Seasonal variations in serum concentrations of melatonin,
testosterone, and progesterone in Arabian horse
*Çiğdem ALTINSAAT1, Aykut Göktürk ÜNER2, Nesrin SULU1, Ahmet ERGÜN3
1 Department of Physiology, Faculty of Veterinary Medicine, University of Ankara, Ankara, Turkey; 2 Department of Physiology,
Faculty of Veterinary Medicine, University of Adnan Menderes, Aydın, Turkey; 3 Department of Physiology, Faculty of Medicine,
University of Ankara, Ankara, Turkey.
Summary: The objective of this study was to characterize seasonal variations in serum concentrations of melatonin, testosterone, and progesterone in Arabian horses under natural photoperiodic conditions. Peripheral blood samples were collected during breeding and non-breeding seasons from mares and stallions. Serum concentrations of melatonin, testosterone, and progesterone were determined by radioimmunoassay. Serum concentrations of melatonin were greater in the non-breeding season (42.41±1.59 and 37.68±1.55 pg/ml) when compared to breeding season (23.52±1.24 and 17.22±2.10 pg/ml) in both mares and stallions, respectively. Melatonin concentrations were low but not different between the luteal and follicular phases during breeding season in cyclic mares. Mares had greater concentrations of melatonin than stallions in both breeding and non-breeding season. Mean concentrations of testosterone were greater during breeding season (6.58±0.50 ng/ml) than non-breeding season (3.64±0.48 ng/ml) in stallions. There was a negative correlation (r=-0.658, p<0.01) with melatonin and testosterone levels in both season in stallions. A negative correlation (r=-0.829, p<0.05) in luteal phase and a positive correlation (r=0.847, p<0.05) in non-breeding season were found between melatonin and progesterone concentrations in mares. We conclude that changes in day length act as a regulator and photoperiod can be used to modify the seasonal reproduction in Arabian horse.
Key words: Arabian horse, melatonin, photoperiod, progesterone, testosterone.
Arap atlarında serum melatonin, testosteron ve progesteron seviyelerinin mevsimsel değişimleri Özet: Bu çalışmanın amacı, doğal fotoperiyodik koşullarda Arap atlarının serum melatonin, testosteron ve progesteron seviyelerinin mevsimsel değişimlerini belirlemektir. Kısrak ve aygırlardan üreme döneminde ve üremenin olmadığı dönemde toplanan kan örneklerinden, radioimmunoassay yöntemi ile serum melatonin, testosteron ve progesteron seviyeleri belirlenmiştir. Sırasıyla hem kısrak hem de aygırlarda melatonin seviyeleri üremenin olmadığı dönemde (42.41±1.59 ve 37.68±1.55 pg/ml) üreme dönemine göre (23.52±1.24 ve 17.22±2.10 pg/ml) daha yüksek bulunmuştur. Üreme döneminde östrus siklusu gösteren kısraklarda melatonin seviyelerinin düşük fakat luteal ve foliküler dönemler arasında fark yoktu. Kısraklar hem üreme döneminde hem de üremenin olmadığı dönemde aygırlara göre daha yüksek melatonin seviyelerine sahipti. Aygırlarda üreme dönemindeki (6.58±0.50 ng/ml) testosteron seviyeleri üremenin olmadığı dönemden (3.64±0.48 ng/ml) daha yüksekti. Aygırlarda her iki dönemde melatonin ile testosteron seviyeleri arasında negatif bir ilişki vardı (r=-0.658, p<0.01). Kısrakların melatonin ve progesteron seviyeleri arasında ise luteal dönemde negatif (r=-0.829, p<0.05), üremenin olmadığı dönemde de pozitif bir ilişki (r=0.847, p<0.05) bulundu Bu çalışma ile Arap atlarında, gün uzunluğundaki değişikliklerin mevsimsel üremede bir düzenleyici olarak görev aldığı ve fotoperiyodun bu düzenlemede değiştirici etkisi olduğu bulundu.
Anahtar sözcükler: Arap atı, fotoperiyod, melatonin, progesteron, testosteron.
* This research was supported by a grant of The Scientific and Technological Research Council of Turkey, (TUBITAK, Project
number: 105 O 209).
Introduction
Seasonal changes in day length have effects on reproduction in many species (8). The pineal gland is able to receive photo information and measure day length and regulate the secretion of melatonin accordingly (3,19). Melatonin is a key player in controlling reproduction, sleep-wake cycles and other phenomena showing circadian or circanual rhythm (5,16,24). In northern hemisphere, animals are exposed to more
melatonin during the night, and that during the longer periods of darkness in winter. Conversely, melatonin levels decrease during long days in summer (16,17). The localization of melatonin receptors in the hypothalamus and pituitary indicates a possible interaction of melatonin in regulation of reproduction in horse (35).
The horse is a seasonal polyestrous species with onset of the breeding season occurring in spring, associated with increase in daylight, temperature, and
availability of food (27,30). During the fall, the duration of secretion of melatonin from the pineal gland is increased, reflecting the increased duration of darkness in horse. These longer nights may provide a cue for the mare to enter into anestrus (17). Besides, during the non-breeding season, it has been reported that the stallion exhibits partial testicular atrophy, decreased testosterone levels and libido, and decreased accessory fluid production due to the melatonin concentrations are the highest (38).
The breeding season, which begins from approximately April through October, in the Northern Hemisphere, is the time of the year characterized by sexual patterns in mares and stallions. Anestrus is the period of sexual incompetence, indeed of sexual indifference within the annual reproductive cycle and occurs during November to January in the Northern Hemisphere (29).
The short day length characteristic of fall and winter causes an increase in the daily duration of melatonin secretion, and this result in decreased secretion of gonadotropic hormones and the cessation of ovulatory activity (14). During the non-breeding season, most of the mares maintain progesterone (P4) concentrations below 1 ng/ml which do not show estrus signs and have ovaries that are small and smooth (29). The decrease in the amount of melatonin produced causes an increase in the amount of GnRH, LH and FSH produced and secreted during breeding season. The increase in LH and FSH brings the horse into estrus (9,34).
Exogenous melatonin administration is reported to decrease plasma testosterone concentrations in stallions and change the annual reproductive rhythm in pony mares (32). In contrast, artificial photostimulation produced a sharp rise in testosterone levels in stallion (4).
Limited information is available about the seasonality of reproductive activity of the Arabian horse bred in Turkey. The aim of the present work was to determine melatonin concentrations and its interaction with reproductive activity during breeding and non-breeding seasons in Arabian horses.
Materials and Methods Animals
A group of 38 healthy Arabian mares (n=20) and stallions (n=18), ranged from 4 to 8 yr of age were used in June [15 h of light (L):9 h of darkness (D)] and December (9L:15D) 2006. Mares were classified into two groups as follows: breeding season (estrus cycle period, n=13), non-breeding season (anovulatory period, n=7), and stallions as breeding season (n=12), non-breeding season (n=6). The horses were fed with the
standard adult horse diet and housed in the stable and kept separately according to gender at Anatolia Station/Eskişehir (General Directorate of Agricultural Enterprises - TIGEM) in Turkey. The station is 890 m above sea level with a longitude of 30:32 E and a latitude of 39:46 N. The experiment was conducted under natural photoperiod.
Blood Collection
Peripheral blood samples (10 ml) were collected via jugular venipuncture from each horse into vacuum collection tubes (Vacuette®, Greiner Labortechnik, Kremsmuenster, Austria) in early morning (at 07:00 am) during breeding and non-breeding seasons. Sera were decanted immediately into eppendorfs after centrifugation at 3000 rpm for 10 minutes and stored at – 70 0C until endocrine analysis.
Experimental Procedures
Melatonin, testosterone, and progesterone levels were evaluated in breeding and non- breeding season. Mares were checked for ovarian activity by monitoring serum P4 concentrations in blood samples taken twice a month, in June and December 2006. Mares were checked for estrous behavior with a vigorous stallion. Only mares that had demonstrated typical cyclic patterns of estrus during the breeding season were used. Blood samples taken during the study were subsequently assayed for P4 to confirm that the mares had ovulated and were luteal in each period of sample collection. Mares were considered to be in the luteal phase by P4≥1 ng/ml (12). Mares that were also verified to have P4 levels lower than 1 ng/ml throughout the non-breeding season of blood sample collection were considered to be anestrus (11).
Endocrine Analysis
Melatonin assay: Blood serum melatonin concentrations were measured by using a commercially available kit (Labor Diagnostika Nord GmbH & Co. KG, cat no: BA 3300, Nordhorn, Germany). Serum samples (200 µl) were assayed in duplicate. The sensitivity of the assays averaged 2 pg/ml and intra - assay coefficients of variation were 11.4%.
Testosterone and progesterone assays: Testosterone and progesterone assays were performed using commercial RIA kits (Diagnostic Systems Laboratories, Inc., cat no: DSL-4100 and DSL-3400 Webster, Texas, USA) which has been validated in previous studies in equine blood serum (1,36). The minimum detectable concentrations were 0.10 ng/ml (0.318 nmol/L) in P4 assay and 0.05 ng/ml (0.174 nmol/L) in testosterone assay. The intra - assay coefficients of variation were 5.7% and 16.6% for testosterone and P4 assays, respectively. All samples were run in duplicate and within one assay.
Statistical Analyses
Two - way ANOVA was performed in order to examine the effects of gender or season differences on melatonin levels. We considered that gender (p<0.01) and season (p<0.001) significantly affected on melatonin levels. For that reason, changes in serum concentrations of melatonin were examined in breeding and non-breeding seasons by independent-samples T test in mares and stallions. Seasonal effect was evaluated by same statistical test for testosterone in stallion. Furthermore, in mares, changes of melatonin and P4 levels in luteal phase, follicular phase, and non-breeding season were examined by one-way ANOVA and Kruskall-Wallis ANOVA, respectively. The means were further analyzed by post-hoc comparison tests, such as Tukey and Duncan except P4 levels. Median P4 levels were compared by Mann-Whitney test after a modified Bonferroni correction. The results were presented as the mean±SEM. Correlation analyses were performed to demonstrate a possible relationship between melatonin with other hormones for each gender. Significance of correlation (2-tailed) between melatonin and testosterone was assessed using coefficient of correlation calculated by Pearson correlation coefficient. Correlation between melatonin and progesterone was determined by use of the Spearman rank correlation coefficient. p values <0.05 were considered to be significant. All statistical analyses were performed using a software program (SPSS, serial number for this license: 9024147).
Results
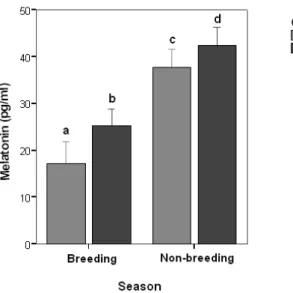
Seasonal changes of mean serum melatonin concentrations in stallions and mares are presented in Table 1. Serum concentrations of melatonin were significantly (p<0.001) increased during the short day period (37.68±1.55 pg/ml) compared to the long day period (17.22±2.10 pg/ml) in stallions. Melatonin concentrations also increased significantly (p<0.001) during the short day period (42.41±1.59 pg/ml) compared to the long day period (23.52±1.24 pg/ml) in mares. In breeding season, melatonin concentrations of cyclic mareswere not significant (p>0.05) between the luteal (23.28±1.96 pg/ml) and follicular (24.66±2.47 pg/ml) phases (Table 2). Besides, mares had significantly greater concentrations of melatonin than stallions in both breeding (p<0.01) and non-breeding season (p<0.05) (Table 1, Fig 1).
Mean serum testosterone levels were 6.58±0.50 ng/ml and 3.64±0.48 ng/ml in breeding and non breeding season, respectively (Table 3). Testosterone concentrations increased significantly (P<0.01) in stallions during breeding season. In addition to this finding, there was a
negative correlation (r=-0.658, p<0.01) with melatonin and testosterone concentrations (Table 3).
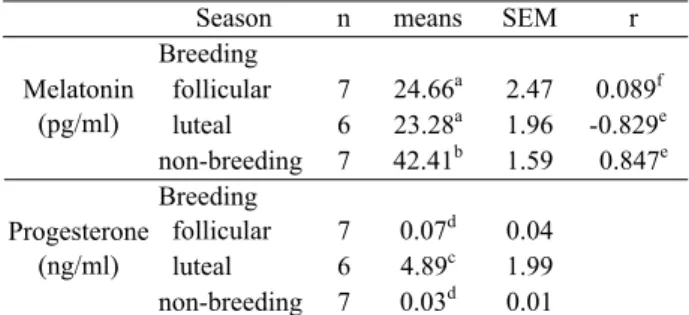
Mean serum P4 concentrations were significantly (p<0.01) highest in luteal phase (4.89±1.99 ng/ml, median: 3.36) when compared follicular phase (0.07±0.04 ng/ml, median: 0) and anovulatory period (0.03±0.01 ng/ml, median: 0) in mares (Table 2). Concentrations of P4 of two mares have greater than 1 ng/ml in non-breeding season suggesting these mares still continue estrous cyclicity. There is a negative correlation between melatonin and P4 concentrations in luteal phase (r=-0.829, p<0.05) and a positive correlation in non-breeding season (r=0.847, p<0.05) whereas, no correlation was found in follicular phase (r=0.089, p>0.05). (Table 2).
Fig. 1. Differences in mean serum melatonin concentrations of Arabian mares and stallions during breeding and non-breeding seasons. Each bar represents the mean±SEM. Different letters above each bar represent statistically differences (a,b: p<0.01 and c,d: p<0.05).
Şekil 1. Arap kısrak ve aygırlarda üreme döneminde ve üremenin olmadığı dönemde ortalama serum melatonin düzeyleri arasındaki farklılık.
Table 1. Serum melatonin concentrations in Arabian mares and stallions in breeding and non-breeding season.
Tablo 1. Üreme döneminde ve üremenin olmadığı dönemde Arap kısrak ve aygırlarda serum melatonin düzeyleri.
Mares Season n means SEM p Breeding 13 23.52 1.24 Melatonin (pg/ml) non-breeding 7 42.41 1.59 <0.001 Stallions Season n means SEM p Breeding 12 17.22 2.10 Melatonin (pg/ml) non-breeding 6 37.68 1.55 <0.001
Table 2. Serum melatonin and progesterone concentrations and correlation analysis during breeding and non breeding season in Arabian mares.
Tablo 2. Üreme döneminde ve üremenin olmadığı dönemde Arap kısraklarda serum melatonin ve progesteron düzeyleri ve korelasyon analizi.
Season n means SEM r
Breeding follicular 7 24.66a 2.47 0.089f luteal 6 23.28a 1.96 -0.829e Melatonin (pg/ml) non-breeding 7 42.41b 1.59 0.847e Breeding follicular 7 0.07d 0.04 luteal 6 4.89c 1.99 Progesterone (ng/ml) non-breeding 7 0.03d 0.01 a,b and c,d: Means with different superscripts within the same
column, in different parameters, are significantly different (a,b: p < 0.001 and c,d: p<0.01)
e,f: Statistical correlation analysis between melatonin and
progesterone levels during breeding and non breeding season in Arabian mares (e: p<0.05, f: p>0.05)
Table 3. Mean serum testosterone concentrations of Arabian stallions during breeding and non breeding season. Correlation analysis between melatonin and testosterone levels in Arabian stallions.
Tablo 3. Arap aygırlarında üreme döneminde ve üremenin olmadığı dönemde ortalama serum testosteron düzeyleri ile melatonin ve testosteron düzeyleri arasındaki ilişki.
Season n Means SEM p r
Breeding 12 6.58 0.50 Testosterone
(ng/ml) non-breeding 6 3.64 0.48 <0.01 -0.658
Φ Φ: Statistical correlation analysis between melatonin and
testosterone levels in Arabian stallions (p<0.01). Discussion and Conclusion
Photoperiodism has been known as a visual signal in seasonal breeders such as sheep, horse, and hamster (11). Regulation of reproductive activity via photoperiodic cues mediated by the secretion of melatonin from the pineal gland is reported in ewes (6,28), but the relationship is less clear in mares (11,25). Under natural photoperiodic conditions, serum concentrations of melatonin are higher in anestrus mares (11,33). In response to artificial photoperiod, alterations in ovulatory season also exhibit in mares. First ovulation occur earlier in mares exposed an artificial, long photoperiod during anovulatory season (15,26). Horse is a seasonal breeder and its reproductive efficiency varies throughout the year. In the northern hemisphere mares enter anestrus and stallions semen quality sharply declines during late fall and early winter (16,30). Information on photoperiod and the physiological reproductive season in Arabian horse is very limited. This study tested the role of melatonin in the regulation
of seasonal physiological change in the Arabian mares and stallions. We found that serum melatonin levels were greater non-breeding season than breeding season in both mares and stallions (p<0.001). Mean concentrations of melatonin in mares were 23.52 pg/ml and 42.41 pg/ml in breeding and non-breeding seasons, respectively. In stallions, melatonin levels were 17.22 pg/ml and 37.68 pg/ml in breeding and non-breeding seasons, respectively. Diekman et al (11) reported that mean melatonin levels in mares were 16 pg/ml in June and 19.5 pg/ml in December. Guillaume et al (19) reported that mean concentration of nocturnal melatonin were 24 pg/ml during autumn in 110 pony mares. However, in that study, variability of nocturnal melatonin levels were very high (ranged from <10 pg/ml to 130 pg/ml). In this study,melatonin concentrations of cyclic mares were low in breeding season. However, there was no difference between the luteal and follicular phases (Table 2). Besides, mares had significantly greater concentrations of melatonin than stallions in both breeding and non-breeding season (Fig 1). Thus, it has been reported that females had greater concentrations of melatonin than males in rat (7,20).
Plasma testosterone concentrations in the stallion vary according to seasonal reproductive state. Thus, plasma testosterone concentrations are higher in the summer and lower in the winter (10,23). Variations in concentrations of testosterone in stallions are reported to be episodic and are not consistently exhibited with pulses in LH concentrations (38,39). Thompson et al. (38) reported that serum testosterone concentrations under an artificial photoperiod of increasing daylight during the winter months showed an accelerated seasonal increase in stallions. In that study, mean concentrations of testosterone in stallions ranged from 0.7 ng/ml in December to 4.1 ng/ml in August. As a long day breeder, horse reduces reproductive capacity during the decreased photoperiod of the winter months, whereas, short day breeders, such as the sheep, are positively affected during the same period (16). In addition, Tamarkin et al (37) reported that melatonin attenuated gonadal function and decreased gonadotropin concentrations in both sexes of hamster, which is a long day breeder. In the present study, mean concentrations of testosterone in Arabian stallions were 3.64 ng/ml and 6.58 ng/ml in non-breeding and breeding season, respectively. Serum testosterone levels were greater in breeding season compared with non-breeding season (p<0.01) and there was a negative correlation between serum melatonin and testosterone levels in Arabian stallions (r=-0.658, p<0.01). Argo et al (4) reported that exogenous melatonin treatment decreased testosterone levels in pony stallions.
Levels of estrogen and P4 reflect ovarian activity. Both are low in anestrus season. Progesterone is baseline throughout anestrus and transition, but rises when a corpus luteum is formed after ovulation. Amer et al. (2) reported that following ovulation, plasma P4 levels showed a continuous increase corresponding 0.58–3.48 ng/ml values in luteal phase in Arabian mares. A similar increase 2.2–5.4 ng/ml at the same period of cycle has been observed in large saddle mares (22). These findings come in agreement with our results where mean concentrations of P4 were 0.07±0.04 and 4.89±1.99 in follicular and luteal phase, respectively. Progesterone concentrations onset of breeding season were significantly affected by the extended photoperiod (21,31). Diekman et al. (11) reported that melatonin levels were not influenced by P4. However, in the present study, there is a significantly correlation between melatonin and P4 concentrations in luteal phase(r=-0.829, p<0.05) and non-breeding season (r=0.847, p<0.05). Besides, two mares had higher P4 level than 1 ng/ml and exhibited estrous cycles in non-breeding season. Moreover, melatonin levels of both mares were similar to that observed in the other mares that were non-cyclic. The studies have shown that treatment of mares with melatonin does not advance the onset of seasonal anestrus in mature mares (14,18). This suggests that other factors exist to maintain estrus cycle in non-breeding season in mares apart from melatonin even if a correlation exists between melatonin and progesterone levels during non-breeding season in this study. Melatonin levels do not seem to be directly affected on reproductive hormones in mares. It has been considered that at the onset anestrus or estrus, aging, pregnancy, lactation, body fatness, and leptin might play a role in mares (13). Nevertheless, the day length seems as a primary signal for the termination of seasonal reproductive activity in mares.
In conclusion, this study tested the effect of day length in the regulation of seasonal reproductive activity in the Arabian mares and stallions. Our observations suggested a role for melatonin in the termination of breeding activity in the Arabian horse. Melatonin concentrations during the non-breeding season differ from the breeding season and melatonin secretion, therefore, provides a physiological unambiguous circannual rhythm with longest periods of secretion in the winter and shortest periods of secretion in the summer. The role of other hormones, such as prolactin, leptin, and ovarian steroids, in this regard, is not known and has not been studied in Arabian horses. However, additional study will be required to determine whether these hormones and other factors such as aging, nutrition, climate are involved in seasonal changes of in reproductive activity of Arabian horses.
References
1. Aljarrah AH (2004): Methods to Induce Earlier Onset of
Cyclicity in Transitional Mares. Master's Thesis, Louisiana
State University.
2. Amer HA, Shawki G, Ismail R (2008): Ovarian and
endocrine changes during oestrus and early pregnancy in Arabian mares. The Internet J Vet. Med., 4, 1.
Access:http://www.ispub.com/ostia/index.php?xmlFilePat h=journals/ijvm/vol4n1/mare.xml Access date: 25.05.2008. 3. Arendt J (2005): Melatonin: characteristics, concerns,
and prospects. J Biol Rhythms, 20, 291-303.
4. Argo CM, Cox JE, Gray JL (1991): Effect of oral
melatonin treatment on the seasonal physiology of pony stallions. J Reprod Fertil Suppl, 44, 115-125.
5. Baker FC, Driver HS (2007): Circadian rhythms, sleep,
and the menstrual cycle. Sleep Med, 8, 613-622.
6. Bittman EL, Dempsey RJ, Karsch FJ (1983): Pineal
melatonin secretion drives the reproductive response to day length in the ewe. Endocrinology, 113, 2276–2283.
7. Brotto LA, Barr AM, Gorzalka BB (2000): Sex
differences in forced-swim and open-field test behaviours after chronic administration of melatonin. Eur J
Pharmacol, 402, 87-93.
8. Clay CM, Clay JN (1992): Endocrine and testicular
changes associated with season, artificial photoperiod, and the peri-pubertal period in stallions. Vet Clin North
Am Equine Pract, 8, 31-56.
9. Cleaver BD, Grubaugh WR, Davis SD, Sheerin PC, Franklin KJ, Sharp DC (1991): Effect of constant light
exposure on circulating gonadotrophin levels and hypothalamic gonadotrophin-releasing hormone (GnRH) content in the ovariectomized pony mare. J Reprod Fertil
Suppl, 44, 259-266.
10. Cox JE, Redhead PH, Jawad NM (1988): The effect of
artificial photoperiod at the end of the breeding season on plasma testosterone concentrations in stallions. Aust Vet
J, 65, 239-241.
11. Diekman MA, Braun W, Peter D, Cook D (2002):
Seasonal serum concentrations of melatonin in cycling and noncycling mares. J Anim Sci, 80, 2949-2952.
12. Ferreira-Dias G, Claudino F, Carvalho H, Agrícola R, Alpoim-Moreira J, Robalo Silva J (2005): Seasonal
reproduction in the mare: possible role of plasma leptin, body weight and immune status. Domest Anim Endocrinol,
29, 203-213.
13. Fitzgerald BP, Davison LA, McManus CJ (2000):
Evidence for a seasonal variation in the ability of exogenous melatonin to suppress prolactin secretion in the mare. Domest Anim Endocrinol, 18, 395-408.
14. Fitzgerald BP, McManus CJ (2000): Photoperiodic
versus metabolic signals as determinants of seasonal anestrus in the mare. Biol Reprod, 63, 335-340.
15. Freedman LJ, Garcia MC, Ginther OJ (1979): Influence
of photoperiod and ovaries on seasonal reproductive activity in mares. Biol Reprod, 20, 567-574.
16. Gerlach T, Aurich JE (2000): Regulation of seasonal
reproductive activity in the stallion, ram and hamster.
Anim Reprod Sci, 58, 197-213.
17. Guerin MV, Deed JR, Kennaway DJ, Matthews CD (1995): Plasma melatonin in the horse: measurements in
natural photoperiod and in acutely extended darkness throughout the year. J Pineal Res, 19, 7-15.
18. Guillaume D, Rio N, Toutain PL (1995): Kinetic studies
and production rate of melatonin in pony mares. Am J
Physiol, 268, 1236-1241.
19. Guillaume D, Zarazaga LA, Malpaux B, Chemineau P (2006): Variability of plasma melatonin level in pony
mares (Equus caballus), comparison with the hybrid: mules and with jennies (Equus asinus). Reprod Nutr Dev,
46, 633-639.
20. Hill MN, Brotto LA, Lee TT, Gorzalka BB (2003):
Corticosterone attenuates the antidepressant-like effects elicited by melatonin in the forced swim test in both male and female rats. Prog Neuropsychopharmacol Biol
Psychiatry, 27, 905-911.
21. Hodge SL, Kreider JL, Potter GD, Harms PG, Fleeger JL (1982): Influence of photoperiod on the pregnant and
postpartum mare. Am J Vet Res, 43, 1752-1755.
22. Holtan DW, Nett TM, Estergreen VL (1975): Plasma
progestins in pregnant, postpartum and cycling mares. J
Anim Sci, 40, 251-260.
23. Johnson L, Thompson DL Jr (1983): Age-related and
seasonal variation in the Sertoli cell population, daily sperm production and serum concentrations of follicle-stimulating hormone, luteinizing hormone and testosterone in stallions. Biol Reprod, 29, 777-789.
24. Keefe DL, Turek FW (1985): Circadian time keeping
processes in mammalian reproduction. Oxf Rev Reprod
Biol, 7, 346-400.
25. Kilmer DM, Sharp DC, Bergland LA, Grubaugh W, McDowell KJ, Peck LS (1982) Melatonin rhythms in
pony mares and foals. J Reprod (Fertil) Suppl, 32, 303–307.
26. Kooistra LH, Ginther OJ (1975): Effect of photoperiod
on reproductive activity and hair in mares. Am J Vet Res,
36, 1413-1419.
27. Larkin JE, Jones J, Zucker I (2002): Temperature
dependence of gonadal regression in Syrian hamsters exposed to short day lengths. Am J Physiol Regul Integr
Comp Physiol, 282, 744-752.
28. Matthews CD, Guerin MV, Deed JR (1993): Melatonin
and photoperiodic time measurement: Seasonal breeding in the sheep. J Pineal Res 14, 105–116.
29. McKinnon AO, Voss JL (1993): Equine Reproduction. Wiley-Blackwell, USA.
30. Nagy P, Guillaume D, Daels P (2000): Seasonality in
mares. Anim Reprod Sci, 60-61, 245-262.
31. Oxender WD, Noden PA, Hafs HD (1977): Estrus,
ovulation, and serum progesterone, estradiol, and LH concentrations in mares after an increased photoperiod during winter. Am J Vet Res, 38, 203-207.
32. Peltier MR, Robinson G, Sharp DC (1998): Effects of
melatonin implants in pony mares. 2. Long-term effects.
Theriogenology, 49, 1125-1142.
33. Sharp DC, Grubaugh WR, Zavy MT, Vernon MW (1980): Seasonal variation in melatonin secretory patterns
in mares. J Anim Sci, 51, 327.
34. Sharp DC, Grubaugh WR, Weithenauer J, Davis SD, Wilcox CJ (1991): Effects of steroid administration on
pituitary luteinizing hormone and follicle-stimulating hormone in ovariectomized pony mares in the early spring: pituitary responsiveness to gonadotropin-releasing hormone and pituitary gonadotropin content. Biol Reprod,
44, 983-90.
35. Stankov B, Cozzi B, Lucini V, Fumagalli P, Scaglione F, Fraschini F (1991): Characterization and mapping of
melatonin receptors in the brain of three mammalian species: rabbit, horse and sheep. A comparative in vitro binding study. Neuroendocrinology, 53, 214-221.
36. Storer WA (2002): The Effects of Growth Hormone or
Melatonin on the Reproductive Axis of Stallions. Master's
Thesis, Louisiana State University.
37. Tamarkin L, Westrom WK, Hamill AI, Goldman BD (1976): Effect of melatonin on the reproductive systems of
male and female Syrian hamsters: a diurnal rhythm in sensitivity to melatonin. Endocrinology, 99, 1534-1541.
38. Thompson DL Jr, Pickett BW, Berndtson WE, Voss JL, Mett TM (1977): Reproductive physiology of the
stallion. VIII. Artificial photoperiod, collection interval and seminal characteristics, sexual behavior and concentrations of LH and testosterone in serum. J Anim
Sci, 44, 656-664.
39. Thompson DL Jr, St George RL, Jones LS, Garza F Jr (1985): Patterns of secretion of luteinizing hormone,
follicle stimulating hormone and testosterone in stallions during the summer and winter. J Anim Sci, 60, 741-748. Geliş tarihi: 28.05.2008 / Kabul tarihi: 05.09.2008
Address for correspondence
Associate Professor Çiğdem Altınsaat, DVM, PhD, Ankara University, Faculty of Veterinary Medicine Department of Physiology
06110 Diskapi - Ankara, Turkey. e-mail:[email protected]