar 40 yaş sonrasında başlatılmaktadır. Tüm prostat kanserlerinin %10’unun babadan oğula kalıtım yoluyla geçtiği tahmin edilmektedir. Prostat kanseri olan hastaların erkek çocuklarında veya erkek kardeşlerinde prostat kanserine yakalanma riski 2-9 kat daha yüksektir. Prostat kanserine özgü bir başka özellik de hastalığın latent form ve klinik aktif form olmak üzere birbirinden çok farklı karakterde, iki ayrı formunun olmasıdır. Birinci gruptaki hastalık hayat boyu hiç tanı konmadan sessiz kalabilirken, ikinci grupta prostat dışına sıçrayarak yayılan ileri evre hastalık boyutları görülebilmektedir [4].
Çalışmada kullanılan Du-145 prostat kanseri hücre hattı, hipotriploid insan hücre hattıdır (Şekil 3). t(11q12q), del(11)(q23), 16q+, del(9)(p11), del(1)(p32) ve 6 başka kromozom belirteci ihtiva ederken N13 genellikle bulunmaz. Y kromozomunun tanımlanlanmamış bir kromozom segmentine translokasyonu mevcuttur. X koromozomu tek kopya olarak bulunur. Hücreler 69 yaşında, kafkas kökenli, kan grubu O Rh+, erkek bireye aittir. Adenokarsinoma (grade II) prostat primerinden elde edilmiştir. Hücre hattı hormon sensitif değildir. Hücreler prostat antijeni ekspress etmezler. Du-145 ve orijinal tümör hücre hatlarının her ikisinde de ultrastrüktürel analizlerinde mikrovilluslar, tonofilamentler, desmosomlar, mitokondria, golgi ve lizozomlar gözlenmiştir [5].
13
Şekil 3. Du-145 Prostat Kanser Hücre Hattı hücrelerinin kontrast mikroskop altındaki görüntüsü [6].
Kanser hücreleri farklı morfolojik özelliklere sahip hücrelerden oluşmuştur. Her bir tümör tipi için değişen yüzey belirteçlerini içeren bu hücreler farklı proliferasyon kapasitelerine sahip heterojen populasyonlar oluştururlar. Bu heterojenite, genetik ve mikroçevresel farklılıklarla açıklanabildiği gibi hücrelerin farklılaşabilme dereceleriyle de açıklanabilir. Kanser tedavisi amaçlanırken kanser hücresine ait sinyal yolakları, genetik yapı, bağlantı kompleksleri gibi kanserin ortaya çıkış mekanizması kadar yüzey antijenlerinin tespiti de oldukça önemlidir. Kanser hücresinde normal hücreye göre daha fazla eksprese edilen bazı protein, karbonhidrat ve lipit yapılarının ve fonksiyonlarının araştırılması kanser tedavisinde yeni protokollerin oluşturulmasına neden olmaktadır. Yeni yapılan çalışmalar, çeşitli glikolipitlerin kanser hücresinin invazyon, yeni damar yapıları oluşturma ve metastazında immün sistemden kaçış için kullanıldığını göstermektedir [7] [8]. Tedavi olarak glikolipitlerin
14
çeşitli yollarla bloke edilmesi denenmiş ve bazı glikolipitlerin blokajının tümör hücresinin metastazını azalttığı, hatta kanser hücresini apoptozise götürdüğü belirlenmiştir [9]. Gangliozidler, glikolipit yapısındadır ve günümüzde kullanılan bazı kanser ilaçlarının anti-kanser etkilerini gangliozid moleküllerini bloke ederek gösterdiği belirlenmiştir [10].
Gangliozidler, beynin gri maddesinde, sinir dokusunda, hücre zarlarının dış yüzeylerinde, eritrositlerin stromasında bulunur. Yüzey belirteci olarak, hücre hücre iletişiminde rol alır. Belirli hipofizer hormonlar ve bazı bakteri toksinleri (Kolera toksini gibi) için reseptör görevi görür. Karsinogenezisde doku büyümesinde ve farklılaşmasında görev alırlar [11]. Gangliozidler sadece hücre membranında değil glikolipit yapılarından ötürü lipit yapı içeren içeren nucleus membranı gibi hücresel zarların yapısında da bulunur [12]. Eksikliğinde santral sinir sistemiyle ilgili gelişim anormallikleri oluşurken, gangliozidlerin yıkımında defekt olursa yıkımdan sorumlu lizozomal enzime göre kas ve iskelet sistemi, karaçiğer, dalak ve santral sinir sitemi gibi organlarda birikim ve buna bağlı olarak sfingolipidozlar denen bir grup hastalığa ismini veren patolojiler ortaya çıkar [13].
Vücutta gangliozid yapımı ve yıkımı belirli bir dengede olmak zorundadır. Ancak kanser hücrelerinde bu mekanizma değişmektedir. Kanser hücreleri gangliozidler ve bazı diğer glikolipitleri immün sistemden kaçmak için, anjiogenez, invazyon ve metastaz yapmak için kullanmaktadır [7] [8].
15 Şekil 4. GM3’ün biyokimyasal yapısı[14].
16
Gangliozidler seramidden sentezlenen bir ya da daha fazla sialik asit içeren glikolipitlerdir. Yapısı seramid+oligosakkarid ve 1 veya daha fazla N-Asetil Nöraminik Asid’den oluşmuştur. Gangliozid sentezi seramidin GCS enzimi ile Glukoseramid (Glc-Cer)’e dönüşümü ile başlar. Galaktoziltransferaz I enzimi ile Laktozilseramid (Lac-Cer, Gal-Glc-Cer) oluşur. GM3 Sentaz enzimi ise aradığımız gangliozid olan GM3’ün (Monosialodihexosyl Ganglioside) sentezlenmesini sağlar (Şekil 6). Diğer gangliozidler GM3 (Şekil 4) üzerinden sentezlenir. GM2 GalNac-T (B4GALNT1, Beta-1,4 N-acetyl galactosaminyl transferase 1) enzimi ile GM3’den sentezlenirken. GD3 (Disialosyllactosylceramide) (Şekil 5) ise GD3S (GD3 Synthase, Alpha-N-acetyl neuraminide alpha-2,8-sialyl transferase) enzimi ile yine GM3’den sentezlenir. GM2 GM3’den, GM1 ise GM2’den sentezlenir. Yani GM3 (Şekil 6) diğer gangliozidlerin sentezi için anahtar konumdadır [16]. Yapılan çeşitli çalışmalarda seramidden GM3’ün yani bütün gangliozidlerin sentezinin GCS basamağında çeşitli moleküller ile başarılı şekilde inhibe edilebildiği gösterilmiştir [17]. Enzim inhibisyonunda temel olarak kullanılan moleküller Genz-123346, PPPP, PMPP, PDMP, NB-DNJ (N-butyl-deoxynojirimycin) ve Genz-529468 olarak sıralanabilir [17]. GM3’den GD3 sentezindeki GD3 sentaz enzimi ise yapılan çalışmalarda Triptolide isimli molekül ile bloke edilebilmiştir [7].
KDN (2-keto-3-deoxy-D-glycero-D-galacto-nonanic acid) ise sialik asit ailesinin farklı bir türüdür. KDN ortak bir Sia içinde yer alan C5 pozisyonunda aseto-amid grubu yerine hidroksil grubu eklenmesi ile oluşan, biyosentezi ve fonksiyonu açısından sialik asit ailesinin önemli bir üyesidir. KDN çeşitli sialidazlara duyarsızlık ve polisialik asidinin uzaması için bir “stop sinyali” içerme gibi farklı özelliklere sahiptir. Memeli hücrelerinde KDN genellikle çok düşük seviyelerde ifade edilmektedir. İlk olarak Inoue ve ark tarafından polisialo-glikoprotein olarak izole edilmişlerdir [18]. KDN molekülü iki ayrı yolak sonucunda sentez edilir. Bunlardan birincisi Neu5Ac den de-asetilasyon ve de-aminasyon yolu ile sentezlenmesi, diğer bir yol ise farklı enzimlerle de novo olarak eldesidir. Bununla beraber Neu5Ac ile pek çok özelliklerini paylaşırlar:
1) KDN bakterilerden, insanları da içeren yüksek vertebralılara kadar pek çok canlıda bulunur,
17
2) Her tip glikokonjugat içerisinde (glikolipit, glikoprotein, kapsüler polisakkarid) bulunur,
3) KDN rezidüleri Neu5Ac’nin bulunduğu yerlerdeki tüm glikanlara bağlı olarak bulunabilir,
4) KDN, mannozdan de novo olarak bir seri reaksiyon sonucunda, Neu5Ac sentezine benzer şekilde N-asetilmannozamin prekürsör şeker olarak kullanılarak sentezlenebilir. KDN sonrasında CMP-KDN ye aktive olur ve şeker rezidülerine transfer edilir [19].
1.1.1. GCS Enzimi:
GCS enzimi seramidden gangliozid sentezinde kritik öneme sahiptir. GCS enzimi seramidden Lac-Cer sentezler, Lac-Cer’den de GM3 sentezlenir. Diğer gangliozidler GM3’den sentezlenir. GCS sentezinin engellenmesi Gangliozid sentezini de inhibe edecektir [17] (Şekil 6).
1.1.2. GD3 Sentaz Enzimi:
GD3 Sentaz enzimi GM3’den GD3 sentezinde görev alan enzimdir. GD ve GT gangliozid serilerinin sentezi için kritik öneme sahiptir [16].
18
Şekil 6. Seramidden Gangliozid sentezinin basamakları [20].
1.1.3. GCS İnhibitörleri:
GCS enziminin inhibisyonunda kullanılan moleküller Genz-123346, PPPP, PMPP, PDMP, NB-DNJ (N-butyl-deoxynojirimycin) ve Genz-529468 olarak sıralanabilir [17]. Çalışmamızda Genz-123346 kullanılmıştır.
1.1.4. GD3 Sentaz İnhibitörleri:
GD3S enzimi, ShRNA ve bir kanser ilacı olan Triptolide ile bloke edilebilmektedir [7]. GD3S enzimi inhibisyonu için çalışmamızda Triptolide kullanılmıştır.
GM3 seramidden sentezlenen tek sialik asitli NANA-Gal-Glc-seramid yapısında glikosfingolipit ailesinden bir ganglioziddir. KDN daha önce alabalık sperminde GM3
19
ile birlikte olarak gösterilmiş [21] fakat insan hücresinde nereye bağlı olduğu tespit edilmemiştir.
Bu projenin amacı gangliozid sentez yolaklarının kullandığımız kimyasallarla ne düzeyde etkileşime uğradığını saptamak, inhibe edildiğini göstermek, KDN’nin Du-145 insan prostat kanser hücrelerinde bulunduğu yeri aydınlatmaya çalışarak kanser tedavisi araştırmalarına yeni bir çalışma alanı kazandırmaktır. Bu proje ile KDN veya bağlı bulunduğu molekül hedef alınarak planlanan kanser tedavisi modellerinin çalışılması mümkün olabilecektir.
20 2. BÖLÜM 2.1. GEREÇ VE YÖNTEMLER:
2.1.1. Çalışmada Kullanılan Kimyasal Maddeler ve Cihazlar:
Kimyasal Üretici Firma Kullanım Amacı
FBS (Fetal Bovine Serum) Biowest (S1810) Besi ortamına eklenen besleyici madde Penisilin/Streptomisin Thermo Scientific
(SV30010)
Besi ortamına eklenen antibiyotik
Amfoterisin B Lonza (17-836E) Besi ortamına eklenen antifungal
PBS (Phosphate Buffer Saline)
Biovision (2113-500) Pasaj yapılırken hücrelerin yıkanmasında kullanılan tampon çözelti
Tripsin-EDTA Lonza (BE12-167F)
Kültüre edilen adheziv hücreleri flask yüzeyinden ayırmak için kullanılan kimyasal
L-Glutamine Thermo Scientific
(SH3003401)
Besi ortamına eklenen (RPMI-1640’a) amino asit Count&Viability kit Milipore, ABD Hücre sayımı ve canlılık
testi
Etanol Merck Fiksasyon
Tablo 1. Çalışmada kullanılan kimyasal maddeler, üretici firma bilgileri ve kullanım amaçları.
21
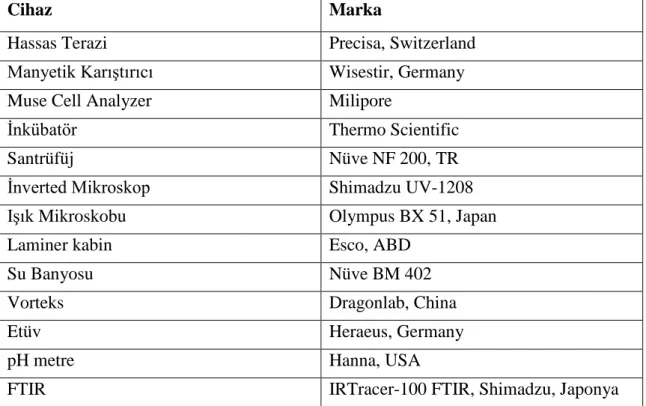
Cihaz Marka
Hassas Terazi Precisa, Switzerland
Manyetik Karıştırıcı Wisestir, Germany
Muse Cell Analyzer Milipore
İnkübatör Thermo Scientific
Santrüfüj Nüve NF 200, TR
İnverted Mikroskop Shimadzu UV-1208
Işık Mikroskobu Olympus BX 51, Japan
Laminer kabin Esco, ABD
Su Banyosu Nüve BM 402
Vorteks Dragonlab, China
Etüv Heraeus, Germany
pH metre Hanna, USA
FTIR IRTracer-100 FTIR, Shimadzu, Japonya
Tablo 2. Çalışmada kullanılan cihazlar ve markaları.
2.1.2. Çalışmada Kullanılan Hücre Hatlarının Temini ve Üretilmesi:
Prostat kanseri hücre hatları (Du-145) Amerikan Tip Kültür Koleksiyonu’ndan (ATCC) temin edilmiştir.
DU-145 insan prostat kanseri hücre hattının çoğaltılmasında ve sürdürülmesinde RPMI 1640 besi ortamı kullanılmıştır. 500 ml’lik steril besi ortamının içerisine %1 oranında Penisilin/Streptomisin, %10 ısı ile inaktive edilmiş Fetal Sığır Serumu, %1 oranında Amfoterisin B ve %1 oranında L-Glutamin eklenerek, 37 °C sıcaklıkta, %5 CO2 ve nem içeren inkübatörde çoğaltıldı.
Hücre hatları canlılık, çoğalma ve enfeksiyon açısından inverted mikroskopta günlük olarak izlendi. Flasklarda %80’in üzerinde hücre yoğunluğu gözlendiğinde hücreler pasajlanarak çoğaltıldı.
22
2.2. ÇALIŞMADA KULLANILAN İLAÇLARIN VE KIMYASALLARIN HAZIRLANMASI:
2.2.1. Genz-123346‘nın stok solüsyonlarının hazırlanması:
Genz-123346 (MA: 554.637 g/mol) kimyasalından 100 mM 15 ml ara stok hazırlanmıştır. Kimyasalların ara stokları kullanılarak aşağıda belirtildiği şekilde uygulama dozları ayarlanmıştır.
GENZ 100 µM ilaç stoğu RPMI Besi yeri Toplam Hacim
10 nM 1 µl 14999 µl 15 ml
100 nM 15 µl 14985 µl 15 ml
1000 nM 150 µl 14850 µl 15 ml
2.2.2. Triptolide’in stok solüsyonun hazırlanması:
Triptolide (MA: 360.406 g/mol) kimyasalından 100 mM 15 ml ara stok hazırlanmıştır. Kimyasalların ara stokları kullanılarak aşağıda belirtildiği şekilde uygulama dozları ayarlanmıştır.
TRİPTOLİDE 100 µM ilaç stoğu RPMI Besi yeri Toplam Hacim
20 nM 3 µl 14997 µl 15 ml
40 nM 6 µl 14994 µl 15 ml
60 nM 9 µl 14991 µl 15 ml
2.2.3. Tampon Çözeltisinin Hazırlanması:
8.78 g NaCl (Sigma Aldrich), 0,13 g Na2PO4.2H2O (Merck) ve 4,040 g Na2PO4.7H2O (Merck) maddeleri hassas terazide tartılarak 1 L distile su içerisinde
23
çözülüp pH 7.3’e ayarlanarak fosfat tamponu (PBS, Phosphate Buffered Saline) hazırlandı.
2.2.4. Paraformaldehid’in Hazırlanması:
8 g Paraformaldehit (PFA) (Merck) hasas terazide tartılarak 100 ml distile suya eklendi. Sıcaklığı 58 oC’yi geçmeyecek şekilde ölçülerek ve ısıtıcı üzerinde sürekli karıştırılarak renginin saydamlaşması beklendi. Rengi saydamlaştıktan sonra soğuması beklenerek süzüldü. Hazırlana %8’lik paraformaldehidin kullanıma hazır hale getirilmesi için aynı miktarda fosfat tamponuyla karıştırılarak ve %4’lük paraformaldehid hazırlandı. Daha sonra pH 7.2 olacak şekilde ayarlanarak immünofloresan boyama yöntemi uygulanacak hücrelerin fiksasyonunda kullanıldı.
2.3. DENEYİN KURGULANMASI VE DENEY GRUPLARI:
Çalışmamız hücre kültürü çalışmasıdır, araştırmamızda hücre serileri kullanılacağı için Etik Kurulu onayı gerekmemektedir. Projemizde kullanılan hücre serileri ve deney grupları;
Grup 1: İnsan prostat kanseri hücre hattı hücreleri (DU-145) Kontrol grubu Grup 2: İnsan prostat kanseri hücre hattı hücreleri (DU-145) Genz-123346 uygulanmış grup
Grup 3: İnsan prostat kanseri hücre hattı hücreleri (DU-145) Triptolide uygulanmış grup
24
2.4. ÇALIŞMADA KULLANILAN YÖNTEMLER:
2.4.1. Hücre Sayısı ve Canlılığının Belirlenmesi:
Hücre sayısı ve canlılığı Muse™ Count & Viability Kit (Katalog No: MCH100102) kullanılarak Muse™ Cell Analyzer ile belirlenmiştir (Şekil 7). Yöntem, reaktif içindeki DNA-bağlayıcı boyaların, membran bütünlüğünün hücreler tarafından kaybedilmesi ile ölü ve canlı hücre nukleusunun ayrı ayrı boyanmasını prensibine dayanmaktadır. Hücreler 1x104 hücre/kuyucuk olacak şekilde kuyucuklu plaklara (6-well plate) ekildi. 24 saat sonra, hücreler plaklara tutunduğunda GCS inhibitörü olarak Genz-123346 (Boc Sciences, Katalog No: 491833-30-8) ve GD3S inhibitörü olarak Triptolidin (Biorbyt, Katalog No: orb106086) farklı konsantrayonları ile 24, 48 ve 72 saat inkübe edildi. İnkübasyon süresinin sonunda hücreler %0.05 tripsin ile kaldırıldı. Tripsin inaktivasyonu için RPMI 1640 eklenerek 5 dk 1000 rpm ile santrifüj edildi ve sonra kültür medyumu eklenerek resüspansiye edildi. Hücre süspansiyonundan 20 μl ve Muse™ Count & Viability reaktifinden 380 μl ilave edilerek 5 dk oda sıcaklığında, karanlık ortamda inkübe edilerek cihazda ölçüm yapıldı.
25
Şekil 7. Muse™ Cell Analyzer cihazında % canlılık değerlerinin belirlenmesi [22].
2.4.2. İmmunohistokimyasal Yöntem:
Tek tabakalı olarak kültüre edilen hücreler 24’lü plaklarda çoğaltıldı ve paraformaldehit ile fikse edildi. Primer antikorlar KDN (özel üretim), Anti-Ganglioside GD3 Antibody (ab11779) ve Anti-GM3 Monoclonal Antibody (AMS.A2582) ile muamele edilen hücreler nemli ortamda bir gece 4°C’de inkübe edilip modifiye streptavidin-peroksidaz yöntemi uygulandı. DAB ile inkübasyon sonrasında örneklere zemin boyama (counterstain) için Mayer’s hematoksilen kullanıldı. Olympus BX-51 ışık mikroskobu ve Olympus C-5050 dijital fotoğraf makinası kullanılarak moleküllerin immünoreaktivitesi değerlendirildi. Boyanmalar 2 bağımsız gözlemci tarafından semi-kantitatif bir skalaya göre (az, orta ve koyu boyanma) derecelendirildi.
26
2.4.3. (Fourier Transform Infrared Spectroscopy Assay) FTIR Analizi:
Fourier Dönüşümlü Infrared Spektrofotometre (FTIR), FTIR Spektrum Cihazı organik bileşiklerin tanımlanmasında kullanılır. Optik izomerler dışında bütün bileşiklerin IR spektrumu birbirinden farklıdır. IR bölgesi elektromanyetik spektrumun görünür bölgesi ile mikro dalga bölgesi arasında yer alır.
Cam infraredi kuvvetli olarak absorpladığından deneyler için KBr, AgCl, NaCl veya CaF2‘den yapılmış kaplar veya prizmalar kullanılır. Katı maddeler KBr ile toz haline getirilerek ve belli bir ölçüde preslenerek IR spektrumu alınır. Veyahut nujol veya çözeltide incelenir. Bu amaç için çözücü olarak sadece CCl4 veya CS2 gibi çok az absorpsiyon bandları gösteren çözücüler kullanılır.
27 Kullanım Alanları:
IR spektrumlarının kullanım alanları çok yönlüdür; Bir reaksiyonun yürümesi veya bir kramatografi ayrılım gidişi belirli zaman aralıklarında yapılan deneylerle ve IR spektrumu alınmasıyla takip edilebilir.
Preperatif çalışma yapan kimyacılar için IR spektrumu kullanımı bağların ispatı için önem taşır. Çünkü, bir reaksiyonda istenen maddenin oluşup oluşmadığına, yan ürünlerin çıkıp çıkmadığına ve oluşan ürünlerin neler olduğuna yanıt verir. Çözücü etkileri ve asosiyasyon dengeleri hakkında yararlı bilgiler verir.
IR Spektrometre başlıca üç kısımdan oluşmuştur: Işın Kaynağı
Monokromatör Alıcı
Kaynaktan çıkan ışının yarısı örnekten, diğer yarısı referanstan geçer. Monokromatörden geçen ışın, dalga boylarına ayrılıp dedektör üzerine düşer. Böylece elektrik sinyaline çevrilir. Çözelti hazırlamanın zor olduğu bileşikler için ATR (Attenuated Total Reflectance) tekniği uygulanır. Bu spektrum, örneğin kalınlığından bağımsızdır. Dolayısıyla uygulanması kolay ve soğurganlığı çok fazla maddeler durumunda çok yararlıdır. ATR tekniği, biyomelüküllerin yapısı ve fonksiyonu ile ilgili değerli bilgiler verir (Şekil 8) [24] [25] [26].
Fourier transform infrared spectroscopy (FTIR) düşük kurulum ve çalıştırma maliyeti ile ve ayrıca zaman kazandıran bir teknik olduğundan, hücreler ve dokular gibi biyolojik örneklerin taranması için kullanılmaktadır. Bir hücre döngüsünde veya hücre ortamına ilaç ilave edildikten sonra biyokimyasal değişiklikleri belirlemek için çeşitli hücre hatlarında (örneğin kanser hücreleri, kök hücreler) kullanılabilir [26] [27] [28] [29].
Hücrelerin FTIR spektrumu, lipidler, proteinler, karbonhidratlar, nükleik asitler ve fosforile edilmiş bileşikler gibi tüm hücresel unsurlardan kaynaklanan IR sinyallerinin bir super impozisyonudur. Orta kızılötesi aralıktaki (4000-400 cm-1) spektral değişiklikleri izleyerek hücre büyümesi, hücre döngüsü, proliferasyon, farklılaşma, hücre ölümü ve apoptoz gibi hücresel olaylar ve hücrelerdeki makromoleküllerin moleküler kompozisyonu, dinamiği ve yapısıyla ilgili kapsamlı bilgi edinmeyi sağlar. [28] [30] [31].
28 Örnek Hazırlama
FTIR spektroskopisi için, Du-145 hücre hattı yukarıda tarif edildiği gibi kültürlenmiştir. T75 cam kaplarda hücre hatları yetiştirildi ve ortam her hafta 2-3 kez değiştirildi. Hücreleri hasat etmeden önce madde atıldı ve hücreler ölü hücreleri çıkarmak için 1 ml % 0.05'lik tripsin (Sigma Aldrich) ile hızlıca yıkandı. Daha sonra hücreler kültür ortamından 1ml tripsin tamponu ile 37 °C'de %5’lik CO2 ortamında 5 dakika süreyle muamele edilerek ayrıldı. Tripsin reaksiyonu, hücrelerin %10 FBS (fetal sığır serumu) ile 5 ml kültür ortamı ile yıkanmasıyla durduruldu. Hücreler, 2 dakikalık bir santrifüjle (1000rpm) pellet haline getirildi ve tripsin ile kültür ortamının tamamen çıkarılmasını sağlamak için üç kez steril izotonik solüsyon (% 0.9 NaCl) ile yıkandı. Hücreler daha sonra ATR-FTIR analizi için NaCl çözeltisinde yeniden süspanse edildi.
ATR-FTIR Spektroskopik Ölçümleri
Tüm ölçümler, bir attenuated total reflection (ATR) birimi ile birleştirilmiş ve bir DLATGS detektörü ile donatılmış bir IRTracer-100 FTIR spektrometresi (Shimadzu, Japonya) ile oda sıcaklığında gerçekleştirilmiştir. 2 µl hücre süspansiyonu miktarı (yaklaşık 1x106 hücre/ml) Diamond/ZnSe mercek tek yansımalı ATR plakası üzerine kondu ve ölçümlerden önce hafif atmosfer koşullarında 10 dakika kurutuldu. Her koşul için üç bağımsız kültür yetiştirildi ve her kültürden alınan üç numune ölçüldü ve böylece ölçümler üçlü hale getirildi. Her bir ölçüm için en az 10 spektrum 4000-800 cm-1 aralığında kaydedildi. Her interferogram için 4 cm-1 spektral çözünürlükte toplam 128 tarama yapıldı. Hava spektrumu, ATR plakası boşaldığında arka plan olarak kaydedildi.
29 Veri Analizleri
IR spektrumları, fark spektrumları ve Student t-testlerinin düzeltilmesi ve Temel bileşen analizi (PCA, Principal component analysis), MATLAB altında çalışan Kinetics yazılımı ile (Prof. Dr. Erik Goormaghtigh tarafından sağlanmıştır) kullanılarak gerçekleştirildi.
30 3. BÖLÜM 3.1. BULGULAR:
3.1.1. Hücre Sayısı ve Canlılığının Belirlenmesi Sonuçları:
GCS İnhibitörünün Sitotoksik Etkileri:
GCS inhibitörü’nün Du-145 insan prostat kanseri hücre hattında hücre canlılığı üzerine etkileri, hem artan doza (10-1000 nM) hem de zamana (24., 48. ve 72. saat) bağlı olarak çalışıldı.
GCS inhibitörü ile 24, 48 ve 72 saat muamele sonunda, Du-145 hücre hatlarında 10-1000 nM arasındaki konsantrasyonlar kontrol ile kıyaslandığında anlamlı sitotoksik etki saptanmadı. Genz-123346’nın hücreler üzerine sitotoksik olmadığı görüldü. 72 saat sonunda 1000 nM dozunda dahi sırasıyla Du-145 hücre hatlarında %95,5, %96, %95,9 olarak bulundu. Genz-123346’nın IC-50 değeri bulunamadı (Şekil 12).
31
Du-145, Genz, 24 h, Kontrol Du-145, Genz, 24 h, 10 nM
Du-145, Genz, 24 h, 100 nM Du-145, Genz, 24 h, 1000 nM
Şekil 9.Genz-123346’nın çeşitli dozlardaki 24. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı).
32
Du-145, Genz, 48 h, Kontrol Du-145, Genz, 48 h, 10 nM
Du-145, Genz, 48 h, 100 nM Du-145, Genz, 48 h, 100 nM
Şekil 10.Genz-123346’nın çeşitli dozlardaki 48. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı).
33
Du-145, Genz, 72 h, Kontrol Du-145, Genz, 72 h, 10 nM
Du-145, Genz, 72 h, 100 nM Du-145, Genz, 72 h, 1000 nM
Şekil 11. Genz-123346’nın çeşitli dozlardaki 72. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı).
34
Şekil 12. GCS inhibitörü Genz-123346’nın Du-145 insan prostat kanseri hücre hattında zamana bağlı olarak hücre canlılığı (%) üzerindeki etkileri.
GCS inhibitörü’nün Du-145 insan prostat kanseri hücre hattında 1, 100, 1000 nM konsantrasyonlarının zamana (24, 48 ve 72 saat) bağlı sitotoksik etkisi istatistiksel olarak anlamlı olmadığı bulunmuştur (Şekil 12).
GD3S İnhibitörünün Sitotoksik Etkileri:
GD3 inhibitörü’nün Du-145 prostat kanseri hücre hattında hücre canlılığı üzerine etkileri, hem artan doza (20-60 nM) hem de zamana (24., 48. ve 72. saat) bağlı olarak çalışıldı.
GD3 inhibitörü ile 24, 48 ve 72 saat muamele sonunda, Du-145 hücre hatlarında 10-1000 nM arasındaki konsantrasyonlar kontrol ile kıyaslandığında anlamlı
98 ,5 97 ,2 95 ,8 95 ,5 98 ,5 98 97 ,4 96 98 ,5 97 ,8 96 ,1 95 ,9 K O N T R O L 1 0 N M 1 0 0 N M 1 0 0 0 N M
GENZ DU-145 CELL LINE IC-50
35
sitotoksik etki saptandı. 72 saat sonunda 60 nM dozunda bütün hücre hatlarında IC-50 değerleri tespit edildi (Şekil 16).
Du-145, Triptolide, 24 h, Kontrol Du-145, Triptolide, 24 h, 20 nM
Du-145, Triptolide, 24 h, 40 nM Du-145, Triptolide, 24 h, 60 nM
Şekil 13. GD3S inhibitörü Triptolide’in çeşitli dozlardaki 24. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı).
36
Du-145, Triptolide, 48 h, Kontrol Du-145, Triptolide, 48 h, 20 nM
Du-145, Triptolide, 48 h, 40 nM Du-145, Triptolide, 48 h, 60 nM
Şekil 14. GD3S inhibitörü Triptolide’in çeşitli dozlardaki 48. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı).
37
Du-145, Triptolide, 72 h, Kontrol Du-145, Triptolide, 72 h, 20 nM
Du-145, Triptolide, 72 h, 40 nM Du-145, Triptolide, 72 h, 60 nM
Şekil 15. GD3S inhibitörü Triptolide’in çeşitli dozlardaki 72. saatteki etkileri (Du-145 insan prostat kanseri hücre hattı).
38
Şekil 16. GD3S inhibitörü Triptolide’in Du-145 insan prostat kanseri hücre hattında zamana bağlı olarak hücre canlılığı (%) üzerindeki etkileri.
GD3S inhibitörü’nün Du-145 insan prostat kanseri hücre hattı üzerinde 20, 40 ve 60 nM konsantrasyonlarının zamana (24, 48 ve 72 saat) bağlı sitotoksik etkisi istatistiksel olarak anlamlı bulunmuştur (Şekil 16).
3.1.2. İmmunohistokimya Boyama Sonuçları:
GCS inhibitörü ile muamele edilen Du-145 insan prostat kanseri hücre hattında GM3, GD3 ve KDN’de meydana gelen değişimler, immunohistokimya boyama yöntemi ile belirlendi. Hücreler 1000 nM dozundaki GCS inhibitörü ile 24 saat muamele edildi. Kontrol grubuna GCS inhibitörü eklenmedi. GCS inhibitörü eklenmeyen kontrol grubu ile kıyaslandığında, bütün hücre hatlarında Genz-123346 uygulama sonrası boyanmada GM3 ve GD3 antikorları ile yapılan immunfleurosan boyanmada anlamlı azalma tesbit edilmiştir (Şekil 17, Şekil 18), KDN ile yapılan boyamada ise azalma tespit edilmemiştir (Şekil 19).
97 ,6 90 ,1 87 ,3 84 ,1 95 ,8 79 ,3 65 ,5 55 ,3 93 ,1 58 ,3 50 ,2 47 ,2 K O N T R O L 2 0 N M 4 0 N M 6 0 N M
TRIPTOLIDE DU-145 CELL LINE IC-50
39
Şekil 17. GCS inhibitörü Genz-123346’nın 1000 µM dozu ile 24 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GM3 gangliozidinde meydana gelen değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X).
40
Şekil 18. GCS inhibitörü Genz-123346’nın 1000 µM dozu ile 24 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GD3 gangliozidinde meydana gelen değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X).
41
Şekil 19. GCS inhibitörü Genz-123346’nın 1000 µM dozu ile 24 saat muamele edilen Du-145 insan prostat kanseri hücre hattında KDN’de meydana gelen değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X).
GD3 inhibitörü ile muamele edilen Du-145 insan prostat kanseri hücre hattında GM3, GD3 ve KDN’de meydana gelen değişimler, immunohistokimya boyama yöntemi ile belirlendi. Hücreler 60 nM dozundaki GD3 inhibitörü ile 72 saat muamele edildi. Kontrol grubuna GD3 inhibitörü eklenmedi. GD3 inhibitörü eklenmeyen kontrol grubu ile kıyaslandığında, bütün hücre hatlarında Triptolide uygulama sonrası immunfleurosan boyanmada GD3’de anlamlı azalma tespit edilmişken (Şekil 21), GM3 ve KDN’de azalma tespit edilmedi (Şekil 20, Şekil 22).
42
Şekil 20. GD3 inhibitörü Triptolide’in 60 µM dozu ile 72 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GM3 gangliozidinde meydana gelen değişimin ışık mikroskobu görüntüsü (Ölçek: 20X, 40X).
43
Şekil 21. GD3 inhibitörü Triptolide’in 60 µM dozu ile 72 saat muamele edilen Du-145 insan prostat kanseri hücre hattında GD3 gangliozidinde meydana gelen değişimin ışık mikroskobu görüntüsü(Ölçek: 20X, 40X).
44
Şekil 22. GD3 inhibitörü Triptolide’in 60 µM dozu ile 72 saat muamele edilen Du-145 insan prostat kanseri hücre hattında KDN’de meydana gelen değişimin ışık mikroskobu görüntüsü(Ölçek: 20X, 40X).
45
3.1.3. Fourier Transform Infrared Spectroscopy Assay (FTIR) Sonuçları:
Genz-123346:
Apoptosis gözlenmemektedir. Bu nedenle sadece GM3 blokajından dolayı meydana gelen değişiklikler gözlenmektedir. Prostat kanser hücre hattında α-helix yapısındaki proteinlerde (1654 ve 1549 cm-1) artış gözlenmektedir (Amide I ve Amide II bantlarının pozitif absorbans olması). Lipit sinyallerinde Triptolide uygulamasına nazaran daha fazla artış gözlenmektedir [Doymamış=CH: 3011 cm-1, doymuş CH2: 2922 ve 2853 cm-1, CH2 eğilme (bending) (1460 cm-1) ve C=O: 1745 cm-1].
Çalışmamızda karbonhidrat (şeker türevleri) sinyalleri ileaynı zamanda sialik asit sinyallerinin de azaldığı (1118, 1044 ve 1011 cm-1) gösterilmiştir (Şekil 23, Şekil 24, Şekil 25).
Triptolide:
Prostat kanser hücre hattı proteinlerinin düzenli yapıları azalmaktadır (Amide I ve Amide II bantlarının negatif absorbans olması): protein α-helix yapıları (1659 cm -1) azalırken beta yapraklar (1629 cm-1) artmıştır. Bu durum tedavi sonrasında, hücrede apoptosis meydana geldiğini göstermektedir.
Lipit sinyallerinde artış gözlenmektedir. [Doymamış =CH 3013 cm-1, doymuş CH2 2922 ve 2853 cm-1, CH2 eğilme (bending) 1464 cm-1 ve C=O 1741 cm-1] lipit sinyalleri artıyor. Bu spektrum dizisi, Triptolide uygulanmasıyla oluşan apoptozise bağlı plazma membran kabarcıklanma (blebbing) nedeniyle ayrıca gangliozidlerin, şeker türevlerinin (oligosakkaritler, monosakkaritler) ve sialik asitlerin azalmasına bağlı olarak da membran lipitlerinin yüzeye çıktığını göstermektedir. Karbonhidrat (şeker türevleri) sinyalleri (1180-900 cm-1) ve sialik asit sinyalleri azalmaktadır (1118, 1044 ve 1011 cm-1). Triptolide uygulanmasıyla apoptosis gerçekleştiği için değişen lipit ve protein sinyallerinin diğer biyomoleküllerden gelen sinyalleri baskıladığı gösterilmiştir (Şekil 23, Şekil 24, Şekil 25).
46
Şekil 23. Kontrol, Genz-123346 ve Triptolide ile muamele edilmiş Du-145 insan prostat kanseri hücre hatlarının FTIR spektrumu, 3100-2800 ve 1800-800 cm-1 spektrum bölgeleri.
47
Şekil 24. 3000-2800 ve 1800-800 cm-1 spektrum bölgeleri PCA skoruyla temel bileşenler analizi (Şekil 3.1.4.1’den elde edilmiştir). Çember içindeki değerler her temel bileşenin toplam varyasyon yüzdesini göstermektedir.
48
Şekil 25. FTIR spektrum farkı (treated-untreated) (Şekil 3.1.4.1’den elde edilmiştir). Du-145 hücre hatlarının Triptolide ve Genz-123346 uygulanmış FTIR sonuçları 3100-2800 cm-1 ve 1800-800 cm-1 aralığında gösterilmiştir. Spektrumlar baseline düzeltilmiş olup amid II bandına göre normalize edilmiştir.
49 4. BÖLÜM 4.1. TARTIŞMA:
Gangliozidler, beynin gri maddesinde, sinir dokusunda, hücre zarlarının dış yüzeylerinde, eritrositlerin stromasında bulunur. Yüzey belirteci olarak, hücre-hücre iletişiminde rol alır. Belirli hipofizer hormonlar ve bazı bakteri toksinleri (Kolera toksini gibi) için reseptör görevi görür. Karsinogenezisde doku büyümesinde ve farklılaşmasında görev alırlar [11]. Gangliozidler sadece hücre membranında değil glikolipit yapılarından ötürü lipit yapı içeren içeren nucleus membranı gibi hücresel zarların yapısında da bulunur [12]. Eksikliğinde santral sinir sistemiyle ilgili gelişim anormallikleri oluşurken, gangliozidlerin yıkımında defekt olursa yıkımdan sorumlu lizozomal enzime göre kas ve iskelet sistemi, karaciğer, dalak ve santral sinir sistemi gibi organlarda birikim ve buna bağlı olarak sfingolipidozlar denen bir grup hastalığa ismini veren patolojiler ortaya çıkar [13].
Vücutta gangliozid yapımı ve yıkımı belirli bir dengede olmak zorundadır. Ancak kanser hücrelerinde bu mekanizma değişmektedir. Kanser hücreleri gangliozidler ve bazı diğer glikolipitleri immün sistemden kaçmak için, anjiogenez, invazyon ve metastaz yapmak için kullanmaktadır [7] [8]. Tedavi olarak glikolipitlerin çeşitli yollarla bloke edilmesi denenmiş ve bazı glikolipitlerin blokajının tümör hücresinin metastazını azalttığı, hatta kanser hücresini apoptozise götürdüğü belirlenmiştir [9].
GCS inhibisyonu için yapılan az sayıdaki çalışmada Genz-123346 kullanılmıştır [32] [33]. Natoli TA ve Karman’ın 2010 yılındaki çalışmalarında Genz-123346 ile fareler tedavi edilmiştir. Tedavi edilen farelerin böbreklerindeki etkileri göstermiştir. In-vivo yapılan bu çalışma Genz-123346 ile tedaviyle immünohistokimyasal ve Western Blot metodları ile hem GSC’nin hem de GM3’ün azaldığını ortaya koymuştur [32]. Karman aynı sonucu fare akciğerinde Western Blot metodu ile bulmuştur [33]. Çalışmamızda ise benzer sonuçlar Du-145 insan prostat kanseri hücre hattı kültüründe elde edildi.
Haw-Young Kwon’un çalışmasında Triptolide ile yapılan GD3 inhibisyonu, SK-MEL-2 insan melanoma hücrelerinde hST8Sia I gen ekspesyonundaki azalma ile
50
gösterilmiştir [34]. Çalışmamızda Du-145 hücre hattında Triptolide ile yapılan GD3s inhibisyonu immunohistokimyasal boyamayla ve FTIR ölçümüyle gösterilmiştir.
KDN ve GM3 ilişkisi Thin Layer Chromatography (TLC), Western Blot ve immünoelektron mikroskobuyla Song Yu tarafından gösterilmiştir [21]. Çalışmamızda KDN’nin GM3’e spesifik olarak bağlı olup olmadığı ortaya konmaya çalışılmıştır. Fakat yaptığımız immunohistokimyasal boyama ve FTIR sonuçlarına göre GM3 ilişkisini reddetmemekle beraber herhangi bir şekilde yalnızca GM3’e spesifik bir şekilde bağlı olmaktan bahsetmenin mümkün olamayacağı ortaya konmuştur.
Gangliozid seviyelerinin artışının tümör prognozunu değerlendirmede kullanılabileceği mesane kanserinde serum ve dokulardaki GM3 miktarının arttığı HPTLC yöntemiyle gösterilmiştir [35]. 2015 yılındaki bir çalışmada GD3 gangliozidinin meme kanserinde Snail, Twist ve TGF-β1 ile epitelyo-mezenkimal geçiş ve metastazla ilişkisi ortaya konmuştur. In vitro SUM159 and MDA-MB-231 hücre hatlarında yapılan çalışmada yara iyileşmesi, invazyon ve kök hücre fonksiyonu için GD3 gereklidir. In vivo fare çalışmalarında ise GD3S’ın inhibe edilmesi metastazı önlemektedir [8]. İnsan kolon kanserinde gangliozid GM3’ün Cisplatin ile indüklenmiş apoptosis ile ilgili olduğu, siRNA ile GM3 sentaz ve dolayısıyla GM3 başta olmak üzere prekürsörü olduğu diğer bütün Gangliozidlerin azaltılmasının da HCT116 insan kolon karsinomu hücrelerini Cisplatin aracılı apoptozisten koruduğu, Bax, Bcl-2 ve p53 gibi serbest oksijen radikali hasarına bağlı apoptotik sinyal yolaklarının inhibe olduğu gösterilmiştir [10].
FTIR analizinde, Genz-123346 uygulandığında apoptosis gözlenmemektedir. Bu nedenle sadece GM3 blokajından dolayı meydana gelen değişiklikler gözlenmektedir. Prostat kanser hücre hattında α-helix yapısındaki proteinlerde 1654 ve 1549 cm-1 de artış gözlenmektedir (Amide I ve Amide II bantlarının pozitif absorbans olması). Lipit sinyallerinde triptolide uygulamasına nazaran daha fazla artış gözlenmektedir [doymamış=CH: 3011 cm-1, doymuş CH2: 2922 ve 2853 cm-1, CH2 eğilme (bending) 1460 cm-1 ve C=O: 1745 cm-1]. Bu spektrum dizisi, GM3 sentezini bloke etmeye neden olan Genz-123346 tedavisi sonrasında, hücre yüzeyinde bulunan şeker türevleri (oligosakkaritler, monosakkaritler), sialik asitler ve dolayısıyla da gangliozidlerde azalmanın bir sonucu olarak lipit yağ asitlerinin hücre yüzeyine çıkışını yansıtır. Bu da plasma membranı yağ asitlerinin önemli derecede açığa
51
çıktığını göstermektedir. 1616 ve 1514 cm-1 IR sinyalindeki azalma (C-C bağları) büyük olasılıkla karbonhidratlar, şeker türevleri ve sialik asitlerin eksikliği nedeniyle olmaktadır. Genz tedavisi sonrası negatif yüklü sialik asitlerin miktarındaki belirgin azalma nedeniyle negatif yüklü COO- moleküler grup sinyalleri azalmaktadır (1616-1576 ve 1393 cm-1).
Çalışmamızda karbonhidrat (şeker türevleri) sinyalleri azalıyorken (1150-950 cm-1 deki glukozidik bağların C-O gerilme ve C-O-H eğilme titreşimleri) [25] [36] aynı zamanda sialik asit sinyallerinin de azaldığı gösterilmiştir. (1118, 1044 ve 1011 cm-1) [25].
Triptolide uygulandığında, prostat kanser hücre hattı proteinlerinin düzenli yapıları azalmaktadır. Amide I ve Amide II bantlarının negatif absorbans olması: protein α-helix yapıları 1659 cm-1de azalırken beta yapraklar 1629 cm-1 de artmış. Bu durum tedavi sonrasında, hücrede apoptosis meydana geldiğini göstermektedir [37].
Triptolide uygulaması nedeniyle, lipit sinyallerinde artış gözlenmektedir. Lipit sinyalleri sırasıyla 3013 cm-1 (doymamış=CH), 2922 ve 2853 cm-1 (doymuş=CH2), 1464 cm-1 (CH2 eğilme modu) ve 1741 cm-1’de (ester C=O) artıyor. Bu spektrum dizisi, Triptolide uygulanmasıyla oluşan apoptozise bağlı plazma membran kabarcıklanması (blebbing) nedeniyle ayrıca gangliozidlerin, şeker türevlerinin (oligosakkaritler, monosakkaritler) ve sialik asitlerin azalmasına bağlı olarak da membran lipitlerinin yüzeye çıktığını göstermektedir. Büyük olasılıkla negatif yüklü sialik asitlerin ve amino asitlerin miktarındaki belirgin azalma nedeniyle negatif yüklü COO- moleküler grup sinyalleri azalmaktadır (1595-1576 ve 1415-1397 cm-1). 1250-1200 cm-1 arasındaki spektral bölgede DNA’nın fosfat grupları (fosfodiester vibrasyonlar) pik vermekte olup, bu sinyaldeki azalma nükleik asitlerden gelen IR sinyallerinin azaldığını gösterir. Bu durum, apoptozise bağlı DNA parçalanmasını göstermektedir [38] [39]. Bununla birlikte, karbonhidrat (şeker türevleri) sinyalleri 1180-900 cm-1 de azalmaktadır [25] ve 1118, 1044 ve 1011 cm-1 de de sialik asit sinyalleri azalmaktadır [25]. Triptolide uygulanmasıyla apoptosis gerçekleştiği için değişen lipit ve protein sinyalleri diğer biyomoleküllerden gelen sinyalleri muhtemelen baskılamaktadır.
52
Çalışmaların gösterdiği sonuçlara göre ganglizoid ailesinin farklı üyelerinin farklı kanser türlerinde EMT, apoptoz, metastaz ve invazyon gibi tümör oluşumu ve yayılmasındaki süreçlerde farklı davrandığı sonucu çıkarılabilir.
53 5. BÖLÜM 5.1. SONUÇLAR:
Çalışmamızda Triptolide’in IC50 değerine ulaşılmış ancak Genz-123346’nın IC50 değeri bulunamamıştır. GM3 ve GD3 sentezi bloke edilerek KDN boyaması yapılmış ve sonuçlar FTIR ölçümleriyle desteklenmiştir. GCS enzimi Genz-123346 ile bloke edilmiş GM3 ve türevi GD3 miktarı azalmıştır. Yine GD3S enzimi Triptolide ile bloke edilmiş ve yalnızca GD3 azalmıştır. Kullanılan ilaçların etkinliği bu şekilde gösterilmiştir. Triptolide’in bir kanser ilacı olması ve hücre üzerine apoptotik etki göstermesi nedeniyle IC50 değerlerine kolaylıkla ulaşılmış ve Muse Viabilite çalışmasıyla 72 saat 60 nM olarak belirlenmiştir. Genz-123346’nın ise hücre üzerine toksik etkisi olmadığı için IC50 değerine ulaşılamamış olup literatür temel alınarak 24 saat süreyle 1000 nM olarak hücreler muamele edilmiştir.
FTIR ile yapılan ölçümlerde hem ilgili ilaçlar ile ilaçlı medyumlar hem de ilaçlı ve ilaçsız hücre hatları karbon bağları üzerinden ölçülmüştür. Triptolide’in apoptoz yaptığı hücresel proteinlerin azalması, DNA hasarı ve lipit artışıyla ortaya konmuş ve böylece Muse ile elde ettiğimiz sonuçların sağlaması yapılmıştır. Yine FTIR sayesinde Genz-123346 ile yaptığımız çalışmanın hücresel protein yapılarını bozmadığı gösterilerek Muse verilerinin sonuçları kontrol edilmiştir.
Çalışmamızda, gerek yaptığımız boyamada gerekse FTIR sonuçlarında GCS enziminin bloke edilmesiyle bütün gangliozid serisininde anlamlı azalma olmasına rağmen KDN’de azalma tespit edilmemiştir. Bu sonuçlarla çalışmanın hedefi olan KDN’nin GM3 üzerinde gösterilmesi gerçekleştirilmemiş olup ancak başka bir tartışmaya yol açmıştır. KDN sadece GM3 üzerinde bulunmayıp hücresel membranlar üzerinde başkaca yapılara bağlı veya serbest halde bulunabilir. Bu çalışmadan KDN’nin GM3 üzerinde bulunmadığı sonucuna varılamaz ancak tek bulunduğu yerin yalnızca GM3 olmadığı sonucu çıkarılabilir. KDN’nin fonksiyonu, yapısı ve karsinogenezisdeki rolünün anlaşılabilmesi için konuyla ilgili daha birçok ileri çalışmaya ihtiyaç vardır.
54
6. KAYNAKLAR 6.1. KAYNAKLAR
1. Abraham L Kierszenbaum, Histology and Cell Biology.: Palme, 2006.
2. McGraw-Hill Global Education Holdings. LLC. (2017) Access Medicine. http://accessmedicine.mhmedical.com/content.aspx?bookid=574§ionid=42 524607
3. American Urological Association. (2017, Haziran) American Urological Association. http://www.auanet.org/education/educational-programs/e-
learning/pathology-for-urologists/normal-histology-and-important-histo-anatomic-structures/prostate
4. The American Cancer Society. (2017, Mayıs) American Cancer Society. https://www.cancer.org/cancer/prostate-cancer.html
5. ATCC. (2016, Haziran) ATCC:The Global Bioresource Center.
https://www.lgcstandards-atcc.org/products/all/HTB-81.aspx?geo_country=tr#documentation 6. Lalita Sharma. (2015, Haziran) Researchgate.
https://www.researchgate.net/post/what_is_the_morphology_of_DU145_cells 7. Venkata Lokesh Battula, Yuexi Shi, Kurt W. Evans et al, "Ganglioside GD2
identifies breast cancer stem cells and promotes tumorigenesis," J Clin Invest, pp. 122(6): 2066–2078, 2012.
8. Tapasree Roy Sarkar, Venkata Lokesh Battula, Steven J Werden et al, "GD3 synthase regulates epithelial-mesenchymal transition and metastasis in breast cancer," Oncogene, pp. 34(23):2958-67, 2015.
9. Adharsh P. Ponnapakam, Jiawang Liu, Kaustubh N. Bhinge et al, "3-Ketone-4,6-diene ceramide analogs exclusively induce apoptosis in chemo-resistant cancer cells," Bioorg Med Chem, pp. 22(4): 1412–1420, 2014.
10. Tae-Wook Chung, Hee-Jung Choi, Seok-Jo Kim et al, "The Ganglioside GM3 Is Associated with Cisplatin-Induced Apoptosis in Human Colon Cancer Cells," PLoS One, pp. 9(5): e92786, 2014.
55
11. Michael M Cox, David L Nelson, "Lipids," in Lehninger Principles of Biochemistry.: WH Freeman, 2005, p. 357.
12. Robert W Ledeen and Gusheng Wu, "GM1 ganglioside: another nuclear lipid that modulates nuclear calcium. GM1 potentiates the nuclear sodium-calcium exchanger.," Can J Physiol Pharmacol, pp. 84(3-4):393-402., 2006.
13. Jeanine R Utz, Thomas Crutcher, Joseph Schneider et al, "Biomarkers of Central Nervous System Inflammation in Infantile and Juvenile Gangliosidoses," Mol Genet Metab, pp. 114(2):274–280, 2016.
14. Donald Voet, Judith Voet and Charlotte Pratt "Structure of GM1, GM2, GM3," in Principles of Biochemistry, 3rd Edition., 2012, p. 253.
15. Hany Farwanah and Thomas Kolter. (2012) MDPI. http://www.mdpi.com/2218-1989/2/1/134/htm
16. Ute Krengel and Paula A. Bousquet "Molecular Recognition of Gangliosides and Their Potential for Cancer Immunotherapies," Front Immunol, pp. 5: 325, 2014. 17. Yong-Yu Liu, Ronald A. Hill and Yu-Teh Li "Ceramide Glycosylation Catalyzed
by Glucosylceramide Synthase and Cancer Drug Resistance," in Advances In Cancer Research V.117 The Role of Sphingolipids in Cancer Development and Therapy.: Academic Press, 2013, pp. 70-79.
18. Daita Nadano, Mariko Iwasaki, Satoshi Endo et al, "A naturally occurring deaminated neuraminic acid, 3-deoxy-D-glycero-D-galacto-nonulosonic acid (KDN). Its unique occurrence at the nonreducing ends of oligosialyl chains in polysialoglycoprotein of rainbow trout eggs.," J Biol Chem, pp. 261(25):11550-7., 1986.
19. Sadako Inoue and Ken Kitajima, "KDN (Deaminated neuraminic acid): Dreamful past and exciting future of the newest member of the sialic acid family," Glycoconjugate Journal, pp. 23(5): 277–290, July 2006.
20. Paola Giussani, Cristina Tringali, Laura Riboni et al, "Sphingolipids: Key Regulators of Apoptosis and Pivotal Players in Cancer Drug Resistance," Int J Mol Sci, pp. 4356-4392, 2014. [Online]. http://www.mdpi.com/1422-0067/15/3/4356/htm
56
21. Song Yu, Naoya Kojima, Sen-itiroh Hakomori et al, "Binding of rainbow trout sperm to egg is mediated by strong carbohydrate-to-carbohydrate interaction between (KDN) GM3 (deaminated neuraminyl ganglioside) and Gg3-like epitope," Cell Biology, pp. 99(5): 2854–2859, 2002.
22. MekaNews Lite powered by WordPress. (2016, Temmuz) MUSE COUNT VIABILITY KIT USERS GUIDE. http://circabook.com/muse-count-viability-kit-users-guide/
23. (2017, Haziran) SlideShare. https://image.slidesharecdn.com/irspectroscopy-140210002609-phpapp01/95/dispersive-ftir-22-638.jpg?cb=1391992050
24. Peter R Griffiths and James A De Haseth, Fourier Transform Infrared
Spectrometry., 2007. https://en.wikipedia.org/wiki/Fourier_transform_infrared_spectroscopy
25. Lewis AT, Jones K, Lewis KE et al, "Detection of Lewis antigen structural change by FTIR spectroscopy," Carbohydr Polym, pp. 15;92(2):1294-301, 2013. 26. Allison Derenne, Magali Verdoncka and Erik Goormaghtigh, "The effect of
anticancer drugs on seven cell lines monitored by FTIR spectroscopy," Analyst, no. 137(14):3255-64, 2012.
27. Allison Derenne, Vincent Van Hemelryck, Delphine Lamoral-Theys et al, "FTIR spectroscopy: A new valuable tool to classify the effects of polyphenolic compounds on cancer cells," Biochim Biophys Acta, no. 1832(1):46-56, 2013. 28. Alix Mignolet, Allison Derenne, Margarita Smolina et al, "FTIR spectral
signature of anticancer drugs. Can drug mode of action?," Biochimica et Biophysica Acta, 2015.08.010.
29. Saroj Kumar, Thankaraj Salammal Shabi and Erik Goormaghtigh, "A FTIR Imaging Characterization of Fibroblasts Stimulated by Various Breast Cancer Cell Lines," PLoS One, no. 9(11), 2014.
30. A Salman, RK Sahu, E Bernshtain et al, "Probing cell proliferation in the human colon using vibrational spectroscopy: a novel use of FTIR-microspectroscopy," Vibrational spectroscopy, no. 34, 2004.
57
31. Mourant JR,Yamada YR, Carpenter S et al, "FTIR spectroscopy demonstrates biochemical differences in mammalian cell cultures at different growth stages," Biophys J, no. 85(3):1938-47, 2003.
32. Thomas A Natoli, Laurie A Smith, Kelly A Rogers et al, "Inhibition of glucosylceramide accumulation results in effective blockade of polycystic kidney disease in mouse models.," Nat Med, pp. 16(7): 788-92, 2010.
33. Jozsef Karman, Jennifer L Tedstone, Nathan K Gumlaw et al, "Reducing glycosphingolipid biosynthesis in airway cells partially ameliorates disease manifestations in a mouse model of asthma.," Int Immunol, pp. 22(7):593-603, 2010.
34. Haw-Young Kwon, Seok-Jo Kim, Cheorl-Ho Kim et al, "Triptolide downregulates human GD3 synthase (hST8Sia I) gene expression in SK-MEL-2 human melanoma cells," Exp Mol Med, pp. 42(12): 849–855, 2010.
35. Mark Slevin, B Costello, S Kumar and J Gaffney, "The ganglioside GM(3) is raised in the sera and tissue of patients with bladder tumours," Int J Oncol, pp. 8(2):271-4, 1996.
36. Marta Kačuráková and Mohamed Mathlouthi, "FTIR and laser-Raman spectra of oligosaccharides in water: characterization of the glycosidic bond," Carbohydr Res, pp. 284(2):145-57, 1996.
37. Karlien Le Roux, Linda C Prinsloo and Debra Meyer, "Fourier Transform Infrared spectroscopy discloses different types of cell death in flow cytometrically sorted cells," Toxicol In Vitro, pp. 29(7):1932-40, 2015.
38. Yanfeng Gao, Xiongwei Huo, Liu Dong et al, "Fourier transform infrared microspectroscopy monitoring of 5-fluorouracil-induced apoptosis in SW620 colon cancer cells," Mol Med Rep, pp. 11(4): 2585–2591, 2015.
39. .
Plaimee P, Weerapreeyakul N, Thumanu K et al, "Melatonin induces apoptosis through biomolecular changes, in SK-LU-1 human lung adenocarcinoma cells," Cell Prolif, pp. 47(6):564-77, 2014.
![Şekil 1. Prostat [2].](https://thumb-eu.123doks.com/thumbv2/9libnet/3036968.2657/11.892.180.774.645.1001/sekil-prostat.webp)
![Şekil 3. Du-145 Prostat Kanser Hücre Hattı hücrelerinin kontrast mikroskop altındaki görüntüsü [6]](https://thumb-eu.123doks.com/thumbv2/9libnet/3036968.2657/14.892.172.787.126.591/sekil-prostat-kanser-hucrelerinin-kontrast-mikroskop-altindaki-goruntusu.webp)
![Şekil 5. GD3’ün biyokimyasal yapısı [15].](https://thumb-eu.123doks.com/thumbv2/9libnet/3036968.2657/16.892.196.767.141.643/sekil-gd-un-biyokimyasal-yapisi.webp)
![Şekil 6. Seramidden Gangliozid sentezinin basamakları [20].](https://thumb-eu.123doks.com/thumbv2/9libnet/3036968.2657/19.892.175.800.134.461/sekil-seramidden-gangliozid-sentezinin-basamaklari.webp)

![Şekil 7. Muse™ Cell Analyzer cihazında % canlılık değerlerinin belirlenmesi [22].](https://thumb-eu.123doks.com/thumbv2/9libnet/3036968.2657/26.892.287.671.124.460/sekil-muse-cell-analyzer-cihazinda-canlilik-degerlerinin-belirlenmesi.webp)
![Şekil 8. Fourier Transform Infrared Spectroscopy Assay (FTIR) [23]](https://thumb-eu.123doks.com/thumbv2/9libnet/3036968.2657/27.892.238.737.623.902/sekil-fourier-transform-infrared-spectroscopy-assay-ftir.webp)


