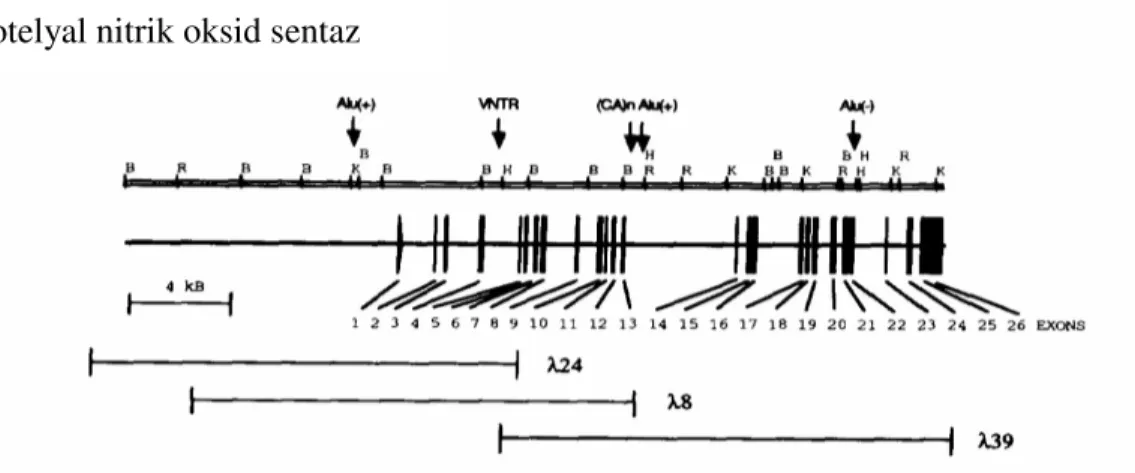
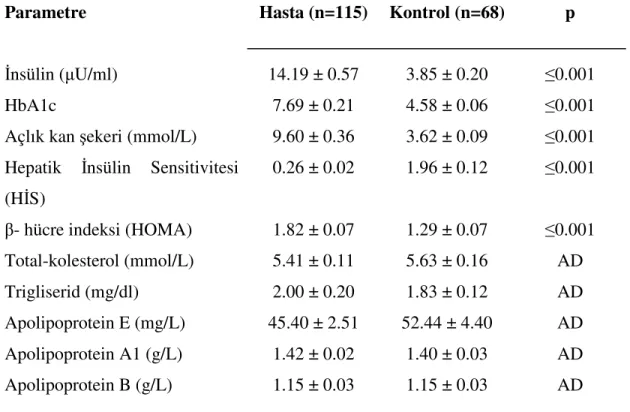
İnsüline bağımlı olmayan diabetes mellitus’lu hastalarda endotelyal nitrik oksid sentaz (eNOS) GLU298ASP polimorfizminin araştırılması
Tam metin
Şekil








Benzer Belgeler
These are: “The more battery, the more brightness of the bulbs” (3%), “Current is consumed by its component” (3%), “The concept of current, energy and potential differences
Üç-boyutlu ERT çalışmaları paralel veya birbirine dik profiller üzerinde ölçümler alınarak gerçekleştirilebilmektedir. Jeofizik ölçümlerde profil doğrultusunun aranan
Fiziki sermayeyi temsilen kullanılan brüt sabit sermaye oluşumunun gayri safi yurtiçi hâsıladaki payı ile ekonomik büyümenin vekil değişkeni; 2010 yılı sabit fiyatlarla
Borsa İstanbul’da işlem gören 168 şirketin 20.664 aylık getiri değişkeninden oluşan toplam 127.008 gözlem değeri ile şirketlerin finansal başarısızlıkları
Bu açıdan Doğu Anadolu ağızlarına baktığımızda karşımıza Eski Türkçedeki ŋ sesinin g olarak korunması: ög ‘ön’, söz içi ve sonundaki g sesinin
Kombinasyon grubunda, 12 aydan daha uzun süre ACEi/ARB tedavisi uygulanmış olan hastalarımızın, tedavi öncesi protein ve albumin atılımlarının, bu tedavileri 12 aydan daha
arasındaki ilişkinin anlamlılık düzeyleri s.141 Tablo 40: Anlamlılık düzeylerine göre kadın çalışanların çalıştıkları birime bağlı örgütsel bağlılık
İkinci bölümde ise kapalı uçlu olarak “boş zaman değerlendirilmesinde tercih edilen kaynaklar, doğal çevrede yapılan rekreasyon eylemleri, Afyonkarahisar’ı
