Effects of TGF-β1 Overexpression on Biological Characteristics of Human Dental Pulp-derived Mesenchymal Stromal Cells
Tam metin
Şekil

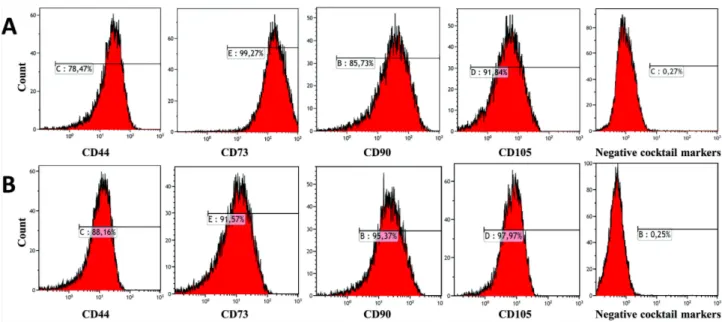
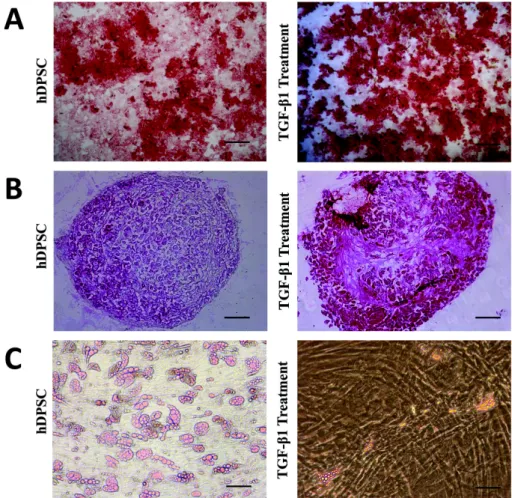
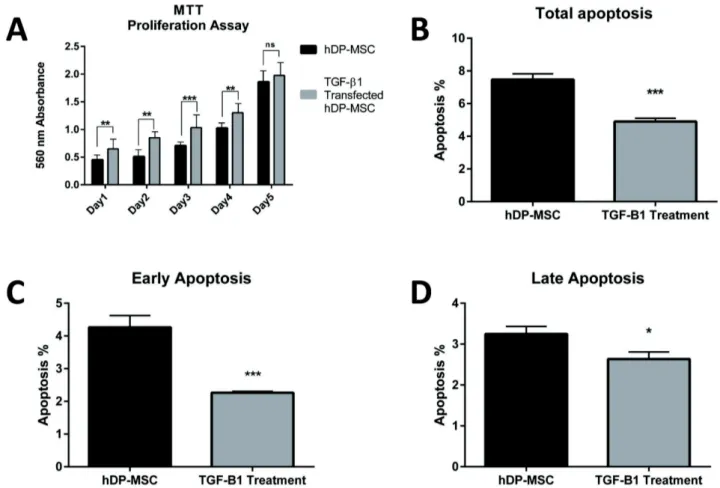
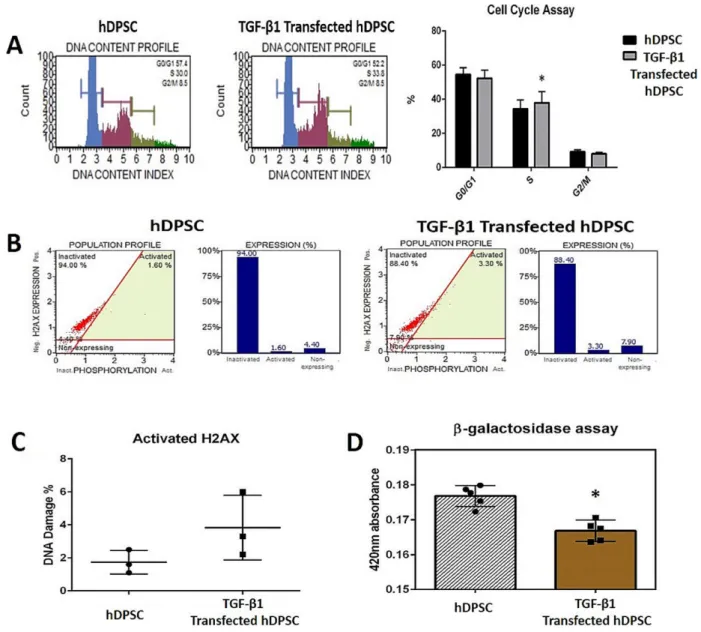
Benzer Belgeler
alternative procedures to get an initial feasible solution: l j we randomly generate 100 feasible solutions and run the heuristic starting from each, recording the best
The average tensile strength values and Duncan test results of the standard- and micro-size Scots pine wood specimens are shown in Table 2. The tensile strength values of
In our study we investigated quite a large group of tissues and it was also clearly seen in our study that there was a significant increase in antioxidant capacity (particular- ly
Araştırma evreni Düzce İl Merkezi’nde sokakta çalışan çocuklardan ve onların ailelerinden oluşmaktadır. Düzce İl Em- niyet Müdürlüğü Çocuk Şubesi’nden alı-
Anabilim Dalımızca verilen görüş ile yeni cetvele göre yapılan değerlendirmeye göre farklı sonuç çıkan 5 olgu incelendiğinde “organlardan birinin işlevinin sürekli
Türk edebiyatında modernist görüşlerle bencilleşen, tabiatı kendi arzusuna göre talan eden insanoğlunun eleştirisi, ayrıntılı olarak ele alınabilecek bir konu olmakla
Her 3 gruba günde 1gr kalsiyum ilave edilmifl risedronat gruplar›nda 2 y›l sonundaki spinal ve femur kemik yo¤unlu¤unda artma istatistiksel olarak anlaml› bulunmam›fl, 3
b: Different letters in the same column are statistically significant (p<0.05. Corrected Bonferroni Test)... 88 Figure 3. a) Achilles tendon without collagenase treatment,
