Contents lists available atScienceDirect
Industrial Crops & Products
journal homepage:www.elsevier.com/locate/indcropLC-MS, NMR
fingerprint of Potentilla argentea and Potentilla recta extracts
and their in vitro biopharmaceutical assessment
Stefania Sut
a, Stefano Dall
’Acqua
b, Sengul Uysal
c,⁎, Gokhan Zengin
c, Abdurrahman Aktumsek
c,
Caren Picot-Allain
d, Fawzi Mahomoodally
daDAFNAE, Department of Agronomy, Food, Natural Resources, Animals and Environment, Agripolis Campus, University of Padova, 35020 Legnaro PD, Italy bDepartment of Pharmaceutical and Pharmacological Sciences, University of Padova, Via Marzolo 5, 35131 Padova, Italy
cDepartment of Biology, Science Faculty, Selcuk University, Campus, Konya, Turkey dDepartment of Health Sciences, Faculty of Science, University of Mauritius, Réduit, Mauritius
A R T I C L E I N F O Keywords: Potentilla Ellagitannis Triterpenoids Antioxidant Enzymes Phyto-pharmaceutics A B S T R A C T
Many Potentilla species are used as traditional drugs in several countries and also studied for their possible therapeutic properties. As part of our continuous effort to shed light on the biopharmaceutical potential of such plant species, this study was aimed to establish the phytochemicalfingerprinting and to evaluate biopharma-ceutical potential of methanol, ethyl acetate, and water extracts obtained from the aerial parts of Potentilla recta L. and P. argentea L. Combination of 1 dimension, 2 dimension-nuclear magnetic resonance as well as LC-DAD-ESI-MS and LC-APCI-MS allowed the identification of ellagitannins, flavonol glycosides, and triterpene con-stituents. To quantify different constituents in the extracts obtained with different solvents, the total contents in phenolics,flavonoids, saponins, and triterpenoid were evaluated. The antioxidant activity was assessed by the means of metal chelation, phosphomolybdenum, radical quenching (by 2,2-diphenyl-1-picrylhydrazyl [DPPH] and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)[ABTS]) and reduction ability (by cupric reducing antioxidant capacity [CUPRAC] and ferric reducing antioxidant power [FRAP]) assays. Phytochemical analysis showed similar composition of the tested methanol extracts. Acetylcholinesterase (AChE), butyrylcholinesterase (BChE), tyrosinase,α-amylase, and α-glucosidase were selected as target enzymes to detect enzyme inhibitory potentials of these extracts. The water extract of P. argentea (113.29 mg gallic acid equivalent/g extract) con-tained the highest phenolic amount while the methanol extract (519.70 mg quillaja equivalent /g extract) showed the highest saponin content. The water extract of both Potentilla species was the most potent ABTS scavenger as well as present the highest CUPRAC activity. The most potent inhibitory activity against AChE and BChE was observed for the ethyl acetate extracts of P. recta and P. argentea. The methanol extracts of P. recta (53.16 mmol acarbose equivalent [ACAE]/g extract) and P. argentea (45.68 mmol ACAE/g extract) were the most active inhibitors ofα-glucosidase. Data collected from this study support the multiple therapeutic actions of P. recta and P. argentea and justify further pharmacological investigations for potential applications in the phar-maceutical industry.
1. Introduction
The role of plants as therapeutic tool is a topic of global importance. Indeed, the therapeutic virtues of plants have been used by man since time immemorial. Secondary metabolites, ubiquitously present in plants, possess tremendous biological potential (Zengin et al., 2019). Substantial amount scientific reports have brought the multiple phar-macological applications of plant derived compounds and plant extracts into the limelight. Harnessing the biological potential of plants re-presents a sterling opportunity for the development of novel
therapeutic candidates.
The genus Potentilla (Rosaceae), mainly distributed in temperate regions of the Northern hemisphere, comprises of 300 species of an-nual/biennial herbaceous plants and subshrubs (Grochowski et al., 2017; Şöhretoğlu et al., 2018). Also known as cinquefoils, Potentilla species are well known among the folk community for their diabetic, antidiarrheal, antiviral, inflammatory, anticancer, anti-ulcerogenic, and wound healing effects, making species from this genus interesting candidates for the development of novel pharmaceuticals (Şöhretoğlu et al., 2017;Tomczyk, 2006). Sulphur cinquefoil Potentilla
https://doi.org/10.1016/j.indcrop.2019.01.047
Received 24 December 2018; Received in revised form 17 January 2019; Accepted 23 January 2019
⁎Corresponding author.
E-mail address:[email protected](S. Uysal).
Available online 28 January 2019
0926-6690/ © 2019 Elsevier B.V. All rights reserved.
recta, is a long-lived invasive herbaceous perennial plant native to the eastern Mediterranean region of Eurasia (DiTomaso and Kyser, 2013). In folk medicine, P. recta is used for several purposes as a sytptic, anti-inflammatory or tonic agent (Bazylko et al., 2013). Pharmacological studies have focused on the antioxidant, antimicrobial activities of P. recta, as well as the lipoxidase, tyrosinase, cholinesterases, amylase, glucosidase, and hyaluronidase inhibitory activity of the plant (Bazylko et al., 2013;Tomczyk et al., 2008;Uysal et al., 2019a). P. argentea, also known as, silvery or hoary cinquefoil, has leaves which are silvery-white tomentose on the underside (Stroh, 2014). Several pharmacolo-gical investigations have reported the anti-cancer, antibacterial, and antioxidative activities of compounds isolated from P. argentea (Tomczyk and Latté, 2009). Previous investigation on Potentilla species revealed the presence of several classes of secondary metabolites as triterpene, procyanidins, andflavonoids (Xue et al., 2006).
Potentilla species in the Turkish traditional medicine are highly praised and used which ten to justify the need for more accurate in-vestigation of the chemical constituents and the possible biological targets. The aerial parts of two species, P. recta L. and P. argentea L., were explored for their potential bioactivity using different in vitro and enzymatic assays correlating the observed effects with the chemical composition offering new information about these medicinal species. Specifically, the enzyme inhibitory and antioxidant potentials of P. recta and P. argentea aerial parts extracted by different solvents extracts were evaluated. NMR and LC-ESI-DAD-MS analysis were performed for chemical characterization of plant materials.
2. Materials and methods
2.1. Plant materials and extraction procedure
The Potentilla species were collected at July 2015 in Turkey (P. ar-gentea: Bursa, Uludag, Süleymaniye village; P. recta: Konya, Taskent district). The botanist Dr. Murad Aydın Sanda (Mus Alpaslan University, Mus, Turkey) confirmed taxonomically the plant species and one voucher specimen was kept at the herbarium of Selçuk University, Konya, Turkey. The aerial parts were used and these parts dried for 10 days at the room temperature. After this period, the dried samples were powdered by using a laboratory mill.
Soxhlet technique was selected for obtaining ethyl acetate and methanol extracts (5 g plants were extracted with these solvents for 6–8 h in a Soxhlet apparatus). The extracts were filtered and then concentrated using a rotary evaporator under vacuum at 40 °C. Infusion was prepared as water extract (5 g of plants were kept in 100 ml of boiled water for 20 min). The water extract wasfiltered and then dried by using a lyophilizator. The extracts were stored at + 4 °C until further analysis. The extraction yields for these solvents are given inTable 1. 2.2. Phytochemical profiles
With reference to our previous studies (Uysal et al., 2017), the total amount of phenolics (TPC) (by standard Folin-Ciocalteu method),
flavonoids (TFC) (by AlCl3method), triterpenoids (TTC)
(vanillin-gla-cial acetic acid method), and saponins (TSC) (vanillin-sulfuric acid method) were determined. Standard compounds (gallic acid (mg GAE/ g) for TPC; rutin (mg RE/g) for TFC; Oleanolic acid (mg OAE/g) for TTC and Quillaja (mg QE/g) for TSC, respectively) were used to express the results.
The 1D- and 2D- NMR spectra were obtained using a Bruker Avance III 400 Ultrashield spectrometer 400.11 MHz. The NMR spectra were acquired in MeOD-d4(Sigma-Aldrich) with TMS as an internal standard.
Durian® 4.95 mm NMR tubes (Durian Group) were used filled with 750μL of extract solution. Chemical shifts were calculated in δ values.
1H-NMR and HSQC-DEPT, HMBC, COSY experiments were acquired
using standard Bruker sequences.
HPLC-DAD-ESI-MSnanalysis was used for the analysis of phenolic using a Chromatograph Agilent 1260 (Santa Clara, CA, USA) equipped with 1260 autosampler, column oven and diode array detector (DAD). After colunm theflow was separated by “T” connector and half of the liquid was spiltted to an Agilent/Varian MS-500 ion trap mass spec-trometer. Separation was achieved using an Agilent Eclipse XDB C-18 (3.0 × 150 mm, particle size 3.5μm) as stationary phase. The sample injection volume was 10μL. The mobile phase was 0.1% formic acid in water (A) and acetonitrile (B). The gradient elution start with 90% A and is changed to 0% of A in30 min,flow rate was 0.5 mL/min. The DAD detector was used to estimate the amount offlavonoids and ellagic acid derivatives. Rutin (Sigma Aldrich, St. Louis, MO, USA) and ellagic acid (Sigma Aldrich) were used as reference compounds. The chroma-tograms were monitored atλ 280 nm (gallic acid derivatives) and λ 350 nm (flavonol derivatives) and UV–vis spectra were acquired in the range of 200–400 nm. MS spectra were acquired in the m/z 50–2000 range, using ESI ion source operating in negative ion mode. Fragmentation of the ionic species was obtained using the turbo data dependent scanning (TDDS) instrument function. The analysis of frag-mentation spectra, the comparison with the literature and reference compounds allowed the identification of constituents. Quantification of was obtained with the method of calibration curve: rutin was used as external standard forflavonoid quantification, ellagic acid was used for its derivatives, chlorogenic acid for caffeic acid derivative. Calibration curves were as follows y = 51.33x + 3.11 (R2= 0.9987) for rutin;
y = 26.22x + 0.98 (R2= 0.9978) for ellagic acid, chlorogenic acid
y = 47,359x + 439,99 (r² = 0,9951);.
For the triterpene analysis the method previously described for apple triterpene was used (Sut et al., 2018a). HPLC-APCI-MSnin
ne-gative ion mode was used. Triterpenes were separated on an Agilent Eclipse XDB C-18 (3.0 × 150 mm) 3.5μm using methanol (A) and H2O (B) as mobile phases. A gradient program was used as follows: [0→ 12th min: A:B (45:55)→ A:B (80:20) 12 → 48th min: A:B (80:20) → A:B (80:20) 48→ 49th min: A:B(80:20) → A:B (45:55) 49→55th min: A:B (45:55) → A:B (45:55)]. Flow rate was 500 μL/min. Calibration curve was obtained using oleanolic acid and in the range 1–50 μg/mL. The calibration curve was y = 23,3214x + 2,789 (r² = 0,9981).
Table 1
Extraction yields and total bioactive components of the tested extracts*.
Plants-Solvents Extraction yields (%)
Total phenolic content (mg GAE/g extract)
Totalflavonoid content (mg RE/ g extract)
Total saponins (mg QE/g extract)
Total triterpenoids (mg OAE/g extract) P. argentea-EA 3.75 17.44 ± 0.76e 41.53 ± 0.39c 432.30 ± 36.80b 3.30 ± 0.03c P. argentea-MeOH 18.99 89.81 ± 0.79c 63.77 ± 1.08a 519.70 ± 29.36a 3.45 ± 0.02b P. argentea-Water 6.46 113.29 ± 1.51a 32.95 ± 0.52d 238.87 ± 16.52d 0.70 ± 0.01e P. recta-EA 2.40 28.50 ± 1.00d 28.20 ± 0.22e 358.41 ± 27.51c 3.78 ± 0.07a P. recta-MeOH 17.43 93.00 ± 1.26c 54.26 ± 0.21b 337.12 ± 28.78c 2.99 ± 0.04d P. recta-Water 11.10 96.51 ± 2.96b 24.91 ± 0.08f 88.22 ± 17.07e 0.19 ± 0.01f
* Values expressed are means ± S.D. of three parallel measurements. GAE: Gallic acid equivalent; RE: Rutin equivalent; QE: Quillaja equivalent; OAE: Oleanolic acid equivalent. EA: Ethyl acetate. MeOH: Methanol. Different letters indicate significant differences in the extracts (p < 0.05).
2.3. Assays for enzyme inhibition and antioxidant capacity
Tyrosinase,α-amylase, α-glucosidase, and cholinesterases were se-lected as target enzyme and the procedures of these assays were de-scribed in our earlier work (Mollica et al., 2017;Uysal et al., 2017). Standard inhibitors (acarbose (for α-amylase and α-glucosidase), ga-lantamine (for AChE and BChE), and kojic acid (for tyrosinase)) were used to express the enzyme inhibitor properties.
Antioxidant capacity of the extracts were spectrophotometrically screened by different experiments as ferrozine assay (for chelating abilities). Phosphomolybdenum, reduction potentials (by FRAP and CUPRAC assays) and radical attenuation (using DPPH and ABTS radi-cals). Standard compounds (TE/g and EDTAE/g) were used to express the antioxidant properties the procedures of assays were given reported in our earlier work (Uysal et al., 2017).
2.4. Data evaluation
The obtained results (mean ± standard deviation (SD)) were sta-tistically evaluated and for this One-way ANOVA (by Tukey test, p < 0.05) was done to indicate differences among the tested extracts. To further statistical evaluation, Pearson correlation, heat map, Principal component (PCA) and Sparse Partial Least Squares (sPLS-DA) analysis were carried out to observe variabilities of the tested extracts. The statistical procedures were performed by R software v. 3.5.1. 3. Results and discussion
3.1. Chemical profiles
The methanolic extracts of aerial parts of two Potentilla species were prepared to compare the phytochemical composition. This approach allow, due to the solvent properties of methanol, the extraction of the secondary metabolites from the plant extracts. As stating point, NMR experiments were acquired. Furthermore LC-DAD-and LC–MS analysis were also obtained. The purpose of this phytochemical screening is to assess the presence of different classes of phytoconstituents. 1D and 2D-NMR experiments were used in order to obtain information about the structure of the chemical constituents of the extracts (Mahrous and Farag, 2015). NMR approach present advantages due to the absence of any chromatographic separation and on the fact that all different phytochemicals containing proton atoms will have the same response factor. The signal superimposition due to the presence of several deri-vatives sharing similar structures can be in part overcome using bidi-mensional experiments and diffusion ordered spectroscopy (Halabalaki et al., 2014;Mahrous and Farag, 2015;Rodrigues et al., 2009). The use of multi technique approach in the characterization of plant extracts can be of advantage allowing the identification of different classes of constituents in plant extracts without the need of isolating pure pro-ducts, using data of both chromatographic and spectroscopic ap-proaches as mass spectrometry and NMR (Fidyt et al., 2016;Rodrigues et al., 2018;Sut et al., 2018b,2019).
The1H NMR of the two extracts presents similarities in particular in the aliphatic region signals supporting the presence of fatty acids can be observed (Table 2,Fig. 1). Assignments of their multiplicity (as CH2)
and values of directly linked carbon was done by HSQC-DEPT (see supplementaryfile) while spin system and long range C–H correlations were obtained due to COSY, TOCSY, and HMBC spectra (supplementary file). Main findings are summarized inTable 2. Fatty acid derivatives also containing double bond have been detected in both extracts. The crowded spectral region between δ 3.0–4.5 suggest the presence of numerous carbohydrate derivatives. Some differences can be observed in the aromatic region δ 6–8 ppm that is characterized for both the species by large number of partially overlapping signals. From the1H
spectra (Fig. 1), it is clear that P. argentea sample present signals that appear to be broader and more superimposed than those observed in P
recta.
Diffusion Ordered Spectroscopy (DOSY) experiments were per-formed to obtain simplified spectra of the mixture on the basis of dif-fusion coefficients (Mahrous and Farag, 2015). This technique has been employed in the analysis of mixtures and to phytochemicals (Halabalaki et al., 2014;Rodrigues et al., 2009). Signals that present similar dif-fusion coefficients can be ascribed to fatty acid with resonances pre-senting diffusion coefficient values ranging from -9.559 and −9.503 log (m2/s). Signals ascribable to sugars were also observed−9.512 log(m2/
s). Projecting row with diffusion coefficient −9.515 log(m2/s) result in
spectrum that revealed the presence of numerous sharp signals in the aliphatic region atδ 1.0–2.0 as well as signals in the region of carbo-hydrate suggesting the possible presence of glycosides. Furthermore, in the aromatic region of the spectra signals in the range ofδ 6–8 support the presence of different compounds characterized in both the Potentilla extracts by group of signals with diffusion coefficients from −9.488 log (m2/s),−9.563 log(m2/s), 9.670 log(m2/s) and 9.651 log(m2/s).
Lim-ited differences were observed from the two extracts suggesting the presence of similar constituents in the plant materials.
To have further information on the structure of the main compounds in the extracts HSQC-DEPT, HMBC to assess heteronuclear correlations, COSY and 1D and 2D TOCSY were acquired. The HSQC DEPT experi-ment was acquired for both the samples allowing the observation of the chemical shift of the non-quaternary positions and identification of CH, CH3, or CH2. Long range H-C correlation were observed in the HMBC
spectra giving further structural information. Data were merged with COSY, 1D and 2D TOCSY yielding in the identification of some specific classes of constituents as reported inTable 2.
The relative carbon signals ascribed to CH and CH3or CH2were
assigned as presented inTable 2. Diagnostic long range correlation from the CH2 signals with carbon resonances at δ 173–174 and δ 28-26
support the presence of carboxyl function and other aliphatic groups thus suggesting the presence of fatty acid moieties. The aliphatic region of the HSQC-DEPT of the two samples present differences. In P. ar-gentea, the aliphatic region of the HSQC-DEPT spectrum present signals ascribed to CH or CH3 as well as several CH2. The numerous sharp
signals in the aliphatic region ranging fromδ 0.60–1.45 for1H andδ 9–30 for13C and their HMBC correlation are in agreement with NMR
assignments of triterpene correlated to ursolic, oleanolic and tormentic acid derivatives (Seebacher et al., 2003;Silva et al., 2012;Waldbauer et al., 2015) and are reported inTable 2.
Sugar region was studied combining HSQC-DEPT, HMBC COSY, and 1D and 2D TOCSY data. Selective 1D TOCSY allowed to observe the presence of a monosaccharide that present six group of signals the coupling constants and the assignments of HSQC are consistent with β-glucopyranoside moiety.
The sp2region showed the presence of proton signal atδ 5.3 cor-relating directly with carbon at 125–127 ppm assigned to olefinic proton of fatty acid or triterpene. Furthermore, signal at δ 5.42 (d J = 3,5) correlating with carbon resonance atδ 92 ppm and in HMBC with carbons atδ 103 and δ 73 supporting the assignment of sucrose (Table 2). The aromatic region presents for P. argentea numerous signals partially overlapped. The most intense correlations observed in the HSQC indicate the presence of aromatic CH. HSQC-DEPT and HMBC correlations allowed the tentative identification of flavonol moieties ascribable to luteolin kaempferol and isorhamnetin, also supported by the methoxyl signal at 3.5 ppm (see Table 2), signals ascribable to glycosidic part of the compounds are revealed as anomeric signals in the regionδ 4.5-5-5. Thus, both the extracts contain flavolol glycosides. Signals ascribable to esterified glucopyranosil moieties can be observed in the rangeδ 4.3–6.40 presenting HMBC correlations with carboxyl functions atδ 168-170. This, in relation with the presence of phenolic moieties, support the presence of polymeric structures containing gal-loyl groups (Li et al., 1999;Vrhovsek et al., 2012). Thus the two extract present similar composition with the presence of ellagitannin structures ascribable to Potentillin or Agrimonin (Vrhovsek et al., 2012),
flavonoids, the P. recta also present more evident the signals ascribable to triterpene derivatives in the aliphatic region.
HPLC-DAD-ESI-MS analysis of P. argentea and P. recta in negative ion mode allowed to compare the composition of phenolics. The chromatograms at 270 nm, shows that the phytochemical composition of the two methanol extracts was comparable (Fig. 2). Intense peaks presenting characteristic UV spectra with absorption between 250 and 360 nm were detected at 19.4, 19.8, 20.2, 20.8 min suggesting the presence of ellagic acid derivatives and related structures.
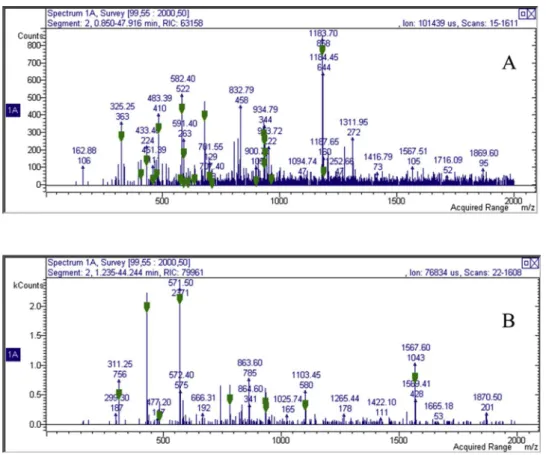
The MS detection allowed to observe the m/z values of the eluted compounds and in P. argentea the LC–MS chromatogram showed nu-merous significant peaks characterized by ionic species in negative ion mode that ranged from m/z 160 to 1200 (Fig. 3). MSnfragmentation
during the HPLC chromatogram run allowed the observation of frag-mentation pathways for several species allowing tentative identification
of compounds that are summarized inTable 3.Flavonoids and ellagi-tannin derivatives was tentatively identify as report inTable 3. Quali quantitative composition of the two extracts resulted to be very similar. The ellagitannin derivatives are resuming the structures of Potentillin on the basis of MS fragmentation (Vrhovsek et al., 2012). LC-APCI-MS was used to perform analysis of triterpene using a method previously published for apple triterpene acids (Sut et al., 2018a). A typical chromatogram is reported (Fig. 4) and the assignments are in-dicated in Table 3. The two samples present the same qualitative composition. Thus the two extracts contain similar levels of triterpene as observed also by spectrophotometric measurements.
The identification of such compounds offer a new information on the composition of these two Potentilla species. The secondary meta-bolites occurring in plants have been studied due to their activities and in several cases they have used to prepare and discovery new drugs that Table 2
NMRfindings in the two Potentilla extracts.
Group and compound δH COSYδH δC (HSQC-DEPT) and multiplicity
δ C HMBC correlations Potentilla species
Fatty acids CH2 0.89 1.35 12.5 -15.0 CH3 A, R 1.30 2.07 28.0 to 36.90 CH2 1.61 2.33 38.1-26.5 CH2 2.05 1.57 38.1; 31.2; 26.3, 126.8; 128.4;28.0 2.10 0.99 24.4 CH2 2.33 1.58 28.7 CH2 174.2; 28.3 2.81 33.3; 28.1 CH2 173.2; 28.2 39.7 CH2 Fatty acid CH sp2 5.31 128.4; 126.8 24.4; 26.3
Tormentic acid/euscaphic acid 0.82 s(CH3-24) – – 27.8 82.3 (C-3); 54.5 (C-5); 16.8 (CH3-23) A 1.03 s(CH3-23) 16.8 82.3 (C-3); 54.5 (C-5); 27.8 (CH3-24) – 54.5-56.2 – 27.3 140.0 (C-13); 27.1 (C-15) 0.86 24.2 42.5 20); 72.7 19); 53.4 (C-18) 1.00 (CH3-27) 53.4 24.1 (CH3-23); 48.2 (C-17); 127.5 2.20 (CH3-28) (C-12); 180.1 (C-30) 82.3 2.52 (CH-18) 3.25 (CH-3) Oleanolic/Ursolic acid 0.84 (CH-5) 54.0 A 0.78(CH3-23) 77.8 (C-3); 54.0 (C-5); 38.0 (C-4); 27.0 (CH3-24) 0.98(CH3-24) 77.8 (C-3) ; 54.0 (C-5); 38.0 (C-4); 16.0 (CH3-23) 5.21 (CH-12) 127.1
Sucrose 5.42 (H-1 glu) d J = 3.4 3.46 91.4 103.8 (C-1fru); 73.3(C-3 glu); A, R Glycosidicflavonol derivatives 6.20 97.2
6.37 105.8
(Kaempferol, Luteolin, Isorhamnetin) 6.63 105.9
7.10 108.7 7.30 127.8 7.50 128.8 β-glucopyranoside 3.84 (OCH3-aromatic) 55.6 4.19 d (J = 7.7) 3.16 102.4 76.5; 73.3; 165.0 4.21 d (J = 7.7)
Ellagitannin derivatives glucose moiety 92.5 168.5
6.40 (H-1) 72.9 168.5
5.34 (H-2) 69.6 170.5
5.24 (H-4) 68.0 169.1
4.30 (H-5) 63.0 166.0
4.94 (H-6)
Ellagic acid and trihydroxybenzoic moieties 6.37 106.2 146.0; 120.0 6.65 114.2 6.78 114.2 6.86 114.2 7.37 110.9 7.44 109.7 A: P. argentea; R: P. recta.
have given contribution to medicine (Woods et al., 2017). Most of these metabolites are not directly involved in growth, development, and re-production of plants, hence were termed as“secondary metabolites” (Ahmad et al., 2017). The expression of secondary metabolites differs in different parts of the plant and is also influenced by the different stages of growth and environmental conditions (Delgoda and Murray, 2017). Once the phytochemical screening was performed, the differences in the extraction that can be obtained using solvents with different pola-rities namely ethyl acetate, methanol, and water, was evaluated. For these purposes colorimetric methods were used to assess the total phenolic, flavonoid, saponins, and triterpenoids contents of the dif-ferent P. recta and P. argentea aerial parts extracts. Upon comparison of the different extracts of both plants, it was found that the water extract of P. argentea (113.29 mg GAE/g extract) was found to contain the highest phenolic content while the ethyl acetate extract of the same
plant (17.44 mg GAE/g extract) contained the lowest phenolic content. Based on this result, it can be suggested that most of the phenolics and reducing compounds present in the plant were hydrophylic. On the other hands theflavonoid content of the studied Potentilla extracts re-sulted better extracted in methanol than in ethyl acetate or water. Also, the methanol extract of P. argentea (519.70 mg QE/g extract) possessed highest saponin content compared with the other solvents indicating that for the extraction of such constituents, alcohol-based solvents ap-pear to be preferable. The water extracts of both Potentilla species showed low triterpenoids content (Table 1). These information can be useful also in the view of the traditional preparations, for example decoction or infusion will contain mainly the antioxidant hydrosoluble compounds but probably will extract only limited amounts of flavo-noids and saponins (Table 4).
Fig. 1.1H-NMR spectra of the methanol extracts from the Potentilla species.
Fig. 3. MS spectrums for the Potentilla species (A: P. argentea; B: P. recta).
Table 3
HPLC-DAD-MS qualitative and quantitative analysis of phenolics in methanolic extracts of Potentilla species.
tr Ion Compound Fragments P. argentea P. recta
1.2 377 caffeic acid derivatives 341 179 3.28 1.70
19.3 301 ellagic acid 284 257 229 201 185 12.55 8.56
19.7 477 (-176) quercetin glucorinide 301 273 179 151 24.62 9.07
20,2 593 luteolin/keamferol rhamnoside hexoside 447 285 255 229 221 0.35 2.02
20.8 447 (-132) isorhamnetin pentoside 315 301 285 3.86 1.75 21.1 591(-176) isorhamnetin glucuronide 315 301 1.54 – 23.0 593 (-308) luteolin/keamferol rutinoside 285 255 229 221 21.46 23.85 23.0 593 (-308) luteolin/keamferol rutinoside 447 285 255 229 221 1.92 2.81 23.8 582.5 unknown (-152) 462 (-152) 342 29.6 711 unknown 675 397 15.7 305 unknown 273 245 229 217 189 173 16.6 1103 ellagitannin derivatives 935 757 633 1.41 7.93 16.7 935.4 ellagitannin derivatives 633 481 39.67 54.56 3.0 783.5 ellagitannin derivatives 481 301 13.06 20.26 6.4 783.4 ellagitannin derivatives 481 301 0.46 28.68 1187 luteolin hexoside 901 799 757 635 593 447 285 267 255 151 1.06 0.59 20 1117 ellagitannin derivatives 935 633 463 301 5.19 6.18
19.4 1200 ellagic acid derivative 1083 935 783 301 1.50 3.05
total mg/g 132 171
Fig. 4. A typical triterpenoid chromatogram from Potentilla species.
Table 4
HPLC-DAD-MS qualitative and quantitative analysis of triterpenoids in metha-nolic extracts of Potentilla species.
tr Ion Compound Fragments P. argentea P. recta
14.7 487.5 Tormentic acid 467 425 405 387 475 203 0.95 0.81 14.9 487.5 Euscaphyc acid 467 425 405 387 475 203 0.13 0.21 17.2 471.5 Pomolic acid 453 411 0.39 0.22 23.2 471.5 Corosolic acid 423 407 393 0.17 0.15 37.3 455.5 Oleanolic acid 407 389 377 0.50 0.37 38.7 455.5 Ursolic acid 407 389 377 0.93 0.66 Total mg/g 3.07 2.42
3.2. Antioxidant properties
The DPPH, ABTS, FRAP, CUPRAC, phosphomolybdenum, and metal chelating assays have been used to evaluate the antioxidant properties of the different extracts of the aerial parts of P. recta and P. argentea. The methanol extract of both Potentilla species showed highest DPPH scavenging properties (Table 5). Previously,Bazylko et al. (2013) re-ported the DPPH scavenging potential of P. recta water and ethanol extracts and fractions. In a paper published byPaduch et al. (2015), the aqueous extract of P. argentea exhibited concentration-dependent DPPH scavenging effect. The ability of the tested extracts of P. recta and P. argentea to quench ABTS has been investigated. Data presented in Table 5, showed that the water extracts of both Potentilla species were the most potent ABTS scavenger compared with the ethyl acetate and methanol extracts. Likewise, the water extracts of the aerial parts of P. reptans and P. speciosa showed good ABTS scavenging potential (Uysal et al., 2017). The DPPH and ABTS scavenging activities of the methanol extract of the aerial part of the studied Potentilla species were lower when compared with the methanol extract of the roots of the same species (Uysal et al., 2019a).
The reducing potential of the different extracts of the aerial part of P. recta and P. argentea was assessed using the CUPRAC and FRAP methods. The CUPRAC activity was higher for water and methanol extracts of both species while ethyl acetate resulted much lower (Table 5). FRAP assays showed more differences between the three extracts being water > methanol > ethyl acetate for both species. As comparison the FRAP and CUPRAC activity of the aerial part of P. anatolica ranged between 223.6–233.0 mg TE/g and 291.7–340.8 mg TE/g for the FRAP and CUPRAC assay respectively (Uysal and Aktumsek, 2015). The CUPRAC and FRAP activity of the tested extracts of P. reptans and P. speciosa have been recently reported. The CUPRAC values ranged from 131.03 to 263.39 mg TE/g extract for P. reptans and 91.85–269.18 mg TE/g extract for P. speciosa, while FRAP ranged from 81.59 to 219.97 mg TE/g extract for P. reptans and 56.97–214.49 mg TE/g extract for P. speciosa (Uysal et al., 2017). The water and methanol extracts of P. reptans and P. speciosa showed higher CUPRAC activity compared with the Potentilla species studied in the present investiga-tion. The ethyl acetate extract exhibited lowest activity for CUPRAC and metal chelating assays.Uysal and Aktumsek (2015)reported that
the ethyl acetate extract of P. anatolica also showed low activity for the metal chelating and CUPRAC assays. In the phosphomolybdenum method (Table 5), the ethyl acetate extracts were least active compared with the methanol and water extracts.Grochowski et al. (2017) re-ported that the water and methanol extracts of P. thuringiaca aerial parts possessed high total antioxidant capacity in the phosphomo-lybdenum assay. Similarly, the water and methanol extracts of P. re-ptans and P. speciosa aerial parts showed higher antioxidant potential in the phosphomolybdenum assay (Uysal et al., 2017). From recently published papers (Uysal et al., 2019a), it was reported that the me-thanol extract of P. recta roots (4.06 mmol TE/g extract) displayed higher total antioxidant capacity using the phosphomolybdenum method compared with the extracts of the aerial parts of P. recta (2.07–3.51 mmol TE/g extract). On the other hand, the antioxidant activity of the methanol extract of P. argentea root (2.55 mmol TE/g extract) was comparable to the methanol and water extracts of P. ar-gentea aerial parts (Table 5). For the metal chelating assay, the me-thanol extract of P. recta and P. argentea roots (Uysal et al., 2019a) showed lower ion chelating abilities as compared with the aerials parts (Table 5).
3.3. Key enzyme inhibition effects
Enzyme inhibition is regarded as the most effective strategy to manage chronic conditions of common health problems (Mahomoodally et al., 2018). For instance, the first line therapy for Alzheimer’s disease includes cholinesterase inhibitors, such as done-pezil, galantamine, and rivastigmine (Guler et al., 2018). Current treatment modalities to manage type 2 diabetes consist ofα-amylase andα-glucosidase inhibitors, such as acarbose, miglitol, and voglibose (Picot et al., 2017;Uysal et al., 2019b). Epidermal tyrosinase inhibition by kojic acid is currently used to manage skin hyperpigmentation conditions. However, the adverse effects associated to the use of cur-rently available enzyme inhibitors have fueled up the interest infinding novel therapeutic agents. The possible application of Potentilla species in the prevention of chronic complications such as, diabetes and neu-rological complications has been evoked previously (Ozkan et al., 2016). FromTable 6, the ethyl acetate, methanol, and water extracts of P. recta and P. argentea inhibited acetylcholinesterase (AChE), Table 5
Antioxidant activities of the tested extracts*.
Plants-Solvents DPPH (mg TE/g extract)
ABTS (mmol TE/g extract) CUPRAC (mg TE/g extract) FRAP (mg TE/g extract) Phosphomolybdenum (mmol TE/g)
Metal chelating ability (mg EDTAE/g) P. argentea-EA 21.90 ± 2.91f 1.27 ± 0.08d 86.66 ± 0.63d 45.63 ± 0.57f 1.20 ± 0.10d 11.52 ± 1.43e P. argentea-MeOH 319.60 ± 5.21b 3.75 ± 0.05b 223.75 ± 2.72b 166.09 ± 1.79d 2.56 ± 0.10b 29.73 ± 0.54c P. argentea-Water 209.09 ± 5.39d 4.37 ± 0.14a 239.99 ± 1.53a 210.81 ± 3.37b 2.55 ± 0.14b 35.97 ± 0.32a P. recta-EA 41.80 ± 4.32e 0.48 ± 0.10e 95.81 ± 1.02c 58.17 ± 0.40e 2.07 ± 0.06c 23.01 ± 0.32d P. recta-MeOH 327.65 ± 1.57a 2.84 ± 0.10a 226.91 ± 3.82b 175.60 ± 1.55c 3.51 ± 0.08a 33.61 ± 0.77b P. recta-Water 263.35 ± 4.87c 3.85 ± 0.05b 228.13 ± 4.21b 216.77 ± 2.87a 2.50 ± 0.05b 35.19 ± 1.56ab
Values expressed are means ± S.D. of three parallel measurements. TE: Trolox equivalent; EDTAE: EDTA equivalent; EA: Ethylacetate. MeOH: Methanol. Different letters indicate significant differences in the extracts (p < 0.05).
Table 6
Enzyme inhibitory properties of the tested extracts.
Plants-Solvents AChE (mg GALAE/g extract)
BChE (mg GALAE/g extract)
Tyrosinase (mg KAE/g extract)
α-amylase (mmol ACAE/g extract)
α-glucosidase (mmol ACAE/g extract) P. argentea-EA 3.38 ± 0.26b 1.18 ± 0.42b 111.06 ± 2.97a 1.31 ± 0.23a 5.01 ± 1.03f P. argentea-MeOH 3.25 ± 0.21b na 109.26 ± 3.09a 1.30 ± 0.09a 45.68 ± 0.51c P. argentea-Water 0.40 ± 0.32c na 25.70 ± 4.10c 0.35 ± 0.03b 35.64 ± 2.55d P. recta-EA 4.26 ± 0.10a 6.18 ± 2.13a 117.78 ± 6.42a 1.61 ± 0.10a 8.49 ± 0.72e P. recta-MeOH 3.39 ± 0.10b na 116.32 ± 6.84a 1.45 ± 0.20a 53.16 ± 1.52a P. recta-Water 0.76 ± 0.59c na 38.92 ± 1.56b 0.58 ± 0.26b 50.61 ± 0.53b
Values expressed are means ± S.D. of three parallel measurements. GALAE: Galatamine equivalent; KAE: Kojic acid equivalent; ACAE: Acarbose equivalent; na: not active. EA: Ethylacetate. MeOH: Methanol. Different letters indicate significant differences in the extracts (p < 0.05).
tyrosinase,α-amylase, and α-glucosidase. The data collected demon-strated that butyrylcholinesterase (BChE) was inhibited by the ethyl acetate extracts only. Additionally, the ethyl acetate extracts of P. recta and P. argentea showed the highest inhibitory activity against AChE as compared with the methanol and water extracts (Table 6). The me-thanol extracts of P. recta and P. argentea roots have been reported to exhibit inhibitory activity against both AChE (P. recta: 4.10 mg GALAE/ g extract, P. argentea: 3.83 mg GALAE/g extract) and BChE (P. recta: 0.37 mg GALAE/g extract, P. argentea: 1.04 mg GALAE/g extract) (Uysal et al., 2017). It is worth mentioning that the ethyl acetate extract of P. recta aerial part exhibited the highest BChE inhibitory action (6.18 mg GALAE/g extract) and this value was equivalent to the in-hibitory action of the ethyl acetate extract of P. reptans aerial part on BChE (6.15 mg GALAE/g extract) (Uysal et al., 2017). Uysal and Aktumsek (2015)reported that the ethyl acetate and methanol extracts of P. anatolica aerial part were potent inhibitors of AChE. This ob-servation is in concordance with the present study. The tyrosinase in-hibitory activity of the methanol extract of P. recta and P. argentea roots (Uysal et al., 2019a) was higher when compared with the different extracts of P. recta and P. argentea aerial parts (Table 6). Several Po-tentilla species, namely P. anatolica, P. bifurca, P. reptans, P. speciosa, P. fruticosa, and P. norvegica were found to act as tyrosinase inhibitors (Piao et al., 2009;Uysal and Aktumsek, 2015;Uysal et al., 2017). Be-sides, Potentilla species have been found to be potent inhibitors of α-glucosidase, a carbohydrate hydrolysing enzyme targeted in the man-agement of type 2 diabetes. P. fulgens, traditionally used to manage type 2 diabetes, revealed to be a potent α-glucosidase inhibitor (Kumar et al., 2013). The methanol extract and fractions of the aerial part of P. astracanica showed strong inhibitory effect on α-glucosidase (Şöhretoğlu et al., 2017). Likewise, in the present study, it was found that the methanol extracts of P. recta (53.16 mmol ACAE/g extract) and P. argentea (45.68 mmol ACAE/g extract) aerial parts were the most active inhibitors ofα-glucosidase compared with the ethyl acetate (P.
recta: 8.49 mmol ACAE/g extract, P. argentea: 5.01 mmol ACAE/g ex-tract) and water (P. recta: 50.61 mmol ACAE/g extract, P. argentea: 35.64 mmol ACAE/g extract) extracts. The inhibitory activity of P. recta and P. argentea roots extracts was more pronounced against α-glucosi-dase compared withα-amylase (Uysal et al., 2019a). Similarly, the tested extracts showed higher glucosidase inhibition and lower α-amylase inhibition (Table 6). Indeed, hypoglycemic agents with mild α-amylase and stronger α-glucosidase inhibition are preferred, since pronouncedα-amylase inhibition prevents proper digestion of carbo-hydrates, which eventually cause gastrointestinal discomforts (Uysal et al., 2018).
3.4. Statistical evaluation
To provide comprehensive insights on the tested extracts further statistical analysis was carried out. The results are shown inFig. 5. In the current study, a good correlation between TPC and CUPRAC (R = 0.99), FRAP (R = 0.98), ABTS (R = 0.94), MCA (R = 0.94) was obtained. Also, significant positive correlations were found between TTC and AChE (R = 0.96), TYR (R = 0.96), and amylase (R = 0.9). In this sense, triterpenoids could be considered as major inhibitors in the tested enzymes (Fig. 5A). Principal component analysis’s (PCA) were performed to detect the relation between the variables of biological properties as well as the tested extracts. In PCA, thefirst two principal components accounted for 88.4% of the total variance (PC1: 64.5% and PC2: 23.9%). Partial least squares discriminant Analysis (PLS-DA) was done to distinguish similar or close extracts (Fig C). According to PLS-DA results, the extracts were classified based on the Potentilla species used in this work. Moreover, three clusters from the tested extracts were observed by Heatmap analysis. The VIP values of TSC, PPBD, CUPRAC, and amylase assays (the value of VIP > 1.0) were the highest and these bioactivities have played an important to classify the tested extracts.
Fig. 5. Statistical evaluations A: Correlation coefficients between total bioactive compounds and biological activities (Pearson Correlation Coefficient (R), p < 0.05); B: Distribution of biological activities on the correlation circle based on PCA; C: Distribution of the extraction methods on the correlation circle based on sPLS-DA, individual sample replications (n = 4) are given in the class prediction model score plot and influence of biological parameters for the total map (VIP: variable importance in the prediction)D: Heatmap of extracts in according to bioactive compounds and biological activities; PPBD: Phosphomolybdenum; MCA: Metal chelating assay; TYR: Tyrosinase inhibition assay; TPC: Total phenolics content; TFC: Totalflavonoids content; TTC: Total triterpenoid content; TSC: Total saponin content).
4. Conclusion
Chemical characterization of the extracts obtained from P. recta and P. argentea aerial parts revealed the presence of polyphenols, triterpene, and sugars. Analysis of the two species by NMR and LC–MS analysis revealed that the pattern of secondary metabolites was almost similar. However, from a quantitative point of view, the highest phenolic, fla-vonoid, and saponin contents were observed from the water and me-thanol extracts of P. argentea. Solvent with different polarities have been used to prepare extracts and the biological activity of the ethyl acetate, methanol, and water extracts of the aerial parts of P. recta and P. argentea were investigated using multiple in vitro standard bio-assays. The higher level of polyphenols can be related also to the higher ob-served antioxidant activity for the methanol and water extracts. Bio-assays namely, DPPH, ABTS, CUPRAC, FRAP, and phosphomo-lybdenum and metal chelating ability, revealed that the in vitro activ-ities are comparable for the two species, whereby the most hydrophilic extracts exhibited very similar biological activities. Comparative ana-lysis of the biological properties of the studied Potentilla species and their different extracts provided an additional insight into the possible therapeutic application of the extracts. The ethyl acetate extracts of the studied Potentilla species were inhibitors of cholinesterases and tyr-osinase in significant way. Methanol extracts showed significant α-glucosidase inhibition. The enzyme inhibitory activities coupled with the observed antioxidant properties can be considered as thefirst at-tempt to establish preliminary data towards the valorisation of these species in the management of several chronic diseases. Data collected from this study tend to support the multiple biological potential ap-plication of the secondary metabolites that are present in the P. recta and P. argentea. This further justify the need for further pharmacolo-gical investigations geared towards potential application in the phar-maceutical, cosmetic, and nutraceutical industries.
Appendix A. Supplementary data
Supplementary material related to this article can be found, in the online version, at doi:https://doi.org/10.1016/j.indcrop.2019.01.047. References
Ahmad, E., Arshad, M., Zakriyya Khan, M., Amjad, M.S., Sadaf, H., Rawalpindi, P., Riaz, I., Sabir, S., Nabila, A.S., 2017. Secondary metabolites and their multidimensional prospective in plant life. J. Pharmacogn. Phytochem. 205, 205–214.
Bazylko, A., Piwowarski, J.P., Filipek, A., Bonarewicz, J., Tomczyk, M., 2013. In vitro antioxidant and anti-inflammatory activities of extracts from Potentilla recta and its main ellagitannin, agrimoniin. J. Ethnopharmacol. 149, 222–227.
Delgoda, R., Murray, J.E., 2017. Chapter 7 - evolutionary perspectives on the role of plant secondary metabolites. In: Badal, S., Delgoda, R. (Eds.), Pharmacognosy. Academic Press, Boston, pp. pp. 93–100.
DiTomaso, J., Kyser, G., 2013. Weed Control in Natural Areas in the Western United States. University of California Weed Research and Information Center, CA, USA.
Fidyt, K., Fiedorowicz, A., Strządała, L., Szumny, A., 2016. β‐caryophyllene and β‐caryophyllene oxide—natural compounds of anticancer and analgesic properties. Cancer Med. 5, 3007–3017.
Grochowski, D.M., Uysal, S., Aktumsek, A., Granica, S., Zengin, G., Ceylan, R., Locatelli, M., Tomczyk, M., 2017. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 20, 365–372.
Guler, G.O., Zengin, G., Karadag, F., Mollica, A., Picot, C.M.N., Mahomoodally, M.F., 2018. HPLC-DAD profiles and pharmacological insights of Onobrychis argyrea subsp isaurica extracts. Comput. Biol. Chem. 76, 256–263.
Halabalaki, M., Vougogiannopoulou, K., Mikros, E., Skaltsounis, A.L., 2014. Recent ad-vances and new strategies in the NMR-based identification of natural products. Curr. Opin. Biotechnol. 25, 1–7.
Kumar, D., Ghosh, R., Pal, B.C., 2013.α-Glucosidase inhibitory terpenoids from Potentilla fulgens and their quantitative estimation by validated HPLC method. J. Funct. Food. 5, 1135–1141.
Li, X.C., Elsohly, H.N., Hufford, C.D., Clark, A.M., 1999. NMR assignments of ellagic acid derivatives. Magn. Reson. Chem. 37, 856–859.
Mahomoodally, M.F., Atalay, A., Nancy Picot, M.C., Bender, O., Celebi, E., Mollica, A., Zengin, G., 2018. Chemical, biological and molecular modelling analyses to probe into the pharmacological potential of Antidesma madagascariense Lam.: a multi-functional agent for developing novel therapeutic formulations. J. Pharm. Biomed. Anal. 161, 425–435.
Mahrous, E.A., Farag, M.A., 2015. Two dimensional NMR spectroscopic approaches for
exploring plant metabolome: a review. J. Adv. Res. 6, 3–15.
Mollica, A., Zengin, G., Locatelli, M., Stefanucci, A., Mocan, A., Macedonio, G., Carradori, S., Onaolapo, O., Onaolapo, A., Adegoke, J., Olaniyan, M., Aktumsek, A., Novellino, E., 2017. Anti-diabetic and anti-hyperlipidemic properties of Capparis spinosa L.: in vivo and in vitro evaluation of its nutraceutical potential. J. Funct. Food. 35, 32–42.
Ozkan, G., Kamiloglu, S., Ozdal, T., Boyacioglu, D., Capanoglu, E., 2016. Potential use of Turkish medicinal plants in the treatment of various diseases. Molecules 21, 257.
Paduch, R., Wiater, A., Locatelli, M., Pleszczynska, M., Tomczyk, M., 2015. Aqueous extracts of selected Potentilla species modulate biological activity of human normal colon cells. Curr. Drug Targets 16, 1495–1502.
Piao, X., Tian, Y., Mi, X., Cui, J., 2009. Tyrosinase inhibition of Potentilla bifurca. China J. Chin. Matera Med. 34, 1952–1954.
Picot, M.C.N., Zengin, G., Mollica, A., Stefanucci, A., Carradori, S., Mahomoodally, M.F., 2017. In vitro and in silico studies of mangiferin from Aphloia theiformis on key en-zymes linked to diabetes Type 2 and associated complications. Med. Chem. (Los Angeles) 13, 633–640.
Rodrigues, E.D., da Silva, D.B., de Oliveira, D.C.R., da Silva, G.V.J., 2009. DOSY NMR applied to analysis offlavonoid glycosides from Bidens sulphurea. Magn. Reson. Chem. 47, 1095–1100.
Rodrigues, F., Moreira, T., Pinto, D., Pimentel, F.B., Costa, A., Nunes, M.A., Albuquerque, T., Costa, H.S., Palmeira-de-Oliveira, A., Oliveira, A.I., 2018. The phytochemical and bioactivity profiles of wild Calluna vulgaris L. flowers. Food Res. Int. 111, 724–731.
Seebacher, W., Simic, N., Weis, R., Saf, R., Kunert, O., 2003. Complete assignments of 1H and 13C NMR resonances of oleanolic acid, 18α‐oleanolic acid, ursolic acid and their 11‐oxo derivatives. Magn. Reson. Chem. 41, 636–638.
Silva, M., David, J.P., Silva, L.C., Santos, R.A., David, J.M., Lima, L.S., Reis, P.S., Fontana, R., 2012. Bioactive oleanane, lupane and ursane triterpene acid derivatives. Molecules 17, 12197–12205.
Şöhretoğlu, D., Sari, S., Özel, A., Barut, B., 2017. α-Glucosidase inhibitory effect of Potentilla astracanica and some isoflavones: inhibition kinetics and mechanistic in-sights through in vitro and in silico studies. Int. J. Biol. Macromol. 105, 1062–1070.
Şöhretoğlu, D., Sari, S., Šoral, M., Barut, B., Özel, A., Liptaj, T., 2018. Potential of Potentilla inclinata and its polyphenolic compounds inα-glucosidase inhibition: ki-netics and interaction mechanism merged with docking simulations. Int. J. Biol. Macromol. 108, 81–87.
Stroh, P., 2014. Potentilla Argentea L. Hoary Cinquefoil. Species Account. Botanical Society of Britain and Ireland Article no: 2683.
Sut, S., Poloniato, G., Malagoli, M., Dall’Acqua, S., 2018a. Fragmentation of the main triterpene acids of apple by LC‐APCI‐MSn. J. Mass Spectr. 53, 882–892.
Sut, S., Dall’Acqua, S., Baldan, V., Kamte, S.L.N., Ranjbarian, F., Nya, P.C.B., Vittori, S., Benelli, G., Maggi, F., Cappellacci, L., 2018b. Identification of tagitinin C from Tithonia diversifolia as antitrypanosomal compound using bioactivity-guided fractio-nation. Fitoterapia 124, 145–151.
Sut, S., Zengin, G., Dall’Acqua, S., Gazdová, M., Šmejkal, K., Bulut, G., Dogan, A., Haznedaroglu, M.Z., Aumeeruddy, M.Z., Maggi, F., 2019. Paeonia arietina and Paeonia kesrounansis bioactive constituents: NMR, LC-DAD-MSfingerprinting and in vitro assays. J. Pharm. Biomed. Anal. 165, 1–11.
Tomczyk, M., 2006. Secondary metabolites from Potentilla argentea. Biochem. Syst. Ecol. 34, 770–773.
Tomczyk, M., Latté, K.P., 2009. Potentilla—a review of its phytochemical and pharma-cological profile. J. Ethnopharmacol. 122, 184–204.
Tomczyk, M., Leszczyńska, K., Jakoniuk, P., 2008. Antimicrobial activity of Potentilla species. Fitoterapia 79, 592–594.
Uysal, S., Aktumsek, A., 2015. A phytochemical study on Potentilla anatolica: an endemic Turkish plant. Ind. Crop. Prod. 76, 1001–1007.
Uysal, S., Zengin, G., Locatelli, M., Bahadori, M.B., Mocan, A., Bellagamba, G., De Luca, E., Mollica, A., Aktumsek, A., 2017. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. Speciosa L. And P. Reptans Willd.) and their chemical composi-tion. Front. Pharmacol. 8, 290.
Uysal, S., Aktumsek, A., Picot-Allain, C.M., Unuvar, H., Mollica, A., Georgiev, M.I., Zengin, G., Mahomoodally, M.F., 2018. Biological, chemical and in silicofingerprints of Dianthus calocephalus Boiss.: a novel source for rutin. Food Chem. Toxicol. 113, 179–186.
Uysal, S., Zengin, G., Mahomoodally, M.F., Yilmaz, M.A., Aktumsek, A., 2019a. Chemical profile, antioxidant properties and enzyme inhibitory effects of the root extracts of selected Potentilla species. S. Afr. J. Bot. 120, 124–128.
Uysal, A., Ozer, O.Y., Zengin, G., Stefanucci, A., Mollica, A., Picot-Allain, C.M.N., Mahomoodally, M.F., 2019b. Multifunctional approaches to provide potential phar-macophores for the pharmacy shelf: heracleum sphondylium L. Subsp. Ternatum (Velen.) Brummitt. Comp. Biol. Chem. 78, 64–73.
Vrhovsek, U., Guella, G., Gasperotti, M., Pojer, E., Zancato, M., Mattivi, F., 2012. Clarifying the identity of the main ellagitannin in the fruit of the strawberry, Fragaria vesca and Fragaria ananassa Duch. J. Agric. Food Chem. 60, 2507–2516.
Waldbauer, K., Seiringer, Gn., Nguyen, D.L., Winkler, J., Blaschke, M., McKinnon, R., Urban, E., Ladurner, A., Dirsch, V.M., Zehl, M., 2015. Triterpenoic acids from apple pomace enhance the activity of the endothelial nitric oxide synthase (eNOS). J. Agric. Food Chem. 64, 185–194.
Woods, N., Niwasabutra, K., Acevedo, R., Igoli, J., Altwaijry, N.A., Tusiimire, J., Gray, A.I., Watson, D.G., Ferro, V.A., 2017. Chapter 11 - natural vaccine adjuvants and immunopotentiators derived from plants, fungi, marine organisms, and insects. In: Schijns, V.E.J.C., O’Hagan, D.T. (Eds.), Immunopotentiators in Modern Vaccines (Second Edition). Academic Press, pp. pp. 211–229.
Xue, P.F., Zhao, Y.Y., Wang, B., Liang, H., 2006. Secondary metabolites from Potentilla discolor Bunge (Rosaceae). Biochem. Syst. Ecol. 11, 825–828.
Zengin, G., Atasagun, B., Aumeeruddy, M.Z., Saleem, H., Mollica, A., Bahadori, M.B., Mahomoodally, M.F., 2019. Phenolic profiling and in vitro biological properties of two Lamiaceae species (Salvia modesta and Thymus argaeus): a comprehensive eva-luation. Ind. Crop. Prod. 128, 308–314.