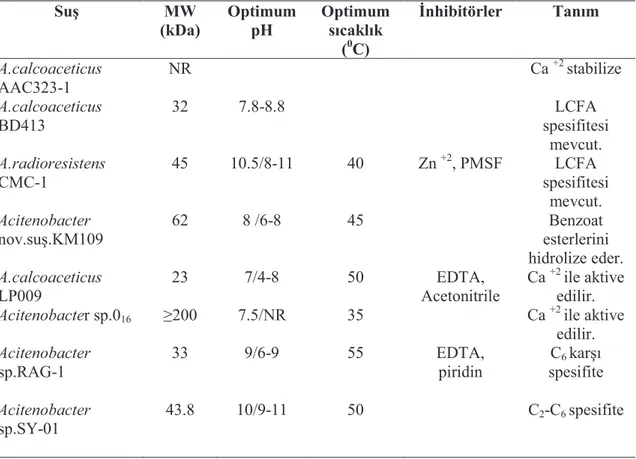
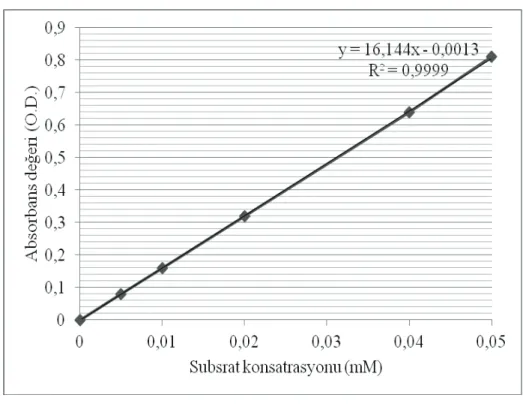
Acinetobacter psychrotolerans Suşlarından İzole Edilen Lipazın Karakterizasyonları
Tam metin
Şekil








Benzer Belgeler
Sonuç olarak; bu çalışma tigesiklinin A.baumannii planktonik hücreleri üzerinde güç- lü aktivitesinin olduğunu ve bu etkinin bakterinin biyofilm oluşturma özelliğinden
With 65 % of respondents understand digital preservation and copyright, 74% have experience on repository platforms, 83.3% can collect, store, and preserve research output of
Also, we characterize some of its properties. Later, we define the vertex and edge chromatic number BF- Complete Bipartite graph. Further we illustrated an example for BFRGS
ÖZ Amaç: Bu çalışmada, yoğun bakım ünitesi (YBÜ) örneklerinden izole edilen Pseudomonas aeruginosa ve Acinetobacter baumannii suşlarının üç yıllık
Kolistin heterodirenci, MİK değeri <=2 µg/ml olan bir izolat için, MİK >2 µg/ml kolistin varlığında saptanabilir alt popülasyonların üreyebilmesi olarak
Çok uzun bir süre toprak ve çevreden izole edilen ve neredeyse avirulan bakteri olarak bilinen Acinetobacter, bu nedenle klinik örnekler- den izole edildiğinde uzun süre
Haziran 2011-Haziran 2013 tarihleri arasında 400 yataklı Kahramanmaraş Necip Fazıl Şehir Hastanesi Mikrobiyoloji Laboratuvarı’na tüm klinik birimlerden
Yoğun bakım ünitesi ve diğer ünitelerde yatan hastalardan izole edilen Acinetobacter baumannii suşlarında in-vitro anti- biyotik direnci, ANKEM Derg
