Mdamb-231 meme kanseri hücre dizisinde prostaglandin endoperoksid H sentaz 2 (PTGS2), kalretikulin (CALR) ve keratin-19 (KRT19) genlerinin transkripsiyon aşamasında anlatımlarının araştırılması
Tam metin
Şekil



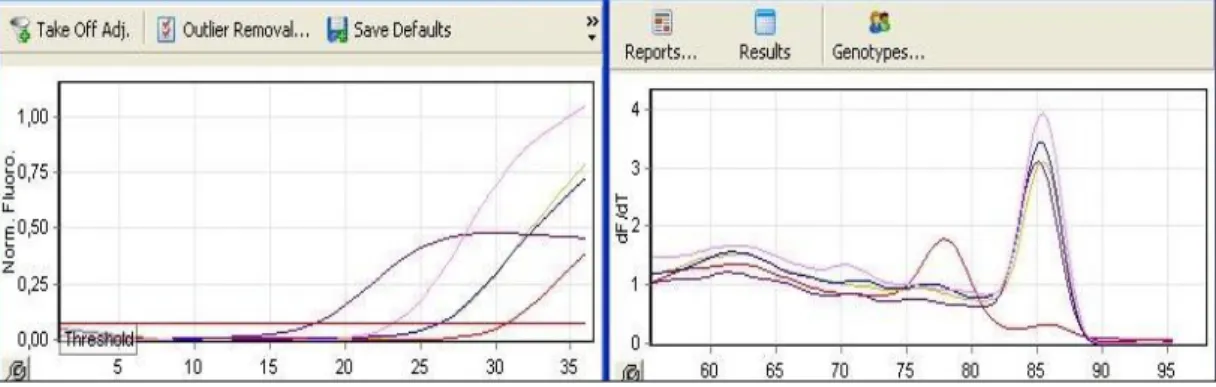
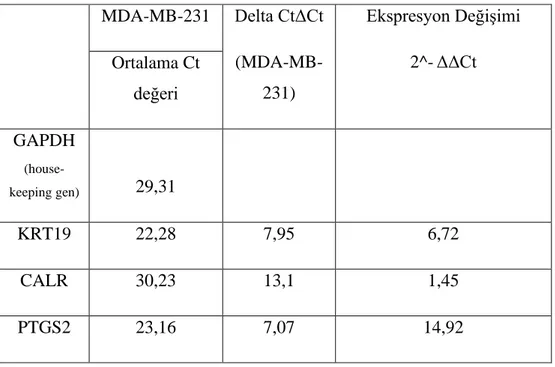
Benzer Belgeler
Ancak yukarıda bulgular kısmında da görüldüğü gibi öğretmenler kendilerine harita aracılığı ile öğrencilerinde daha dengeli bir dünya algısı
(A) No caspase-3 expression in control group, (B) Increased expressions in Purkinje cells (arrows) in LPS group, (C) Decreased immunoreaction in Purkinje cells (arrow) PG treated
The literature has additio- nally emphasized perceived pay to possibly be an important link in the relationship between work demands and negative employee behaviors (DeConinck
In the present study, we have investigated the dynamics of parasite in- fection over the course of a disease season following vaccination with a commercial cell line vaccine,
Kemik geliþiminin geri olmasý, rizomelik kýsalýk, küçük el ve ayak parmaklarý, belirgin alýn, basýk burun kökü, büyüme geliþme geriliði ve hipotoni bulgularý
Hasat dönemlerine göre Iris rizomlarının uçucu yağ verimi, kuru rizom verimlerindeki artışa bağlı olarak artmış ve hasat sonrası rizomlarda 0,123-0,300 L/da
Ayrıca n kodun uzunluğu, m otomorfizmanın mertebesi olmak üzere (n,m)=1 olması durumunda skew cyclic kodun cyclic koda eşit, (n,m)=d olması durumunda quasi-
Bu çalışmada; Afyon Kocatepe Üniversitesi Tıp Fakültesi Hastanesi’nde çalışan hemşirelerin iş tatmin düzeylerini etkileyen faktörler, motive edici faktörler
