Carbonic anhydrase inhibitors: Inhibition of the β-class enzyme from the yeast Saccharomyces cerevisiae with sulfonamides and sulfamates
Tam metin
Şekil

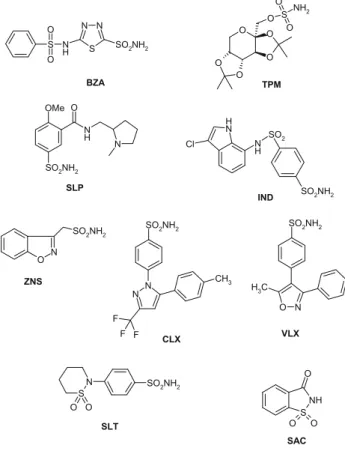
Benzer Belgeler
We thus propose such a model to investigate the optimal quality improvement path for a company given that quality costs depend on both autonomous and induced types of
Biyodizelin en genel tanımı; hayvansal ve bitkisel atıkların eklenmesiyle üretilen dizel yakıttır. Bilimsel olarak tanımlanırsa; organik olan yağların baz ve alkol ile
Alanda tespit edilen Boz ayılar (a: Üretim faaliyeti öncesi, b: Üretim faaliyeti sonrası, c, d: beslenme amaçlı alanı kullanması).. Alanı sonbaharda daha yoğun
Methods For 70 normal eyes of individuals with varying age, the ETDRS chart acuity was determined as well as discriminated target perimetry using high-contrast targets presented for
32-Kullanmış olduğum telefonu kamera özelliği için tercih ettim 0,000 33-Kullanmış olduğum telefonun mp3 çalar özelliğinden dolayı tercih ettim 0,000 34-Kullanmış
The effects of treatment temperature and time on mass loss, wood density, chemical changes, and decay resistance were evaluated using an ANOVA and Duncan’s comparison test..
Örnek alanlarda yol platformu, yol dışı (yol altı ve yol üstü) ve kontrol noktalarında tespit edilen yaban hayvanı görüntü sayısı arasında yapılan varyans
We find that both an active consumer and a market were forming within the budding sixteenth- and seventeenth-century Ottoman con- sumer culture, through the interactions of
